
Effectiveness of TaDreb-B1 and 1-FEH w3 KASP Markers in Spring and Winter Wheat Populations for Marker-Assisted Selection to Improve Drought Tolerance

 , and
, and
Abstract
:1. Introduction
2. Results
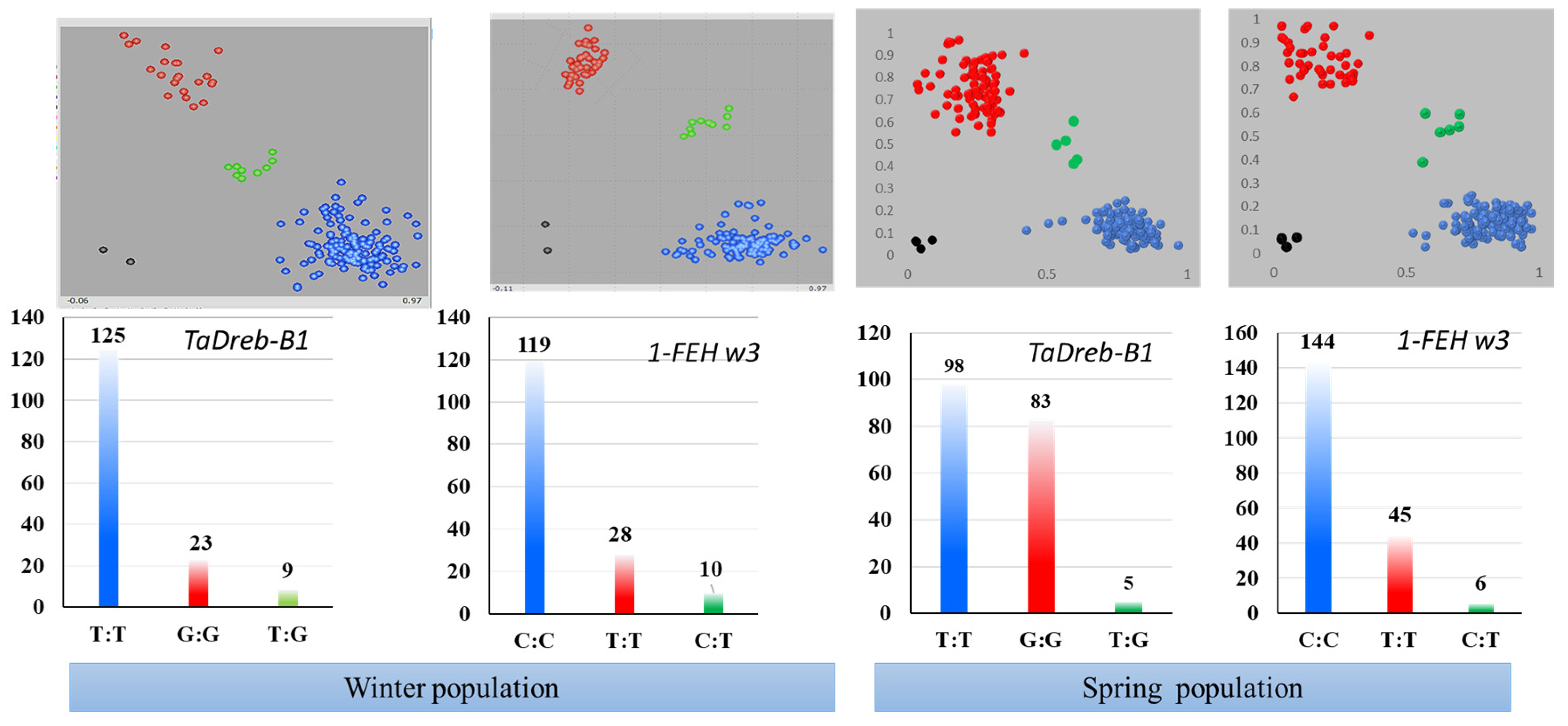
2.1. KASP Genotyping
2.2. The Diversity of TaDreb-B1 and 1-FEH w3 Markers
2.3. Single-Marker Analysis
2.3.1. Seedling Stage
2.3.2. Field Experiments
3. Discussion
3.1. Diversity of KASP Markers
3.2. Seedling Experiments
3.3. Field Experiments
4. Material and Methods
4.1. Plant Material
4.2. Seedlings Experiments
4.3. Field Experiments
4.4. DNA Extraction and Kompetitive Allele-Specific PCR (KASP) Genotyping
4.5. Diversity and Single-Marker Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, J.; Shi, W.; Zhang, Z.; Cheng, J.; Sun, D.; Yu, J.; Li, X.; Guo, P.; Hao, C. Association of Yield-Related Traits in Founder Genotypes and Derivatives of Common Wheat (Triticum aestivum L.). BMC Plant Biol. 2018, 18, 38. [Google Scholar] [CrossRef] [PubMed]
- Devate, N.B.; Krishna, H.; Parmeshwarappa, S.K.V.; Manjunath, K.K.; Chauhan, D.; Singh, S.; Singh, J.B.; Kumar, M.; Patil, R.; Khan, H.; et al. Genome-Wide Association Mapping for Component Traits of Drought and Heat Tolerance in Wheat. Front. Plant Sci. 2022, 13, 2886. [Google Scholar] [CrossRef] [PubMed]
- Pingali, P.L. Green Revolution: Impacts, Limits, and the Path Ahead. Proc. Natl. Acad. Sci. USA 2012, 109, 12302–12308. [Google Scholar] [CrossRef]
- Gajghate, R.; Chourasiya, D.; Sharma, R.K. Plant Morphological, Physiological Traits Associated with Adaptation against Heat Stress in Wheat and Maize. In Plant Stress Biology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 51–81. [Google Scholar]
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Lu, X.; Zhang, M.; Jin, J. Effect of Drought on Agronomic Traits of Rice and Wheat: A Meta-Analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Amin, A.E.E.A.Z.; Dawood, M.F.A. Genetic Variation in Kernel Traits under Lead and Tin Stresses in Spring Wheat Diverse Collection. Environ. Exp. Bot. 2021, 192, 104646. [Google Scholar] [CrossRef]
- Esmail, S.M.; Omar, G.E.; Mourad, A.M.I. In-Depth Understanding of the Genetic Control of Stripe Rust Resistance (Puccinia striiformis f. Sp. Tritici) Induced in Wheat (Triticum aestivum) by Trichoderma Asperellum T34. Plant Dis. 2023, 107, 457–472. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Abou-Zeid, M.A.; Eltaher, S.; Baenziger, P.S.; Börner, A. Identification of Candidate Genes and Genomic Regions Associated with Adult Plant Resistance to Stripe Rust in Spring Wheat. Agronomy 2021, 11, 2585. [Google Scholar] [CrossRef]
- Yang, H.; Jian, J.; Li, X.; Renshaw, D.; Clements, J.; Sweetingham, M.W.; Tan, C.; Li, C. Application of Whole Genome Re-Sequencing Data in the Development of Diagnostic DNA Markers Tightly Linked to a Disease-Resistance Locus for Marker-Assisted Selection in Lupin (Lupinus angustifolius). BMC Genom. 2015, 16, 660. [Google Scholar] [CrossRef]
- Liu, G.; Mullan, D.; Zhang, A.; Liu, H.; Liu, D.; Yan, G. Identification of KASP Markers and Putative Genes for Pre-Harvest Sprouting Resistance in Common Wheat (Triticum aestivum L.). Crop J. 2022, 11, 549–557. [Google Scholar] [CrossRef]
- Han, G.; Liu, S.; Jin, Y.; Jia, M.; Ma, P.; Liu, H.; Wang, J.; An, D. Scale Development and Utilization of Universal PCR-Based and High-Throughput KASP Markers Specific for Chromosome Arms of Rye (Secale cereale L.). BMC Genom. 2020, 21, 206. [Google Scholar] [CrossRef]
- Makhoul, M.; Rambla, C.; Voss-Fels, K.P.; Hickey, L.T.; Snowdon, R.J.; Obermeier, C. Overcoming Polyploidy Pitfalls: A User Guide for Effective SNP Conversion into KASP Markers in Wheat. Theor. Appl. Genet. 2020, 133, 2413–2430. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sehgal, S.K.; Lin, M.; Li, J.; Trick, H.N.; Gill, B.S.; Bai, G. Independent Mis-Splicing Mutations in Ta PHS 1 Causing Loss of Preharvest Sprouting (PHS) Resistance during Wheat Domestication. New Phytol. 2015, 208, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single Nucleotide Polymorphism Genotyping Using Kompetitive Allele Specific PCR (KASP): Overview of the Technology and Its Application in Crop Improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and Validation of KASP Assays for Genes Underpinning Key Economic Traits in Bread Wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef] [PubMed]
- Ur Rehman, S.; Ali Sher, M.; Saddique, M.A.B.; Ali, Z.; Khan, M.A.; Mao, X.; Irshad, A.; Sajjad, M.; Ikram, R.M.; Naeem, M.; et al. Development and Exploitation of KASP Assays for Genes Underpinning Drought Tolerance Among Wheat Cultivars From Pakistan. Front. Genet. 2021, 12, 789. [Google Scholar] [CrossRef]
- Mohamed, E.A.; Ahmed, A.A.M.; Schierenbeck, M.; Hussein, M.Y.; Baenziger, P.S.; Börner, A.; Sallam, A. Screening Spring Wheat Genotypes for TaDreb-B1 and Fehw3 Genes under Severe Drought Stress at the Germination Stage Using KASP Technology. Genes 2023, 14, 373. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Alqudah, A.M.; Baenziger, P.S.; Rasheed, A. Editorial: Genetic Validation and Its Role in Crop Improvement. Front. Genet. 2023, 13, 3705. [Google Scholar] [CrossRef]
- Ahmed, A.A.M.; Dawood, M.F.A.; Elfarash, A.; Mohamed, E.A.; Hussein, M.Y.; Börner, A.; Sallam, A. Genetic and Morpho-Physiological Analyses of the Tolerance and Recovery Mechanisms in Seedling Stage Spring Wheat under Drought Stress. Front. Genet. 2022, 13, 1010272. [Google Scholar] [CrossRef]
- Sallam, A.; Eltaher, S.; Alqudah, A.M.; Belamkar, V.; Baenziger, P.S. Combined GWAS and QTL Mapping Revealed Candidate Genes and SNP Network Controlling Recovery and Tolerance Traits Associated with Drought Tolerance in Seedling Winter Wheat. Genomics 2022, 114, 110358. [Google Scholar] [CrossRef]
- Eltaher, S.; Sallam, A.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Börner, A.; Stephen Baenziger, P.; Mourad, A.M.I. Genome-Wide Association Mapping Revealed SNP Alleles Associated with Spike Traits in Wheat. Agronomy 2022, 12, 1469. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Draz, I.S.; Omar, G.E.; Börner, A.; Esmail, S.M. Genome-Wide Screening of Broad-Spectrum Resistance to Leaf Rust (Puccinia Triticina Eriks) in Spring Wheat (Triticum aestivum L.). Front. Plant Sci. 2022, 13, 921230. [Google Scholar] [CrossRef] [PubMed]
- Alqudah, A.M.; Sallam, A.; Stephen Baenziger, P.; Börner, A. GWAS: Fast-Forwarding Gene Identification and Characterization in Temperate Cereals: Lessons from Barley—A Review. J. Adv. Res. 2020, 22, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a Genetic Linkage Map in Man Using Restriction Fragment Length Polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar] [PubMed]
- Mourad, A.M.I.; Belamkar, V.; Baenziger, P.S. Molecular Genetic Analysis of Spring Wheat Core Collection Using Genetic Diversity, Population Structure, and Linkage Disequilibrium. BMC Genom. 2020, 21, 434. [Google Scholar] [CrossRef]
- Salem, K.F.M.; Sallam, A. Analysis of Population Structure and Genetic Diversity of Egyptian and Exotic Rice (Oryza sativa L.) Genotypes. Comptes Rendus Biol. 2016, 339, 1–9. [Google Scholar] [CrossRef]
- Eltaher, S.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Poland, J.; Baenziger, P.S. Genetic Diversity and Population Structure of F3:6 Nebraska Winter Wheat Genotypes Using Genotyping-by-Sequencing. Front. Genet. 2018, 9, 76. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef]
- Sallam, A.; Mourad, A.M.I.; Hussain, W.; Stephen Baenziger, P. Genetic Variation in Drought Tolerance at Seedling Stage and Grain Yield in Low Rainfall Environments in Wheat (Triticum aestivum L.). Euphytica 2018, 214, 169. [Google Scholar] [CrossRef]
- Ahmed, A.A.M.; Mohamed, E.A.; Hussein, M.Y.; Sallam, A. Genomic Regions Associated with Leaf Wilting Traits under Drought Stress in Spring Wheat at the Seedling Stage Revealed by GWAS. Environ. Exp. Bot. 2021, 184, 104393. [Google Scholar] [CrossRef]
- Certified Crop Advisor Study Resources (Northeast Region). Available online: https://nrcca.cals.cornell.edu/soil/CA2/CA0212.1-3.php (accessed on 6 February 2023).
- Zhang, J.; Xu, Y.; Chen, W.; Dell, B.; Vergauwen, R.; Biddulph, B.; Khan, N.; Luo, H.; Appels, R.; van den Ende, W. A Wheat 1-FEH W3 Variant Underlies Enzyme Activity for Stem WSC Remobilization to Grain under Drought. New Phytol. 2015, 205, 293–305. [Google Scholar] [CrossRef]
- Mondal, S.; Sallam, A.; Sehgal, D.; Sukumaran, S.; Krishnan, J.N.; Kumar, U.; Biswal, A.; Mondal, S.; Sehgal, Á.D.; Sukumaran, Á.S.; et al. Advances in Breeding for Abiotic Stress Tolerance in Wheat. In Genomic Designing for Abiotic Stress Resistant Cereal Crops; Springer: Berlin/Heidelberg, Germany, 2021; pp. 71–103. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Alomari, D.Z.; Alqudah, A.M.; Sallam, A.; Salem, K.F.M. Recent Advances in Wheat (Triticum Spp.) Breeding. In Advances in Plant Breeding Strategies: Cereals; Springer: Berlin/Heidelberg, Germany, 2019; Volume 5. [Google Scholar]
- Yáñez, A.; Tapia, G.; Guerra, F.; del Pozo, A. Stem Carbohydrate Dynamics and Expression of Genes Involved in Fructan Accumulation and Remobilization during Grain Growth in Wheat (Triticum aestivum L.) Genotypes with Contrasting Tolerance to Water Stress. PLoS ONE 2017, 12, e0177667. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.; Coombes, B.; Joynson, R.; Hall, A.; Fellers, J.; Yang, C.Y.; Scholefield, D.; Ashling, S.; Isaac, P.; King, I.P.; et al. Chromosome-Specific KASP Markers for Detecting Amblyopyrum Muticum Segments in Wheat Introgression Lines. Plant Genome 2022, 15, e20193. [Google Scholar] [CrossRef] [PubMed]
- Patterson, G.T.; Carter, M.R. Soil Sampling and Handling, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press Taylor & Francis Group: Abingdon, UK, 2006; Volume 44, ISBN 9780849335860. [Google Scholar]
- Liu, K.; Muse, S.V. PowerMarker: An Integrated Analysis Environment for Genetic Marker Analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An Introduction to Markers, Quantitative Trait Loci (QTL) Mapping and Marker-Assisted Selection for Crop Improvement: The Basic Concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Francis, D.M.; Merk, H.L.; Namuth-Covert, D. Introduction to Single Marker Analysis (SMA). Available online: http://www.extension.org/pages/32552/introduction-to-single-marker-analysis-sma (accessed on 25 March 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://cran.r-project.org/web/packages/ggplot2/citation.html (accessed on 1 March 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Trait | Gene/Target Allele | p-Value | Effect of Target Allele |
|---|---|---|---|---|
| Lincoln | SL | 1-FEH w3/C | 0.01727 ** | −0.38931 |
| TaDreb-B1/T | 0.30918 | 0.17946 | ||
| GNPS | 1-FEH w3/C | 0.00517 ** | −3.2963 | |
| TaDreb-B1/T | 0.66216 | 0.56407 | ||
| SPS | 1-FEH w3/C | 0.03281 ** | −0.53918 | |
| TaDreb-B1/T | 0.95199 | 0.01643 | ||
| TKW | 1-FEH w3/C | 0.78477 | 0.44653 | |
| TaDreb-B1/T | 0.44112 | −0.17361 | ||
| SL | 1-FEH w3/C | 0.50759 | 0.08685 | |
| TaDreb-B1/T | 0.53501 | 0.08919 | ||
| GNPS | 1-FEH w3/C | 0.60044 | 0.34391 | |
| North Platte | TaDreb-B1/T | 0.63706 | 0.33435 | |
| SPS | 1-FEH w3/C | 0.79417 | −0.0495 | |
| TaDreb-B1/T | 0.89495 | 0.0268 | ||
| TKW | 1-FEH w3/C | 0.48396 | −0.3992 | |
| TaDreb-B1/T | 0.16237 | −0.8338 |
| Environment | Trait | Gene/Target Allele | p-Value | Effect of Target Allele |
|---|---|---|---|---|
| Control 2018 | ||||
| SL | 1-FEH w3/C | 0.647 | −0.307 | |
| TaDreb-B1/T | 0.105 | −0.882 | ||
| GNPS | 1-FEH w3/C | 0.143 | −5.190 | |
| TaDreb-B1/T | 0.169 | 4.115 | ||
| SPS | 1-FEH w3/C | 0.941 | −0.033 | |
| TaDreb-B1/T | 0.788 | 0.103 | ||
| TKW | 1-FEH w3/C | 0.852 | −0.297 | |
| TaDreb-B1/T | 0.461 | −1.002 | ||
| Drought 2018 | ||||
| SL | 1-FEH w3/C | 0.371 | −0.381 | |
| TaDreb-B1/T | 0.000096 ** | −1.393 | ||
| GNPS | 1-FEH w3/C | 0.021180 * | 3.464 | |
| TaDreb-B1/T | 0.191 | 5.059 | ||
| SPS | 1-FEH w3/C | 0.831 | −0.078 | |
| TaDreb-B1/T | 0.613 | 0.152 | ||
| TKW | 1-FEH w3/C | 0.966 | −0.073 | |
| TaDreb-B1/T | 0.330 | −1.382 | ||
| Control 2019 | ||||
| SL | 1-FEH w3/C | 0.500 | −0.358 | |
| TaDreb-B1/T | 0.00037 ** | −1.540 | ||
| GNPS | 1-FEH w3/C | 0.02732 * | −6.738 | |
| TaDreb-B1/T | 0.836 | 0.519 | ||
| SPS | 1-FEH w3/C | 0.420 | −0.374 | |
| TaDreb-B1/T | 0.523 | 0.251 | ||
| TKW | 1-FEH w3/C | 0.503 | −0.455 | |
| TaDreb-B1/ T | 0.082 | −0.942 | ||
| Drought 2019 | ||||
| SL | 1-FEH w3/C | 0.651 | −0.198 | |
| TaDreb-B1/T | 0.00051 ** | −1.246 | ||
| GNPS | 1-FEH w3/C | 0.00811 ** | −8.161 | |
| TaDreb-B1/T | 0.524 | -d1.588 | ||
| SPS | 1-FEH w3/C | 0.356 | −0.377 | |
| TaDreb-B1/T | 0.201 | 0.444 | ||
| TKW | 1-FEH w3/C | 0.802 | −0.358 | |
| TaDreb-B1/T | 0.546 | −0.718 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltaher, S.; Hashem, M.; Ahmed, A.A.M.; Baenziger, P.S.; Börner, A.; Sallam, A. Effectiveness of TaDreb-B1 and 1-FEH w3 KASP Markers in Spring and Winter Wheat Populations for Marker-Assisted Selection to Improve Drought Tolerance. Int. J. Mol. Sci. 2023, 24, 8986. https://doi.org/10.3390/ijms24108986
Eltaher S, Hashem M, Ahmed AAM, Baenziger PS, Börner A, Sallam A. Effectiveness of TaDreb-B1 and 1-FEH w3 KASP Markers in Spring and Winter Wheat Populations for Marker-Assisted Selection to Improve Drought Tolerance. International Journal of Molecular Sciences. 2023; 24(10):8986. https://doi.org/10.3390/ijms24108986
Chicago/Turabian StyleEltaher, Shamseldeen, Mostafa Hashem, Asmaa A. M. Ahmed, P. Stephen Baenziger, Andreas Börner, and Ahmed Sallam. 2023. "Effectiveness of TaDreb-B1 and 1-FEH w3 KASP Markers in Spring and Winter Wheat Populations for Marker-Assisted Selection to Improve Drought Tolerance" International Journal of Molecular Sciences 24, no. 10: 8986. https://doi.org/10.3390/ijms24108986