
The Sts Proteins: Modulators of Host Immunity

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Sts Catalytic Domains
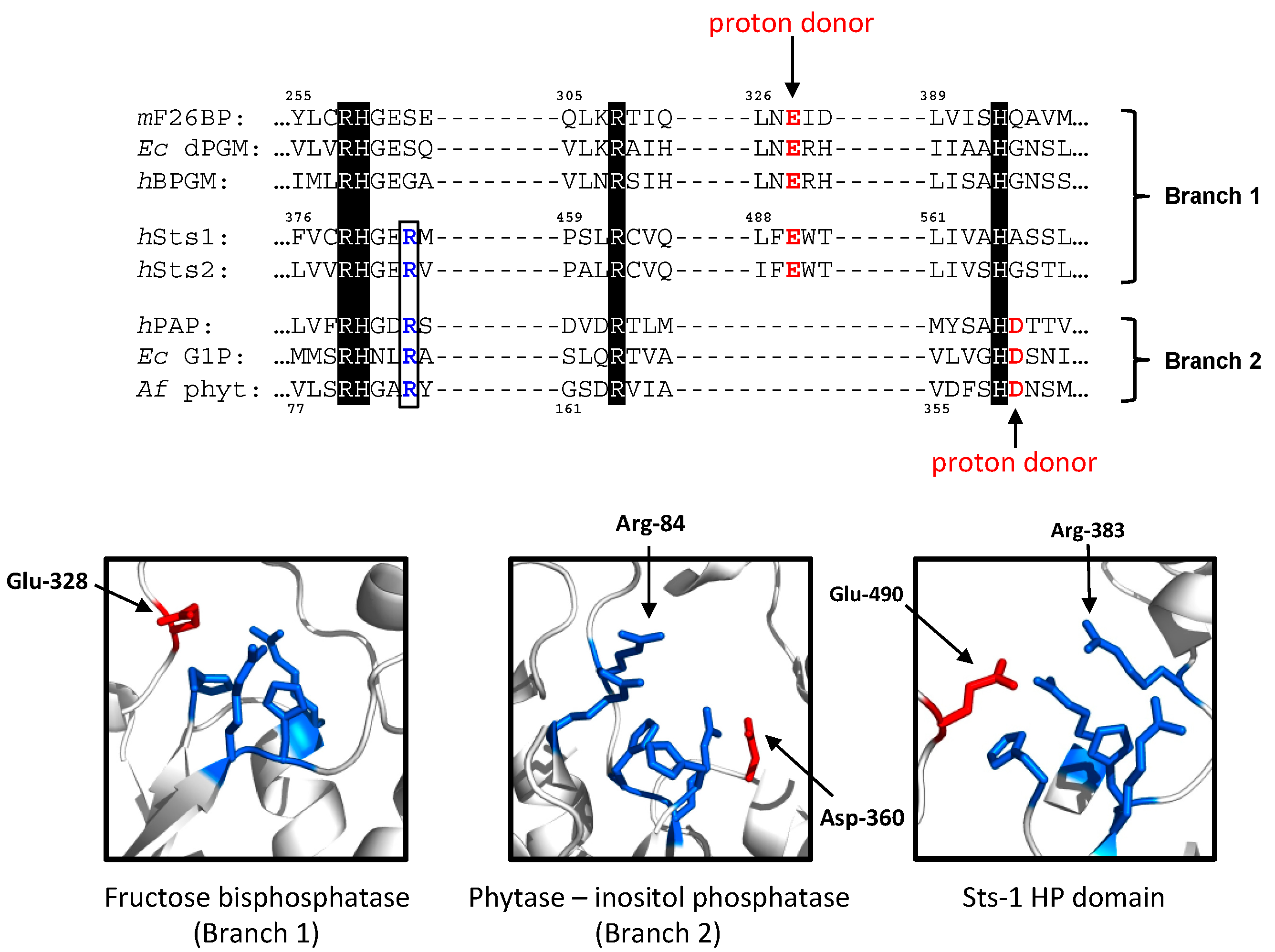
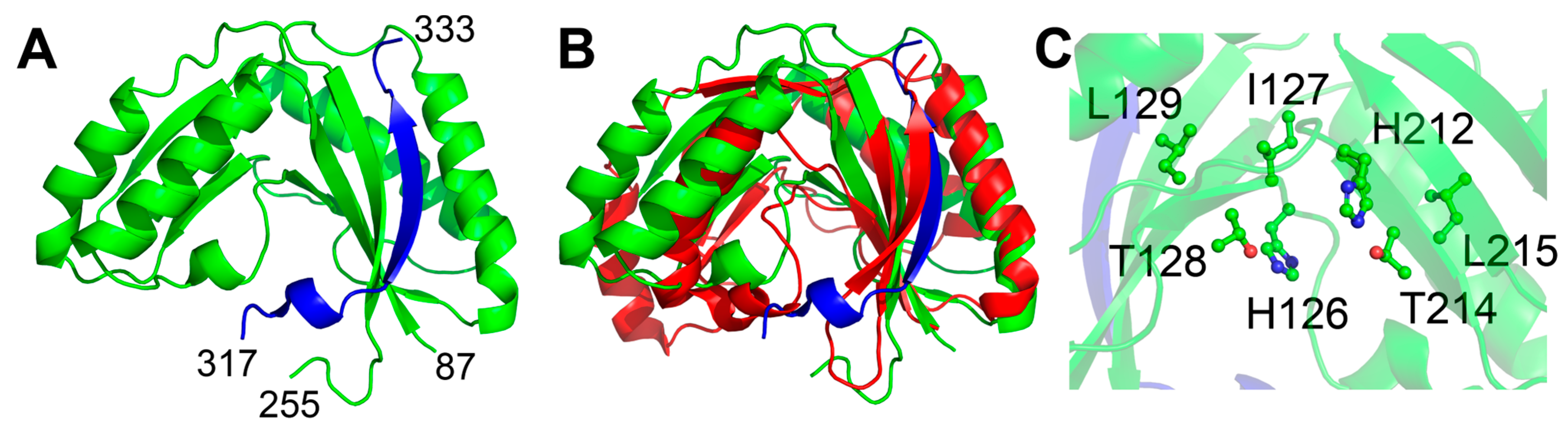
2.1.1. The Sts Histidine Phosphatase (HP) Domain
2.1.2. The Sts Phosphodiesterase (PDE) Domain
2.2. Functional Studies
2.2.1. The Role of Sts in T Cells
2.2.2. The Role of Sts in Additional Cell Types
2.2.3. The Role of Sts in Regulating Anti-Microbial Immunity
- Candida albicans
- 2.
- Francisella tularensis
3. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsygankov, A.Y. TULA proteins as signaling regulators. Cell Signal. 2020, 65, 109424. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Dixon, J.E.; Manning, G. Genomics and evolution of protein phosphatases. Sci. Signal. 2017, 10, eaag1796. [Google Scholar] [CrossRef] [PubMed]
- Carpino, N.; Turner, S.; Mekala, D.; Takahashi, Y.; Zang, H.; Geiger, T.L.; Doherty, P.; Ihle, J.N. Regulation of ZAP-70 activation and TCR signaling by two related proteins, Sts-1 and Sts-2. Immunity 2004, 20, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Rigden, D.J. The histidine phosphatase superfamily: Structure and function. Biochem. J. 2008, 409, 333–348. [Google Scholar] [CrossRef]
- Jedrzejas, M.J. Structure, function, and evolution of phosphoglycerate mutases: Comparison with fructose-2,6-bisphosphatase, acid phosphatase, and alkaline phosphatase. Prog. Biophys. Mol. Biol. 2000, 73, 263–287. [Google Scholar] [CrossRef]
- Winn, S.I.; Watson, H.C.; Harkins, R.N.; Fothergill, L.A. Structure and activity of phosphoglycerate mutase. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1981, 293, 121–130. [Google Scholar]
- Mikhailik, A.; Ford, B.; Keller, J.; Chen, Y.; Nassar, N.; Carpino, N. A phosphatase activity of Sts-1 contributes to the suppression of TCR signaling. Mol. Cell 2007, 27, 486–497. [Google Scholar] [CrossRef]
- Chen, Y.; Jakoncic, J.; Carpino, N.; Nassar, N. Structural and functional characterization of the 2H-phosphatase domain of Sts-2 reveals an acid-dependent phosphatase activity. Biochemistry 2009, 48, 1681–1690. [Google Scholar] [CrossRef]
- San Luis, B.; Nassar, N.; Carpino, N. New insights into the catalytic mechanism of histidine phosphatases revealed by a functionally essential arginine residue within the active site of the Sts phosphatases. Biochem. J. 2013, 453, 27–35. [Google Scholar] [CrossRef]
- Yamada, R.; Sonobe, H. Purification, kinetic characterization, and molecular cloning of a novel enzyme ecdysteroid phosphate phosphatase. J. Biol. Chem. 2003, 278, 26365–26373. [Google Scholar] [CrossRef]
- Davies, L.; Anderson, I.P.; Turner, P.C.; Shirras, A.D.; Rees, H.H.; Rigden, D.J. An unsuspected ecdysteroid/steroid phosphatase activity in the key T-cell regulator, Sts-1, Surprising relationship to insect ecdysteroid phosphate phosphatase. Proteins 2007, 67, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Raguz, J.; Wagner, S.; Dikic, I.; Hoeller, D. Suppressor of T-cell receptor signalling 1 and 2 differentially regulate endocytosis and signalling of receptor tyrosine kinases. FEBS Lett. 2007, 581, 4767–4772. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Carpino, N.; Tsygankov, A. TULA proteins regulate activity of the protein tyrosine kinase Syk. J. Cell. Biochem. 2008, 104, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ren, L.; Kim, S.; Carpino, N.; Daniel, J.L.; Kunapuli, S.P.; Tsygankov, A.Y.; Pei, D. Determination of the substrate specificity of protein-tyrosine phosphatase TULA-2 and identification of Syk as a TULA-2 substrate. J. Biol. Chem. 2010, 285, 31268–31276. [Google Scholar] [CrossRef]
- San Luis, B.; Sondgeroth, B.; Nassar, N.; Carpino, N. Sts-2 is a phosphatase that negatively regulates zeta-associated protein (ZAP)-70 and T cell receptor signaling pathways. J. Biol. Chem. 2011, 286, 15943–15954. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, R.; Iyer, L.M.; Vasudevan, S.; Aravind, L. Detection of novel members, structure-function analysis and evolutionary classification of the 2H phosphoesterase superfamily. Nucleic Acids Res. 2002, 30, 5229–5243. [Google Scholar] [CrossRef] [PubMed]
- Raasakka, A.; Myllykoski, M.; Laulumaa, S.; Lehtimaki, M.; Harlein, M.; Moulin, M.; Kursula, I.; Kursula, P. Determinants of ligand binding and catalytic activity in the myelin enzyme 2′3′-cyclic nucelotide 3′-phosphodiesterase. Sci. Rep. 2015, 5, 16520. [Google Scholar] [CrossRef]
- Raasakka, A.; Kursula, P. The myelin membrane-associated enzyme 2′,3′-cyclic nucleotide 3′-phosphodiesterase: On a highway to structure and function. Neurosci. Bull. 2014, 30, 956–966. [Google Scholar] [CrossRef]
- Yin, Y.; Frank, D.; Zhou, W.; Kaur, N.; French, J.B.; Carpino, N. An unexpected 2-histidine phosphoesterase activity of suppressor of T-cell receptor signaling protein 1 contributes to the suppression of cell signaling. J. Biol. Chem. 2020, 296, 8514–8523. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.; Liang, Y.-H.; Li, L.-F.; Su, X.-D. Crystal structure of B. subtilis YjcG characterizing the YjcV-like group of 2H phosphoesterase superfamily. Proteins 2008, 72, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Kowanetz, K.; Crosetto, N.; Haglund, K.; Schmidt, M.H.; Heldin, C.H.; Dikic, I. Suppressors of T-cell receptor signaling Sts-1 and Sts-2 bind to Cbl and inhibit endocytosis of receptor tyrosine kinases. J. Biol. Chem. 2004, 279, 32786–32795. [Google Scholar] [CrossRef] [PubMed]
- Feshchenko, E.A.; Smirnova, E.V.; Swaminathan, G.; Teckchandani, A.M.; Agrawal, R.; Band, H.; Zhang, X.; Annan, R.S.; Carr, S.A.; Tsygankov, A.Y. TULA: An SH3- and UBA-containing protein that binds to c-Cbl and ubiquitin. Oncogene 2004, 23, 4690–4706. [Google Scholar] [CrossRef] [PubMed]
- Carpino, N.; Kobayashi, R.; Zang, H.; Takahashi, Y.; Jou, S.T.; Feng, J.; Nakajima, H.; Ihle, J.N. Identification, cDNA cloning and targeted deletion of p70, a novel, ubiquitiously expressed SH3 domain-containing protein. Mol. Cell. Biol. 2002, 22, 7491–7500. [Google Scholar] [CrossRef] [PubMed]
- Newman, T.N.; Liverani, E.; Ivanova, E.; Russo, G.L.; Carpino, N.; Ganea, D.; Safadi, F.; Kunapuli, S.P.; Tsygankov, A.Y. Members of the novel UBASH3/STS/TULA family of cellular regulators suppress T-cell-driven inflammatory responses in vivo. Immunol. Cell Biol 2014, 92, 837–850. [Google Scholar] [CrossRef]
- Okabe, N.; Ohmura, K.; Katayama, M.; Akizuki, S.; Carpino, N.; Murakami, K.; Nakashima, R.; Hashimoto, M.; Imura, Y.; Yoshifuji, H.; et al. Suppressor of TCR signaling-2 (Sts-2) suppresses arthritis development in mice. Mod. Rheumatol. 2017, 28, 626–636. [Google Scholar] [CrossRef]
- Wang, H.; Kadlecek, T.A.; Au-Yeung, B.B.; Sjolin Goodfellow, H.E.; Hsu, L.-Y.; Freedman, T.S.; Weiss, A. Zap-70, an essential kinase in T-cell signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a002279. [Google Scholar] [CrossRef] [PubMed]
- Carpino, N.; Chen, Y.; Nassar, N.; Oh, H.-W. The Sts proteins target tyrosine phosphorylated, ubiquitinated proteins within TCR signaling pathways. Mol. Immunol. 2009, 46, 3224–3231. [Google Scholar] [CrossRef]
- Hope, J.L.; Otero, D.C.; Bae, E.-A.; Stairiker, C.J.; Palete, A.B.; Faso, H.A.; Lin, M.; Henriquez, M.L.; Roy, S.; Seo, H.; et al. PSGL-1 attenuates early TCR signaling to suppress CD8+ T cell progenitor differentiation and elicit terminal CD8+ T cell exhaustion. Cell Rep. 2023, 42, 112436. [Google Scholar] [CrossRef]
- Yang, M.; Chen, T.; Li, X.; Yu, Z.; Tang, S.; Wang, C.; Gu, Y.; Liu, Y.; Xu, S.; Li, W.; et al. K33-linked polyubiquitination of Zap70 by Nrdp1 controls CD8(+) T cell activation. Nat. Immunol. 2015, 16, 1253–1262. [Google Scholar] [CrossRef]
- Hu, H.; Wang, H.; Xiao, Y.; Jin, J.; Chang, J.H.; Zou, Q.; Xie, X.; Cheng, X.; Sun, S.C. Otud7b facilitates T cell activation and inflammatory responses by regulating Zap70 ubiquitination. J. Exp. Med. 2016, 213, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Paisie, T.K.; Newman, J.R.B.; McIntyre, L.M.; Concannon, P. UBASH3 mediates risk for Type 1 diabetes through inhibition of T-cell receptor-induced NF-kB signaling. Diabetes 2017, 66, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Paisie, T.K.; Chen, S.; Concannon, P. UBASH3 regulates the synthesis and dynamics of the TCR-CD3 complexes. J. Immunol. 2019, 203, 2827–2836. [Google Scholar] [CrossRef] [PubMed]
- Monticone, G.; Huang, Z.; Csibi, F.; Leit, S.; Ciccone, D.; Champhekar, A.S.; Austin, J.E.; Ucar, D.A.; Hossain, F.; Ibba, S.V.; et al. Targeting the Cbl-b-Notch1 axis as a novel immuno-therapeutic strategy to boost CD8+ T-cell responses. Front. Immunol. 2022, 13, 987298. [Google Scholar] [CrossRef] [PubMed]
- Reppschlager, K.; Gosselin, J.; Dangelmaier, C.A.; Thomas, D.H.; Carpino, N.; McKenzie, S.E.; Kunapuli, S.P.; Tsygankov, A.Y. TULA-2 protein phosphatase suppresses activation of Syk through the GPVI platelet receptor for collagen by dephosphorylating Tyr(P)346, a regulatory site of Syk. J. Biol. Chem. 2016, 291, 22427–22441. [Google Scholar] [CrossRef]
- Thomas, D.H.; Getz, T.M.; Newman, T.N.; Dangelmaier, C.A.; Carpino, N.; Kunapuli, S.P.; Tsygankov, A.Y.; Daniel, J.L. A novel histidine tyrosine phosphatase, TULA-2, associates with Syk and negatively regulates GPVI signaling in platelets. Blood 2016, 116, 2570–2578. [Google Scholar] [CrossRef]
- de Castro, R.O.; Zhang, J.; Groves, J.R.; Barbu, E.A.; Siraganian, R.P. Once phosphorylated, tyrosines in carboxyl terminus of protein-tyrosine kinase Syk interact with signaling proteins, including TULA-2, a negative regulator of mast cell degranulation. J. Biol. Chem. 2012, 287, 8194–8204. [Google Scholar] [CrossRef]
- Back, S.H.; Adapala, N.S.; Barbe, M.F.; Carpino, N.; Tsygankov, A.Y.; Sanjay, A. TULA-2, a novel histidine phosphatase, regulates bone remodeling by modulating osteoclast function. Cell. Mol. Life Sci. 2013, 70, 1269–1284. [Google Scholar] [CrossRef]
- Zhang, J.; Vakhursheva, O.; Bandi, S.R.; Demirel, O.; Kazi, J.U.; Fernandes, R.G.; Jakobi, K.; Eichler, A.; Ronnstrand, L.; Rieger, M.A.; et al. The phosphatases Sts-1 and Sts-2 regulate hematopoietic stem and progenitor cell fitness. Stem Cell Rep. 2015, 5, 633–646. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef]
- Lionakis, M.S. New insights into innate immune control of systemic candidiasis. Med. Mycol. 2014, 52, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Mice with disseminated candidiasis die of progressive sepsis. J. Infect. Dis. 2005, 192, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Naseem, S.; Frank, D.; Konopka, J.B.; Carpino, N. Protection from systemic Candida albicans infection by inactivation of the Sts phosphatases. Infect. Immun. 2015, 83, 637–645. [Google Scholar] [CrossRef]
- Frank, D.; Naseem, S.; Russo, G.L.; Li, C.; Parashar, K.; Konopka, J.B.; Carpino, N. Phagocytes from mice lacking the Sts phosphatases have an enhanced antifungal response to Candida albicans. mBio 2018, 9, e00782-18. [Google Scholar] [CrossRef] [PubMed]
- McLendon, M.K.; Apicella, M.A.; Allen, L.-H.H. Francisella tularensis: Taxonomy, genetics, and immunopathogenesis of a potential agent of biowarfare. Annu. Rev. Micobiol. 2006, 60, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Parashar, K.; Kopping, E.; Frank, D.; Sampath, V.; Thanassi, D.G.; Carpino, N. Increased resistance to intradermal Francisella tularensis LVS infection by inactivation of the Sts phosphatases. Infect. Immun. 2017, 85, e00406-17. [Google Scholar] [CrossRef]
- Parashar, K.; Carpino, N. A role for the Sts phosphatases in negatively regulating IFNγ-mediated production of nitric oxide in monocytes. Immun. Inflamm. Dis. 2020, 8, 523–533. [Google Scholar] [CrossRef]
- Shapiro, M.R.; Thirawatananond, P.; Peters, L.; Sharp, R.C.; Ogundare, S.; Posgai, A.L.; Perry, D.J.; Brusko, T.M. De-coding genetic risk variants in type 1 diabetes. Immunol. Cell Biol. 2021, 99, 496–508. [Google Scholar] [CrossRef]
- Tsygankov, A.Y. TULA-family proteins: Jacks of many trades and then some. J. Cell. Physiol. 2019, 234, 274–288. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaman, A.; French, J.B.; Carpino, N. The Sts Proteins: Modulators of Host Immunity. Int. J. Mol. Sci. 2023, 24, 8834. https://doi.org/10.3390/ijms24108834
Zaman A, French JB, Carpino N. The Sts Proteins: Modulators of Host Immunity. International Journal of Molecular Sciences. 2023; 24(10):8834. https://doi.org/10.3390/ijms24108834
Chicago/Turabian StyleZaman, Anika, Jarrod B. French, and Nick Carpino. 2023. "The Sts Proteins: Modulators of Host Immunity" International Journal of Molecular Sciences 24, no. 10: 8834. https://doi.org/10.3390/ijms24108834