
Modulation of Anthocyanin Biosynthesis-Related Genes during the Ripening of Olea europaea L. cvs Carolea and Tondina Drupes in Relation to Environmental Factors
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Temperature Detection
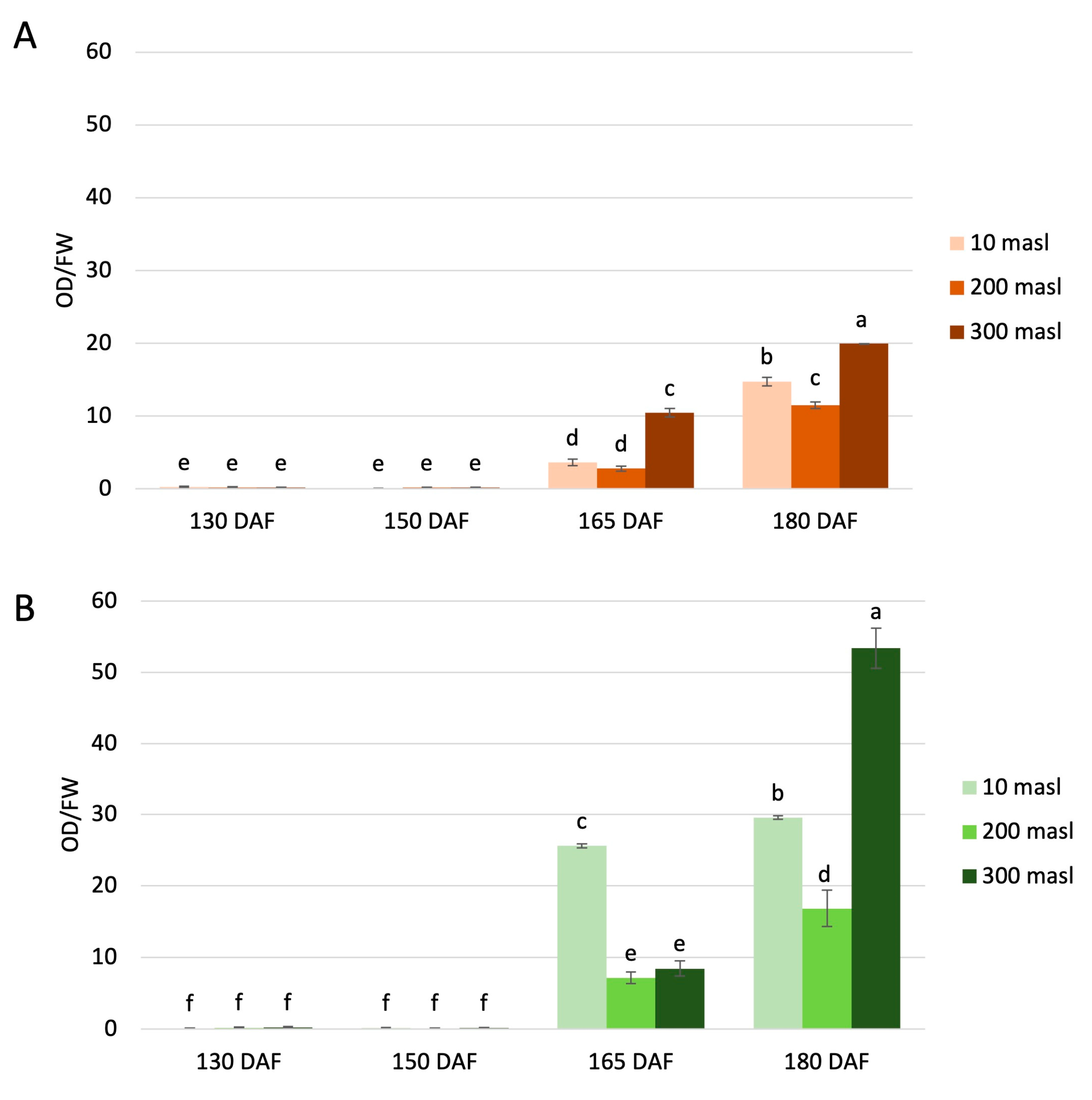
2.2. Total Anthocyanins
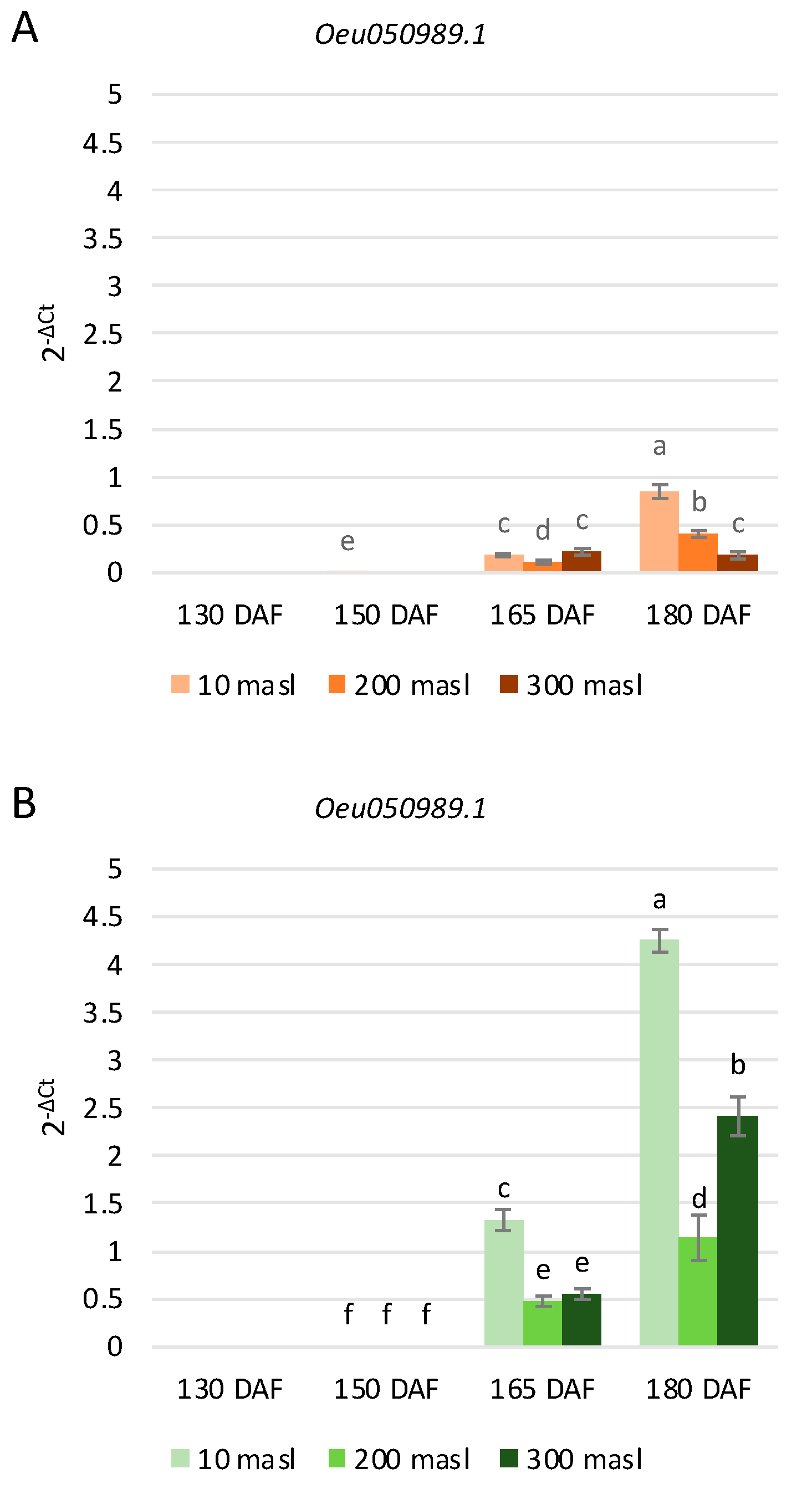
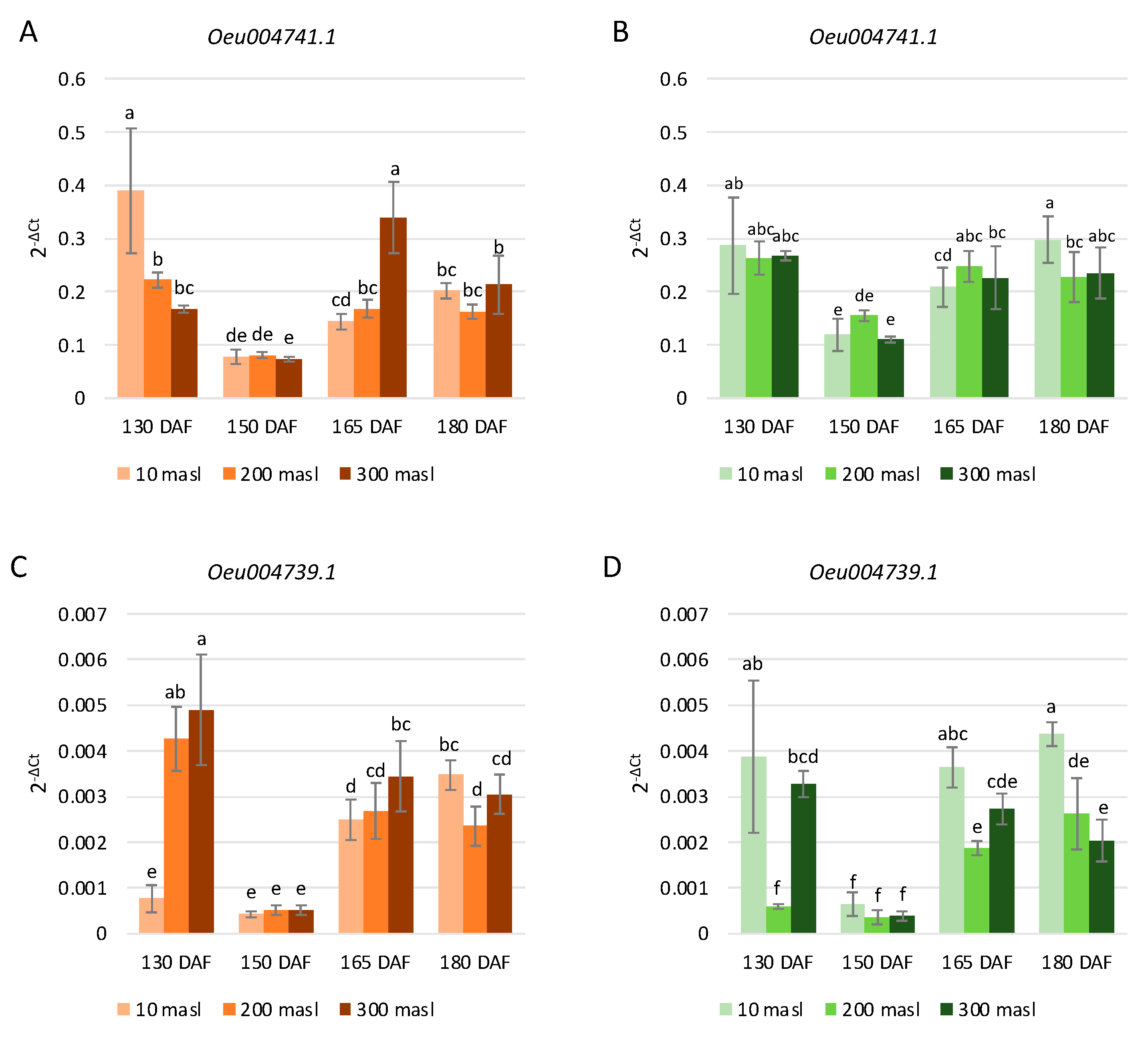
2.3. Expression Profiles of Anthocyanin Biosynthesis-Related Genes
2.4. Selection, Classification, and Phylogenetic Analysis of the R2R3-MYB Gene Family in Olea europaea var sylvestris
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Total Anthocyanin Quantification
4.3. RNA Isolation and Real-Time Quantitative PCR Analysis
4.4. Identification and Phylogenetic Analysis of the O. europaea var sylvestris OeR2R3-MYB Proteins
4.5. Pearson Correlation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carrión, Y.; Ntinou, M.; Badal, E. Olea Europaea L. in the North Mediterranean Basin during the Pleniglacial and the Early–Middle Holocene. Quat. Sci. Rev. 2010, 29, 952–968. [Google Scholar] [CrossRef]
- Besnard, G.; Rubio de Casas, R.; Christin, P.-A.; Vargas, P. Phylogenetics of Olea (Oleaceae) Based on Plastid and Nuclear Ribosomal DNA Sequences: Tertiary Climatic Shifts and Lineage Differentiation Times. Ann. Bot. 2009, 104, 143–160. [Google Scholar] [CrossRef]
- Alagna, F.; D’Agostino, N.; Torchia, L.; Servili, M.; Rao, R.; Pietrella, M.; Giuliano, G.; Chiusano, M.L.; Baldoni, L.; Perrotta, G. Comparative 454 Pyrosequencing of Transcripts from Two Olive Genotypes during Fruit Development. BMC Genom. 2009, 10, 399. [Google Scholar] [CrossRef]
- Bianco, L.; Alagna, F.; Baldoni, L.; Finnie, C.; Svensson, B.; Perrotta, G. Proteome Regulation during Olea Europaea Fruit Development. PLoS ONE 2013, 8, e53563. [Google Scholar] [CrossRef]
- Skodra, C.; Titeli, V.S.; Michailidis, M.; Bazakos, C.; Ganopoulos, I.; Molassiotis, A.; Tanou, G. Olive Fruit Development and Ripening: Break on through to the “-Omics” Side. Int. J. Mol. Sci. 2021, 22, 5806. [Google Scholar] [CrossRef]
- Giovannoni, J.J. Genetic Regulation of Fruit Development and Ripening. Plant Cell 2004, 16, S170–S180. [Google Scholar] [CrossRef]
- Connor, A.M.; Luby, J.J.; Tong, C.B.S.; Finn, C.E.; Hancock, J.F. Genotypic and Environmental Variation in Antioxidant Activity, Total Phenolic Content, and Anthocyanin Content among Blueberry Cultivars. J. Am. Soc. Hortic. Sci. 2002, 127, 89–97. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Di Gaspero, G. Transcriptional Control of Anthocyanin Biosynthetic Genes in Extreme Phenotypes for Berry Pigmentation of Naturally Occurring Grapevines. BMC Plant Biol. 2007, 7, 46. [Google Scholar] [CrossRef]
- Stevenson, D.; Scalzo, J. Anthocyanin Composition and Content of Blueberries from around the World. J. Berry Res. 2012, 2, 179–189. [Google Scholar] [CrossRef]
- Davies, K.M.; Albert, N.W.; Schwinn, K.E. From Landing Lights to Mimicry: The Molecular Regulation of Flower Colouration and Mechanisms for Pigmentation Patterning. Functional Plant Biol. 2012, 39, 619–638. [Google Scholar] [CrossRef]
- Ahmed, N.U.; Park, J.-I.; Jung, H.-J.; Hur, Y.; Nou, I.-S. Anthocyanin Biosynthesis for Cold and Freezing Stress Tolerance and Desirable Color in Brassica Rapa. Funct. Integr. Genom. 2015, 15, 383–394. [Google Scholar] [CrossRef]
- Han, F.; Yang, P.; Wang, H.; Fernandes, I.; Mateus, N.; Liu, Y. Digestion and Absorption of Red Grape and Wine Anthocyanins through the Gastrointestinal Tract. Trends Food Sci. Technol. 2019, 83, 211–224. [Google Scholar] [CrossRef]
- Li, X.; Yao, Z.; Yang, D.; Jiang, X.; Sun, J.; Tian, L.; Hu, J.; Wu, B.; Bai, W. Cyanidin-3-O-Glucoside Restores Spermatogenic Dysfunction in Cadmium-Exposed Pubertal Mice via Histone Ubiquitination and Mitigating Oxidative Damage. J. Hazard. Mater. 2020, 387, 121706. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Verweij, W.; Kroon, A.; Spelt, C.; Mol, J.; Koes, R. PH4 of Petunia Is an R2R3 MYB Protein That Activates Vacuolar Acidification through Interactions with Basic-Helix-Loop-Helix Transcription Factors of the Anthocyanin Pathway. Plant Cell 2006, 18, 1274–1291. [Google Scholar] [CrossRef]
- Li, S. Transcriptional Control of Flavonoid Biosynthesis. Plant Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef]
- Conde, C.; Delrot, S.; Gerós, H. Physiological, Biochemical and Molecular Changes Occurring during Olive Development and Ripening. J. Plant Physiol. 2008, 165, 1545–1562. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB Transcription Factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Reif, H.J.; Niesbach, U.; Deumling, B.; Saedler, H. Cloning and Analysis of Two Genes for Chalcone Synthase from Petunia Hybrida. Mol. Genet. Genom. 1985, 199, 208–215. [Google Scholar] [CrossRef]
- Beld, M.; Martin, C.; Huits, H.; Stuitje, A.R.; Gerats, A.G.M. Flavonoid Synthesis in Petunia Hybrida: Partial Characterization of Dihydroflavonol-4-Reductase Genes. Plant Mol. Biol. 1989, 13, 491–502. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of Plant Pigments: Anthocyanins, Betalains and Carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Martinelli, F.; Tonutti, P. Flavonoid Metabolism and Gene Expression in Developing Olive (Olea Europaea L.) Fruit. Plant Biosyst. 2012, 146, 164–170. [Google Scholar] [CrossRef]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef]
- Iaria, D.L.; Chiappetta, A.; Muzzalupo, I. A De Novo Transcriptomic Approach to Identify Flavonoids and Anthocyanins “Switch-Off” in Olive (Olea Europaea L.) Drupes at Different Stages of Maturation. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef]
- Guodong, R.; Jianguo, Z.; Xiaoxia, L.; Ying, L. Identification of Putative Genes for Polyphenol Biosynthesis in Olive Fruits and Leaves Using Full-Length Transcriptome Sequencing. Food Chem. 2019, 300, 125246. [Google Scholar] [CrossRef]
- Durbin, M.L.; Learn, G.H.; Huttley, G.A.; Clegg, M.T. Evolution of the Chalcone Synthase Gene Family in the Genus Ipomoea. Proc. Natl. Acad. Sci. USA 1995, 92, 3338–3342. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Han, Y.; Wei, G.; Korban, S.S. An Apple MYB Transcription Factor, MdMYB3, Is Involved in Regulation of Anthocyanin Biosynthesis and Flower Development. BMC Plant Biol. 2013, 13, 176. [Google Scholar] [CrossRef]
- Chen, L.; Hu, B.; Qin, Y.; Hu, G.; Zhao, J. Advance of the Negative Regulation of Anthocyanin Biosynthesis by MYB Transcription Factors. Plant Physiol. Biochem. 2019, 136, 178–187. [Google Scholar] [CrossRef]
- Bruno, L.; Picardi, E.; Pacenza, M.; Chiappetta, A.; Muto, A.; Gagliardi, O.; Muzzalupo, I.; Pesole, G.; Bitonti, M.B. Changes in Gene Expression and Metabolic Profile of Drupes of Olea Europaea L. Cv Carolea in Relation to Maturation Stage and Cultivation Area. BMC Plant Biol. 2019, 19, 428. [Google Scholar] [CrossRef]
- Mondal, S.K.; Roy, S. Genome-Wide Sequential, Evolutionary, Organizational and Expression Analyses of Phenylpropanoid Biosynthesis Associated MYB Domain Transcription Factors in Arabidopsis. J. Biomol. Struct. 2018, 36, 1577–1601. [Google Scholar] [CrossRef]
- Li, T.; Jia, K.-P.; Lian, H.-L.; Yang, X.; Li, L.; Yang, H.-Q. Jasmonic Acid Enhancement of Anthocyanin Accumulation Is Dependent on Phytochrome A Signaling Pathway under Far-Red Light in Arabidopsis. Biochem. Biophys. Res. Commun. 2014, 454, 78–83. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional Repression by AtMYB4 Controls Production of UV-Protecting Sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef]
- Amiot, M.J.; Fleuriet, A.; Macheix, J.J. Importance and Evolution of Phenolic Compounds in Olive during Growth and Maturation. J. Agric. Food Chem. 1986, 34, 823–826. [Google Scholar] [CrossRef]
- Karppinen, K.; Zoratti, L.; Nguyenquynh, N.; Häggman, H.; Jaakola, L. On the Developmental and Environmental Regulation of Secondary Metabolism in Vaccinium Spp. Berries. Front. Plant Sci. 2016, 7, 655. [Google Scholar] [CrossRef]
- Zoratti, L.; Jaakola, L.; Häggman, H.; Giongo, L. Modification of Sunlight Radiation through Colored Photo-Selective Nets Affects Anthocyanin Profile in Vaccinium Spp. Berries. PLoS ONE 2015, 10, e0135935. [Google Scholar] [CrossRef]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid Biosynthesis-Related Genes in Grape Skin Are Differentially Regulated by Temperature and Light Conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef]
- Lin-Wang, K.; Micheletti, D.; Palmer, J.; Volz, R.; Lozano, L.; Espley, R.; Hellens, R.P.; Chagné, D.; Rowan, D.D.; Troggio, M.; et al. High Temperature Reduces Apple Fruit Colour via Modulation of the Anthocyanin Regulatory Complex. Plant Cell Environ. 2011, 34, 1176–1190. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Matthews, M.A.; Di Gaspero, G.; Gambetta, G.A. Water Deficits Accelerate Ripening and Induce Changes in Gene Expression Regulating Flavonoid Biosynthesis in Grape Berries. Planta 2007, 227, 101–112. [Google Scholar] [CrossRef]
- Koyama, K.; Goto-Yamamoto, N. Bunch Shading During Different Developmental Stages Affects the Phenolic Biosynthesis in Berry Skins of ‘Cabernet Sauvignon’ Grapes. J. Amer. Soc. Hort. Sci. 2008, 133, 743–753. [Google Scholar] [CrossRef]
- Cohen, S.D.; Tarara, J.M.; Gambetta, G.A.; Matthews, M.A.; Kennedy, J.A. Impact of Diurnal Temperature Variation on Grape Berry Development, Proanthocyanidin Accumulation, and the Expression of Flavonoid Pathway Genes. J. Exp. Bot. 2012, 63, 2655–2665. [Google Scholar] [CrossRef]
- Kiferle, C.; Fantini, E.; Bassolino, L.; Povero, G.; Spelt, C.; Buti, S.; Giuliano, G.; Quattrocchio, F.; Koes, R.; Perata, P.; et al. Tomato R2R3-MYB Proteins SlANT1 and SlAN2: Same Protein Activity, Different Roles. PLoS ONE 2015, 10, e0136365. [Google Scholar] [CrossRef]
- Zheng, J.; Wu, H.; Zhu, H.; Huang, C.; Liu, C.; Chang, Y.; Kong, Z.; Zhou, Z.; Wang, G.; Lin, Y.; et al. Determining Factors, Regulation System, and Domestication of Anthocyanin Biosynthesis in Rice Leaves. New Phytol. 2019, 223, 705–721. [Google Scholar] [CrossRef]
- Dong, N.-Q.; Lin, H.-X. Contribution of Phenylpropanoid Metabolism to Plant Development and Plant–Environment Interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Chen, X.; Wang, P.; Gu, M.; Lin, X.; Hou, B.; Zheng, Y.; Sun, Y.; Jin, S.; Ye, N. R2R3-MYB Transcription Factor Family in Tea Plant (Camellia Sinensis): Genome-Wide Characterization, Phylogeny, Chromosome Location, Structure and Expression Patterns. Genomics 2021, 113, 1565–1578. [Google Scholar] [CrossRef]
- Li, Y.; Liang, J.; Zeng, X.; Guo, H.; Luo, Y.; Kear, P.; Zhang, S.; Zhu, G. Genome-Wide Analysis of MYB Gene Family in Potato Provides Insights into Tissue-Specific Regulation of Anthocyanin Biosynthesis. Hortic. Plant J. 2021, 7, 129–141. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, Q.; Sun, Y.; Yang, L.; Wang, Z. Genome-Wide Identification and Characterization of R2R3-MYB Family in Hypericum Perforatum under Diverse Abiotic Stresses. Int. J. Biol. Macromol. 2020, 145, 341–354. [Google Scholar] [CrossRef]
- Liu, C.; Xie, T.; Chen, C.; Luan, A.; Long, J.; Li, C.; Ding, Y.; He, Y. Genome-Wide Organization and Expression Profiling of the R2R3-MYB Transcription Factor Family in Pineapple (Ananas Comosus). BMC Genom. 2017, 18, 503. [Google Scholar] [CrossRef]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB Gene Family in Arabidopsis Thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Pérez-Díaz, J.R.; Pérez-Díaz, J.; Madrid-Espinoza, J.; González-Villanueva, E.; Moreno, Y.; Ruiz-Lara, S. New Member of the R2R3-MYB Transcription Factors Family in Grapevine Suppresses the Anthocyanin Accumulation in the Flowers of Transgenic Tobacco. Plant Mol. Biol. 2016, 90, 63–76. [Google Scholar] [CrossRef]
- Xu, H.; Wang, N.; Liu, J.; Qu, C.; Wang, Y.; Jiang, S.; Lu, N.; Wang, D.; Zhang, Z.; Chen, X. The Molecular Mechanism Underlying Anthocyanin Metabolism in Apple Using the MdMYB16 and MdbHLH33 Genes. Plant Mol. Biol. 2017, 94, 149–165. [Google Scholar] [CrossRef]
- Anwar, M.; Wang, G.; Wu, J.; Waheed, S.; Allan, A.C.; Zeng, L. Ectopic Overexpression of a Novel R2R3-MYB, NtMYB2 from Chinese Narcissus Represses Anthocyanin Biosynthesis in Tobacco. Molecules 2018, 23, 781. [Google Scholar] [CrossRef]
- Peng, X.; Wood, C.L.; Blalock, E.M.; Chen, K.C.; Landfield, P.W.; Stromberg, A.J. Statistical Implications of Pooling RNA Samples for Microarray Experiments. BMC Bioinform. 2003, 4, 26. [Google Scholar] [CrossRef]
- Karp, N.A.; Lilley, K.S. Investigating Sample Pooling Strategies for DIGE Experiments to Address Biological Variability. Proteomics 2009, 9, 388–397. [Google Scholar] [CrossRef]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis Transcription Factor MYB12 Is a Flavonol-Specific Regulator of Phenylpropanoid Biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef]
- D’Angeli, S.; Falasca, G.; Matteucci, M.; Altamura, M.M. Cold Perception and Gene Expression Differ in Olea Europaea Seed Coat and Embryo during Drupe Cold Acclimation. New Phytol. 2013, 197, 123–138. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nature Protocols 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Muto, A.; Müller, C.T.; Bruno, L.; McGregor, L.; Ferrante, A.; Chiappetta, A.A.C.; Bitonti, M.B.; Rogers, H.J.; Spadafora, N.D. Fruit Volatilome Profiling through GC × GC-ToF-MS and Gene Expression Analyses Reveal Differences amongst Peach Cultivars in Their Response to Cold Storage. Sci. Rep. 2020, 10, 18333. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a Central Hub for Transcription Factors and Regulatory Interactions in Plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Katiyar, A.; Smita, S.; Lenka, S.K.; Rajwanshi, R.; Chinnusamy, V.; Bansal, K.C. Genome-Wide Classification and Expression Analysis of MYB Transcription Factor Families in Rice and Arabidopsis. BMC Genom. 2012, 13, 544. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Reference |
|---|---|---|---|
| OePAL | 5’-ACACATCCATCTTCCAAAAG-3’ | 5’-GTTCCCAGTTCTTCCCTTAC-3’ | [23] |
| Oe4CL | 5’-AAATTTAAAGGCTTCCAGGT-3’ | 5’-GCTTCTTCGGTAAGTTCAAA-3’ | [23] |
| OeCHS | 5’-GATTGGAACTCGATTTTCTG-3’ | 5’-GGACTTTCTCATCTCATCCA-3’ | [23] |
| OeCHI | 5’-AGGGTTCACGTATGGAGTG-3’ | 5’-TGCAAATACAATCTCAGCAG-3’ | [23] |
| OeF3H | 5’-TCCTCTGCCCGTGTGATAGT-3’ | 5’-AATCCGTGTGATGCAGTGAG-3’ | [23] |
| OeF3’H | 5’-GTGGCAGAAGCTGACCTACC-3’ | 5’-CGTAGAGCCCTTTGGAATGA-3’ | [23] |
| OeF3’5’H | 5’-AGTGGTCACCAATGGGATGT-3’ | 5’-CACATCAAACGTGGCTCATT-3’ | [23] |
| OeDFR | 5’-ATTTCAGGTGTTGGCTGAGG-3’ | 5’-ATTCCATATGGCCAGGTCAA-3’ | [21] |
| OeANS | 5’-GCATAGGGTCACTGTCAATGG-3’ | 5’-TCCTTACCATCATGGCCTTT-3’ | [23] |
| OeUFGT | 5’-AATGGCTTTGATGGAAGGTG-3’ | 5’-TTCACGCTGGCATAAACTCA-3’ | [21] |
| Oeu050989.1 | 5′-TGGTCAAATTACTTTTCGGGAAGT-3′ | 5’-TCTCGTGCCATGTGTAATTATGG-3’ | Our study |
| Oeu004741.1 | 5’-ACTGACAGACAGAACCCCATGA-3’ | 5’-ATCTGCAGGATTTGCATGAGAA-3’ | Our study |
| Oeu004739.1 | 5’-TCTCTTATTTTCAACCATACACTCTTCAA-3’ | 5’-AGAGCATGTTTGTTTCTCATTTTCA-3’ | Our study |
| OeCRY2 | 5′-GTCCTACAAGCTCGTCCTATG-3′ | 5′-CTTGTCGCAACTATGCAAGT-3′ | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrari, M.; Muto, A.; Bruno, L.; Muzzalupo, I.; Chiappetta, A. Modulation of Anthocyanin Biosynthesis-Related Genes during the Ripening of Olea europaea L. cvs Carolea and Tondina Drupes in Relation to Environmental Factors. Int. J. Mol. Sci. 2023, 24, 8770. https://doi.org/10.3390/ijms24108770
Ferrari M, Muto A, Bruno L, Muzzalupo I, Chiappetta A. Modulation of Anthocyanin Biosynthesis-Related Genes during the Ripening of Olea europaea L. cvs Carolea and Tondina Drupes in Relation to Environmental Factors. International Journal of Molecular Sciences. 2023; 24(10):8770. https://doi.org/10.3390/ijms24108770
Chicago/Turabian StyleFerrari, Michele, Antonella Muto, Leonardo Bruno, Innocenzo Muzzalupo, and Adriana Chiappetta. 2023. "Modulation of Anthocyanin Biosynthesis-Related Genes during the Ripening of Olea europaea L. cvs Carolea and Tondina Drupes in Relation to Environmental Factors" International Journal of Molecular Sciences 24, no. 10: 8770. https://doi.org/10.3390/ijms24108770