
Alternative Splicing Analysis Revealed the Role of Alpha-Linolenic Acid and Carotenoids in Fruit Development of Osmanthus fragrans
 ,
,
Abstract
:1. Introduction
2. Results
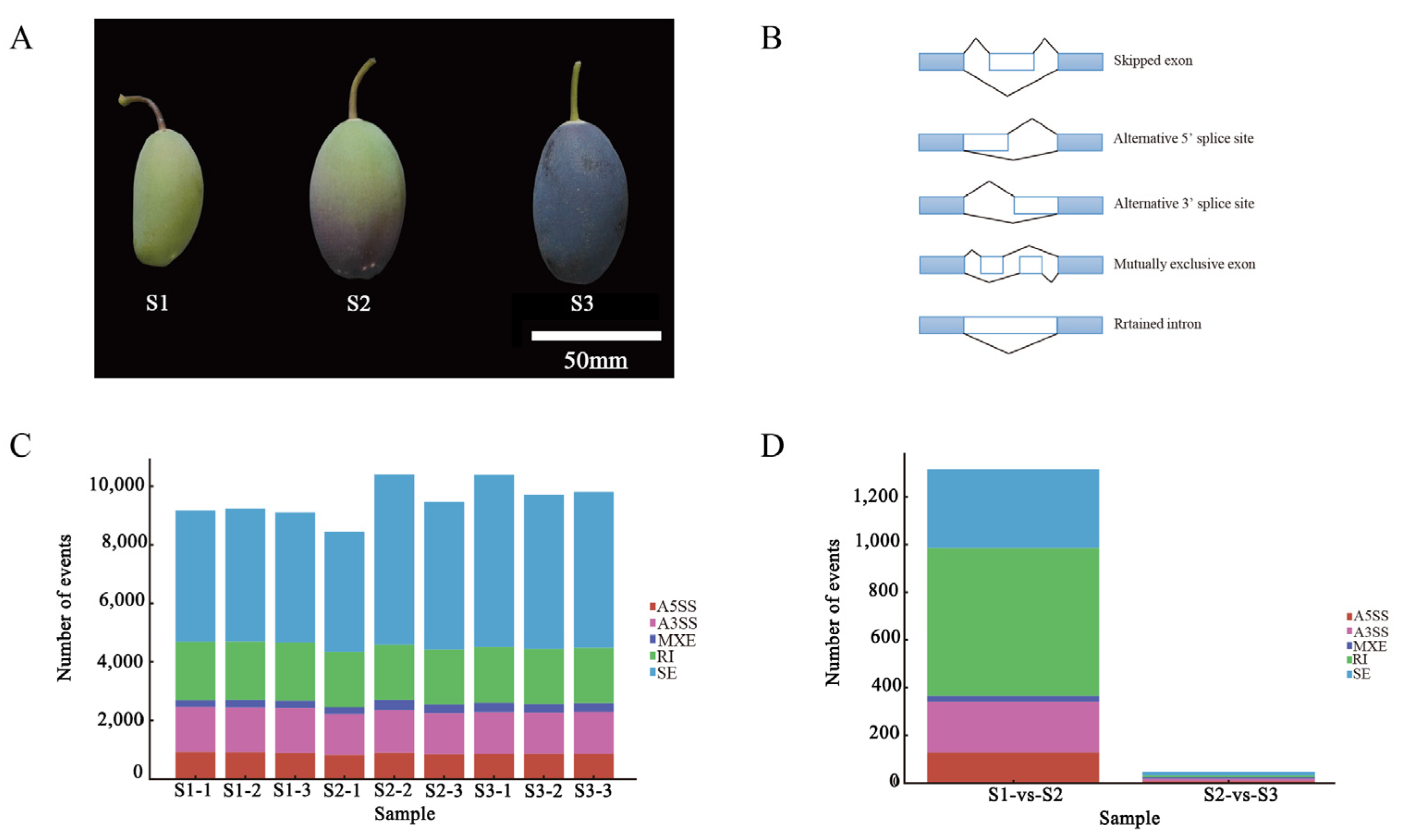
2.1. Phenotype and AS Event Statistics
2.2. Analysis of Differentially Expressed Alternative Splicing Events
2.3. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Enrichment Analysis
2.4. RT-PCR Validation of Differentially Expressed Alternative Splicing Genes
3. Discussion
3.1. Up-Regulation of Alpha-Linolenic Acid Metabolism in O. fragrans Fruit Development
3.2. Carotenoid Metabolism Affects the Color Formation of O. fragrans Fruit
4. Materials and Methods
4.1. Sample Selection and Preparation
4.2. Transcriptome Sequencing and Differential Alternative Splicing Analysis
4.3. RT-PCR Validation of AS Events
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kornblihtt, A.R.; Schor, I.E.; Alló, M.; Dujardin, G.; Petrillo, E.; Muñoz, M.J. Alternative splicing: A pivotal step between eukaryotic transcription and translation. Nat. Rev. Mol. Cell Biol. 2013, 14, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W. Why genes in pieces? Nature 1978, 271, 501. [Google Scholar] [CrossRef] [PubMed]
- Berget, S.M.; Moore, C.; Sharp, P.A. Spliced segments at the 5′ terminus of adenovirus 2 late mRNA. Proc. Natl. Acad. Sci. USA 1977, 74, 3171–3175. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.C.; Smith, C.W. Alternative splicing: Combinatorial output from the genome. Curr. Opin. Chem. Biol. 2002, 6, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.S.; Marquez, Y.; Kalyna, M.; Barta, A. Complexity of the Alternative Splicing Landscape in Plants. Plant Cell 2013, 25, 3657–3683. [Google Scholar] [CrossRef]
- McGuire, A.M.; Pearson, M.D.; Neafsey, D.E.; Galagan, J.E. Cross-kingdom patterns of alternative splicing and splice recognition. Genome Biol. 2008, 9, R50. [Google Scholar] [CrossRef]
- Bowler, E.; Oltean, S. Alternative Splicing in Angiogenesis. Int. J. Mol. Sci. 2019, 20, 2067. [Google Scholar] [CrossRef]
- Wajahat, M.; Bracken, C.P.; Orang, A. Emerging Functions for snoRNAs and snoRNA-Derived Fragments. Int. J. Mol. Sci. 2021, 22, 10193. [Google Scholar] [CrossRef]
- Aísa-Marín, I.; García-Arroyo, R.; Mirra, S.; Marfany, G. The Alter Retina: Alternative Splicing of Retinal Genes in Health and Disease. Int. J. Mol. Sci. 2021, 22, 1855. [Google Scholar] [CrossRef]
- Chen, M.-X.; Zhu, F.-Y.; Wang, F.-Z.; Ye, N.-H.; Gao, B.; Chen, X.; Zhao, S.-S.; Fan, T.; Cao, Y.-Y.; Liu, T.-Y.; et al. Alternative splicing and translation play important roles in hypoxic germination in rice. J. Exp. Bot. 2018, 70, 817–833. [Google Scholar] [CrossRef]
- Kufel, J.; Diachenko, N.; Golisz, A. Alternative splicing as a key player in the fine-tuning of the immunity response in Arabidopsis. Mol. Plant Pathol. 2022, 23, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- Lam, P.Y.; Wang, L.; Lo, C.; Zhu, F.-Y. Alternative Splicing and Its Roles in Plant Metabolism. Int. J. Mol. Sci. 2022, 23, 7355. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-Y.; Chen, M.-X.; Ye, N.-H.; Shi, L.; Ma, K.-L.; Yang, J.-F.; Cao, Y.-Y.; Zhang, Y.; Yoshida, T.; Fernie, A.R.; et al. Proteogenomic analysis reveals alternative splicing and translation as part of the abscisic acid response in Arabidopsis seedlings. Plant J. 2017, 91, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Patthy, L. Exon skipping-rich transcriptomes of animals reflect the significance of exon-shuffling in metazoan proteome evolution. Biol. Direct 2019, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Parmar, R.; Seth, R.; Sharma, R.K. Genome-wide identification and characterization of functionally relevant microsatellite markers from transcription factor genes of Tea (Camellia sinensis (L.) O. Kuntze). Sci. Rep. 2022, 1, 201. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Gardner, K.M.; Richards, C.; Chao, C.T.; Schwaninger, H.R.; Fazio, G.; Zhong, G.-Y.; Myles, S. Genomic consequences of apple improvement. Hortic. Res. 2021, 8, 9. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Monaco, C.M.; Proudfoot, R.; Miotto, P.M.; Herbst, E.A.F.; MacPherson, R.E.K.; Holloway, G.P. α-linolenic acid supplementation prevents exercise-induced improvements in white adipose tissue mitochondrial bioenergetics and whole-body glucose homeostasis in obese Zucker rats. Diabetologia 2018, 2, 433–444. [Google Scholar] [CrossRef]
- Goyal, A.; Tanwar, B.; Sihag, M.K.; Sharma, V. Sacha inchi (Plukenetia volubilis L.): An emerging source of nutrients, omega-3 fatty acid and phytochemicals. Food Chem. 2022, 373, 131459. [Google Scholar] [CrossRef]
- Gfeller, A.; Dubugnon, L.; Liechti, R.; Farmer, E.E. Jasmonate Biochemical Pathway. Sci. Signal. 2010, 109, cm3. [Google Scholar] [CrossRef]
- Caracciolo, G.; Magri, A.; Petriccione, M.; Maltoni, M.L.; Baruzzi, G. Influence of Cold Storage on Pear Physico-Chemical Traits and Antioxidant Systems in Relation to Superficial Scald Development. Foods 2020, 9, 1175. [Google Scholar] [CrossRef] [PubMed]
- Boratyński, F.; Szczepańska, E.; De Simeis, D.; Serra, S.; Brenna, E. Bacterial Biotransformation of Oleic Acid: New Findings on the Formation of γ-Dodecalactone and 10-Ketostearic Acid in the Culture of Micrococcus luteus. Molecules 2020, 13, 3024. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros, R.A.B.; Júnior, E.V.D.S.; Barros, Z.M.P.; da Silva, J.H.F.; Brandão, S.C.R.; Azoubel, P.M. Convective drying of mango enriched with phenolic compounds from grape residue flour under different impregnation methods. Food Res. Int. 2022, 158, 111539. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, A.; Adiletta, G.; Di Matteo, M.; Panfili, G.; Niro, S.; Gentile, C.; Farina, V.; Cinquanta, L.; Corona, O. Evolution of Carotenoid Content, Antioxidant Activity and Volatiles Compounds in Dried Mango Fruits (Mangifera Indica L.). Foods 2020, 10, 1424. [Google Scholar] [CrossRef]
- Maldonado-Celis, M.E.; Yahia, E.M.; Bedoya, R.; Landázuri, P.; Loango, N.; Aguillón, J.; Restrepo, B.; Ospina, J.C.G. Chemical Composition of Mango (Mangifera indica L.) Fruit: Nutritional and Phytochemical Compounds. Front. Plant Sci. 2019, 10, 1073. [Google Scholar] [CrossRef]
- Proctor, M.S.; Morey-Burrows, F.S.; Canniffe, D.P.; Martin, E.C.; Swainsbury, D.J.K.; Johnson, M.P.; Hunter, C.N.; Sutherland, G.A.; Hitchcock, A. Zeta-Carotene Isomerase (Z-ISO) Is Required for Light-Independent Carotenoid Biosynthesis in the Cyanobacterium Synechocystis sp. PCC 6803. Microorganisms 2022, 9, 1730. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Lado, J.; Alós, E.; Alquézar, B.; Dery, O.; Hirschberg, J.; Zacarías, L. A mutant allele of ζ-carotene isomerase (Z-ISO) is associated with the yellow pigmentation of the “Pinalate” sweet orange mutant and reveals new insights into its role in fruit carotenogenesis. BMC Plant Biol. 2019, 19, 465. [Google Scholar] [CrossRef] [PubMed]
- Efremov, G.I.; Shchennikova, A.V.; Kochieva, E.Z. Characterization of 15-cis-ζ-Carotene Isomerase Z-ISO in Cultivated and Wild Tomato Species Differing in Ripe Fruit Pigmentation. Plants 2021, 11, 2365. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.F.S.; Chen, C.; Gmitter, F.G.; Moore, G.A.; Costa, M.G.C. Expression and phylogenetic analysis of two new lycopene β-cyclases from Citrus paradisi. Physiol. Plant. 2010, 141, 1–10. [Google Scholar] [CrossRef]
- Fanciullino, A.L.; Cerćos, M.; Dhique-Maye, C.; Froelicher, Y.; Talón, M.; Ollitrault, P.; Morillon, R. Changes in carotenoid content and biosynthetic gene expression in juice sacs of four orange varieties (Citrus sinensis) differing in flesh fruit color. J. Agric. Food Chem. 2008, 10, 3628–3638. [Google Scholar] [CrossRef]
- Nakkanong, K.; Yang, J.H.; Zhang, M.F. Carotenoid Accumulation and Carotenogenic Gene Expression during Fruit Development in Novel Interspecific Inbred Squash Lines and Their Parents. J. Agric. Food Chem. 2012, 60, 5936–5944. [Google Scholar] [CrossRef] [PubMed]
- Bunting, E.L.; Wanyama, D.; Goodwin, R.; Weil, N.; Sabbatini, P.; Andresen, J. Vitis vinifera Production in Michigan: Factors and Trends Driving Cultivation Patterns. Front. Plant Sci. 2021, 12, 704690. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, K.; Hirvelä, E.; Nevala, T.; Sipari, N.; Suokas, M.; Jaakola, L. Changes in the abscisic acid levels and related gene expression during fruit development and ripening in bilberry (Vaccinium myrtillus L.). Phytochemistry 2013, 95, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, N.; Armbruster, U.; Iven, T.; Brückle, L.; Melzer, M.; Feussner, I.; Jahns, P. Tissue-Specific Accumulation and Regulation of Zeaxanthin Epoxidase in Arabidopsis Reflect the Multiple Functions of the Enzyme in Plastids. Plant Cell Physiol. 2015, 2, 346–357. [Google Scholar] [CrossRef]
- Wolters, A.-M.A.; Uitdewilligen, J.G.A.M.L.; Kloosterman, B.A.; Hutten, R.C.B.; Visser, R.G.F.; van Eck, H.J. Identification of alleles of carotenoid pathway genes important for zeaxanthin accumulation in potato tubers. Plant Mol. Biol. 2010, 6, 659–671. [Google Scholar] [CrossRef]
- Cruet-Burgos, C.; Cox, S.; Ioerger, B.P.; Perumal, R.; Hu, Z.; Herald, T.J.; Bean, S.R.; Rhodes, D.H. Advancing provitamin A biofortification in sorghum: Genome-wide association studies of grain carotenoids in global germplasm. Plant Genome 2020, 13, e20013. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jang, S.J.; Jeong, H.B.; Lee, S.Y.; Venkatesh, J.; Lee, J.H.; Kwon, J.K.; Kang, B.C. A mutation in Zeaxanthin epoxidase contributes to orange coloration and alters carotenoid contents in pepper fruit (Capsicum annuum). Plant J. 2021, 106, 1692–1707. [Google Scholar] [CrossRef]
- Wang, X.Y.; Zhang, C.; Zhang, M.; Ma, C.C.; Yan, J.P.; Duan, Y.F. Study of anthocyanin composition and fruit color of Osmanthus fragrans ‘Zi Yingui’. Plant Sci. J. 2022, 5, 569–579. [Google Scholar]
- Mehmood, A.; Laiho, A.; Venäläinen, M.S.; McGlinchey, A.J.; Wang, N.; Elo, L.L. Systematic evaluation of differential splicing tools for RNA-seq studies. Briefings Bioinform. 2020, 6, 2052–2065. [Google Scholar] [CrossRef]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef]
- Berlivet, S.; Guiraud, V.; Houlard, M.; Gérard, M. pHYPER, a shRNA vector for high-efficiency RNA interference in embryonic stem cells. Biotechniques 2007, 6, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Woodcroft, B.J.; Boyd, J.A.; Tyson, G.W. OrfM: A fast open reading frame predictor for metagenomic data. Bioinformatics 2016, 32, 2702–2703. [Google Scholar] [CrossRef] [PubMed]
- Rehmsmeier, M.; Vingron, M. Phylogenetic information improves homology detection. Proteins Struct. Funct. Bioinform. 2001, 45, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Brhane, H.; Haileselassie, T.; Tesfaye, K.; Ortiz, R.; Hammenhag, C.; Abreha, K.B.; Vetukuri, R.R.; Geleta, M. Finger millet RNA-seq reveals differential gene expression associated with tolerance to aluminum toxicity and provides novel genomic resources. Front. Plant Sci. 2022, 13, 1068383. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoform | Gene | Gene-Family | S1 | S2 | S3 |
|---|---|---|---|---|---|
| BGI_novel_T016400 | BGI_novel_G002195 | LOX | 6701.00 | 12,170.67 | 10,720.67 |
| BGI_novel_T016401 | BGI_novel_G002196 | LOX | 9670.33 | 27,281.33 | 22,006.67 |
| BGI_novel_T016402 | BGI_novel_G002197 | LOX | 845.00 | 3869.33 | 4713.33 |
| BGI_novel_T016404 | BGI_novel_G002199 | LOX | 13,446.33 | 27,287.33 | 22,494.00 |
| BGI_novel_T016405 | BGI_novel_G002200 | LOX | 10,720.33 | 22,245.33 | 18,129.33 |
| BGI_novel_T009589 | ofr.gene19109 | LOX | 35,641.67 | 145,639.67 | 129,475.33 |
| ofr.gene19109-mRNA-1 | ofr.gene19109 | LOX | 17.33 | 8.33 | 53.33 |
| BGI_novel_T021032 | ofr.gene24663 | LOX | 79,012.67 | 330,175.00 | 522,090.33 |
| ofr.gene24663-mRNA-1 | ofr.gene24663 | LOX | 1.00 | 25.33 | 19.00 |
| ofr.gene24665-mRNA-1 | ofr.gene24665 | LOX | 20.67 | 228.33 | 691.33 |
| BGI_novel_T021033 | ofr.gene24865 | LOX | 320.67 | 1232.33 | 1758.67 |
| BGI_novel_T021034 | ofr.gene24865 | LOX | 801.00 | 6089.00 | 9685.67 |
| BGI_novel_T021035 | ofr.gene24865 | LOX | 6975.33 | 39,254.00 | 72,009.33 |
| BGI_novel_T021036 | ofr.gene24865 | LOX | 55.67 | 301.33 | 415.33 |
| ofr.gene24865-mRNA-1 | ofr.gene24865 | LOX | 79.33 | 286.33 | 3385.00 |
| BGI_novel_T023460 | ofr.gene34384 | LOX | 16.33 | 647.33 | 1578.33 |
| ofr.gene34384-mRNA-1 | ofr.gene34384 | LOX | 1.00 | 1.00 | 1.00 |
| ofr.gene47520-mRNA-1 | ofr.gene47520 | LOX | 12,844.67 | 2456.33 | 2501.00 |
| BGI_novel_T018820 | ofr.gene10951 | AOS | 2040.33 | 294.00 | 536.00 |
| ofr.gene10951-mRNA-1 | ofr.gene10951 | AOS | 2998.33 | 176.67 | 196.33 |
| ofr.gene12088-mRNA-1 | ofr.gene12088 | AOS | 70.67 | 1040.33 | 759.33 |
| ofr.gene53671-mRNA-1 | ofr.gene53671 | AOS | 14,221.00 | 46,525.67 | 40,584.67 |
| ofr.gene3545-mRNA-1 | ofr.gene3545 | OPR | 1682.00 | 6327.67 | 4652.67 |
| ofr.gene3636-mRNA-1 | ofr.gene3636 | OPR | 1753.00 | 7861.33 | 6424.67 |
| BGI_novel_T003932 | ofr.gene57358 | OPR | 13,911.00 | 163,878.67 | 141,732.00 |
| ofr.gene57358-mRNA-1 | ofr.gene57358 | OPR | 226.00 | 567.67 | 531.67 |
| Isoform | Gene | Gene-Family | S1 | S2 | S3 |
|---|---|---|---|---|---|
| BGI_novel_T017069 | ofr.gene29216 | Z-ISO | 6647.67 | 1403.00 | 1603.67 |
| BGI_novel_T017070 | ofr.gene29242 | Z-ISO | 314.67 | 189.67 | 213.67 |
| ofr.gene29242-mRNA-1 | ofr.gene29242 | Z-ISO | 4735.67 | 820.67 | 1020.33 |
| ofr.gene42137-mRNA-1 | ofr.gene42137 | LCYE | 910.00 | 19.33 | 33.33 |
| ofr.gene8214-mRNA-1 | ofr.gene8214 | LCYE | 910.00 | 19.33 | 33.33 |
| BGI_novel_T016725 | ofr.gene33284 | LCYB | 1004.33 | 55.67 | 108.67 |
| ofr.gene33284-mRNA-1 | ofr.gene33284 | LCYB | 14.67 | 15.33 | 11.67 |
| BGI_novel_T018783 | ofr.gene11033 | ZEP | 1215.33 | 29.00 | 84.33 |
| BGI_novel_T018237 | ofr.gene12387 | ZEP | 7590.00 | 1044.67 | 629.33 |
| BGI_novel_T001884 | ofr.gene23734 | ZEP | 9843.33 | 54.00 | 370.33 |
| BGI_novel_T001885 | ofr.gene23734 | ZEP | 579.00 | 197.67 | 29.33 |
| BGI_novel_T017743 | ofr.gene30820 | ZEP | 1109.33 | 167.67 | 144.67 |
| BGI_novel_T008623 | ofr.gene55834 | ZEP | 3569.00 | 1294.00 | 1092.00 |
| ofr.gene55834-mRNA-1 | ofr.gene55834 | ZEP | 1727.33 | 533.33 | 610.00 |
| ofr.gene8564-mRNA-1 | ofr.gene8564 | ZEP | 2680.33 | 7073.67 | 4566.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Zhang, C.; Wang, X.; Zhu, F.; Wang, X.; Zhang, M.; Duan, Y. Alternative Splicing Analysis Revealed the Role of Alpha-Linolenic Acid and Carotenoids in Fruit Development of Osmanthus fragrans. Int. J. Mol. Sci. 2023, 24, 8666. https://doi.org/10.3390/ijms24108666
Ma C, Zhang C, Wang X, Zhu F, Wang X, Zhang M, Duan Y. Alternative Splicing Analysis Revealed the Role of Alpha-Linolenic Acid and Carotenoids in Fruit Development of Osmanthus fragrans. International Journal of Molecular Sciences. 2023; 24(10):8666. https://doi.org/10.3390/ijms24108666
Chicago/Turabian StyleMa, Cancan, Cheng Zhang, Xiaoyan Wang, Fuyuan Zhu, Xianrong Wang, Min Zhang, and Yifan Duan. 2023. "Alternative Splicing Analysis Revealed the Role of Alpha-Linolenic Acid and Carotenoids in Fruit Development of Osmanthus fragrans" International Journal of Molecular Sciences 24, no. 10: 8666. https://doi.org/10.3390/ijms24108666