
Genome-Wide Analysis of the BBX Genes in Platanus × acerifolia and Their Relationship with Flowering and/or Dormancy
Abstract
:1. Introduction
2. Results
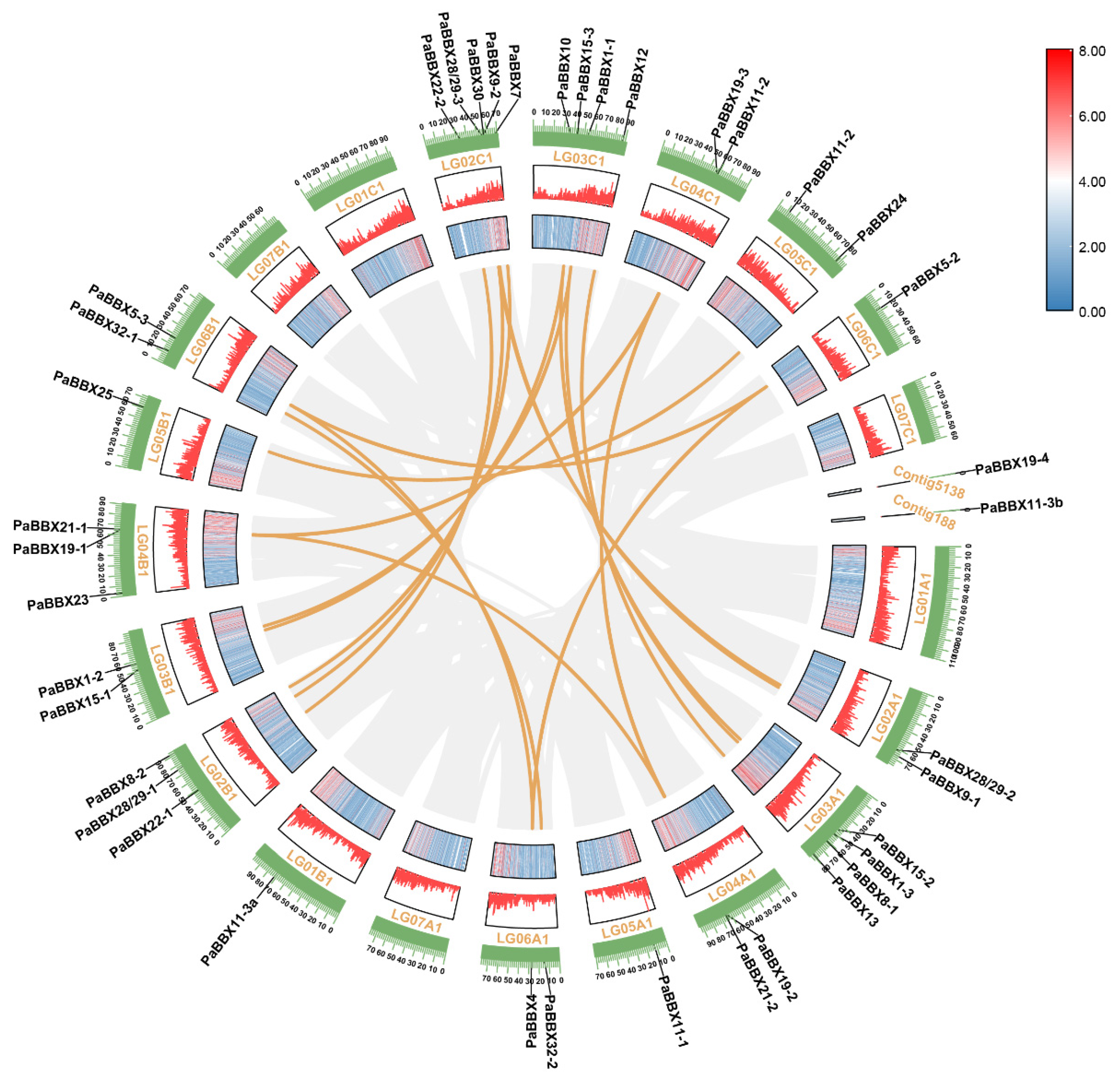
2.1. Identification of BBX Genes in P. × acerifolia
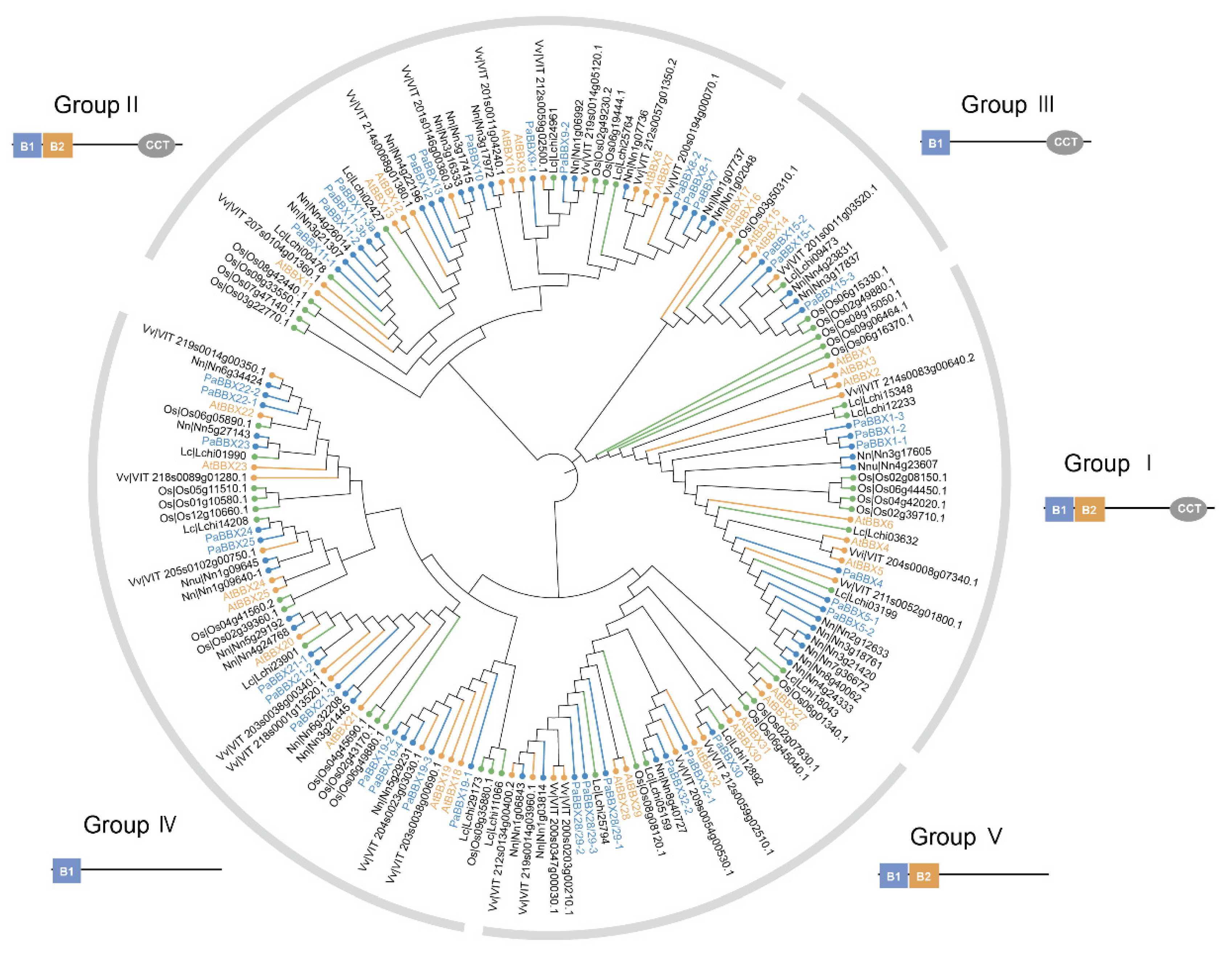
2.2. Phylogenetic Analysis and Multiple Alignments of BBX Proteins
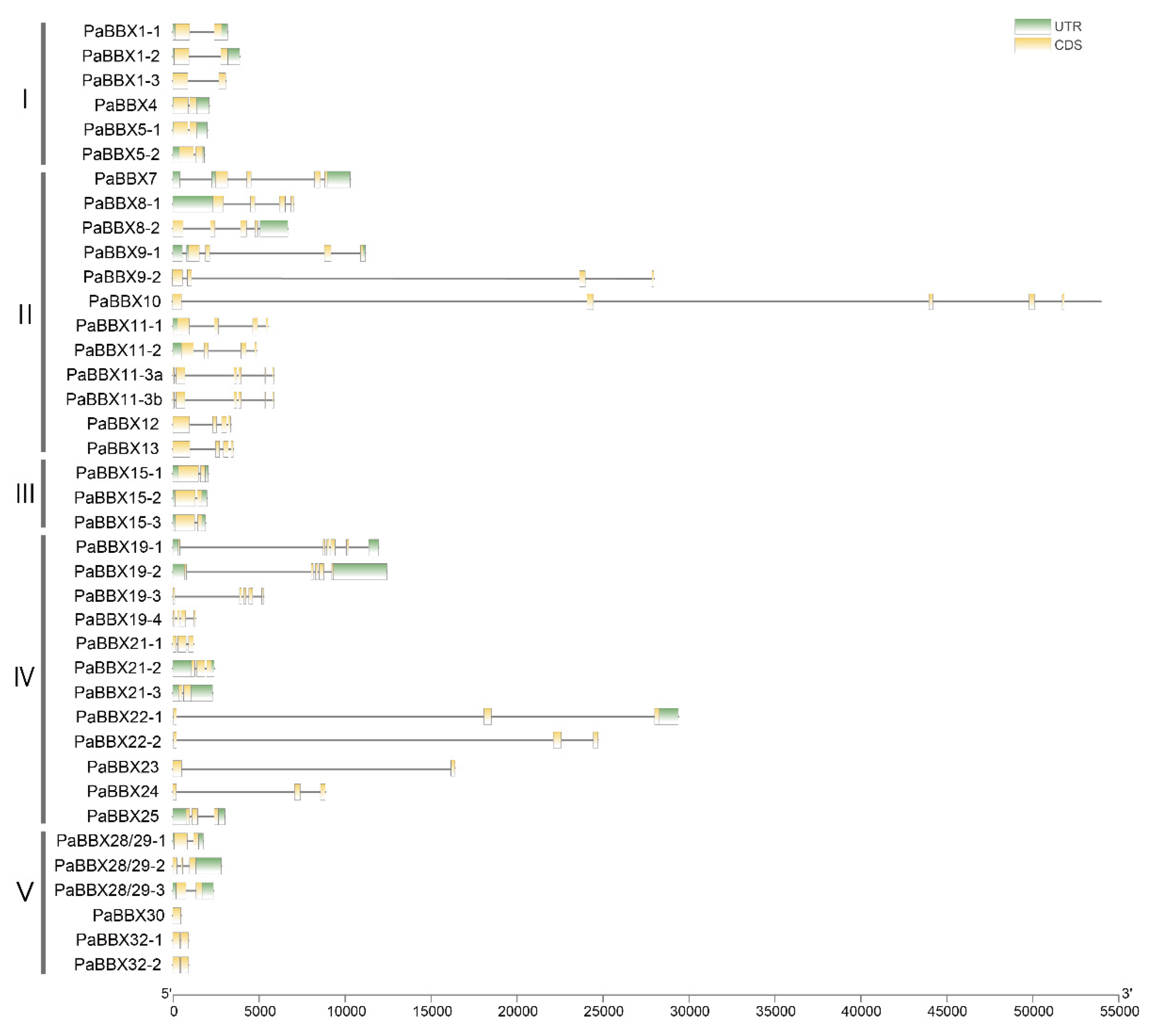
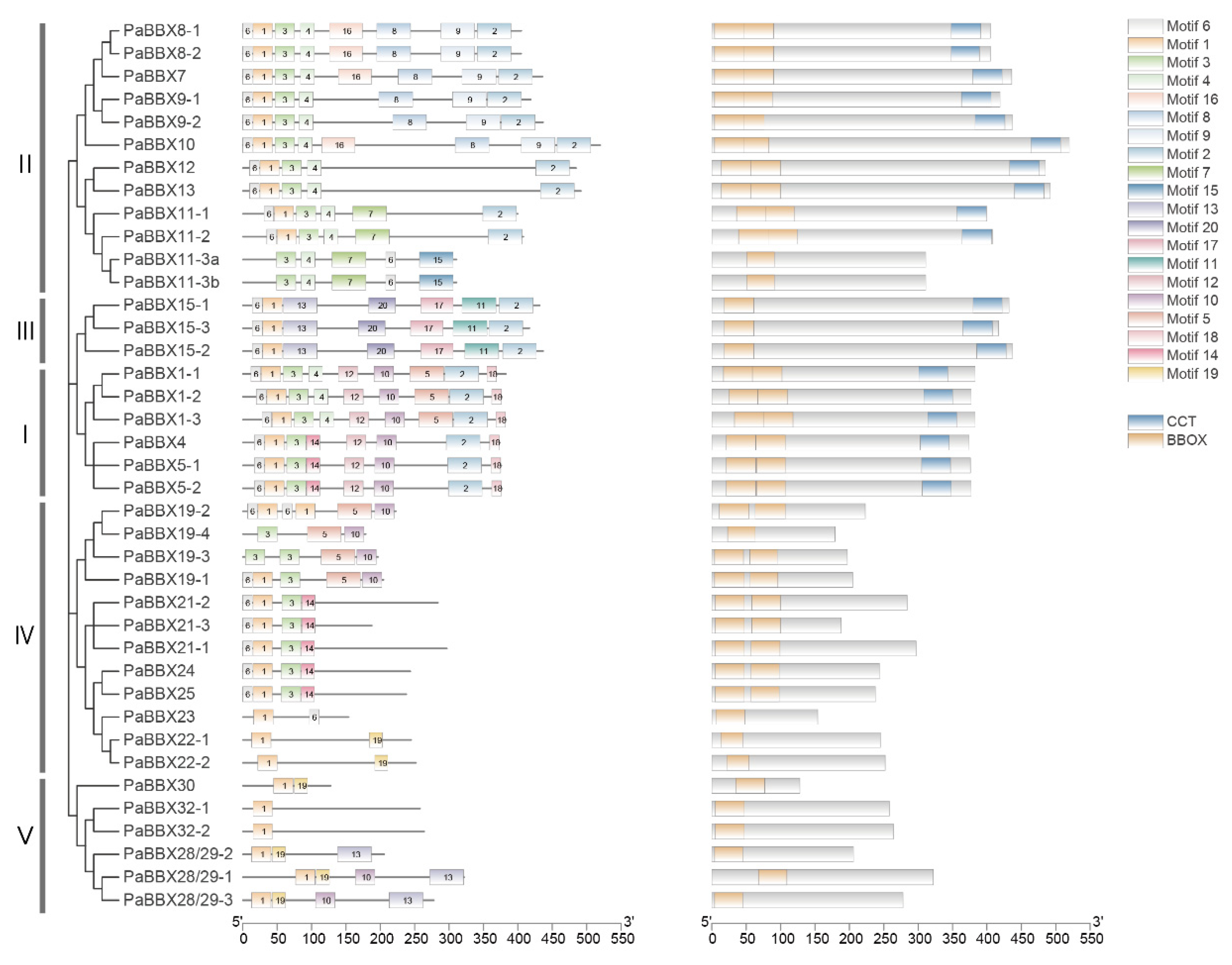
2.3. Gene Structure and Conserved Motif Analysis
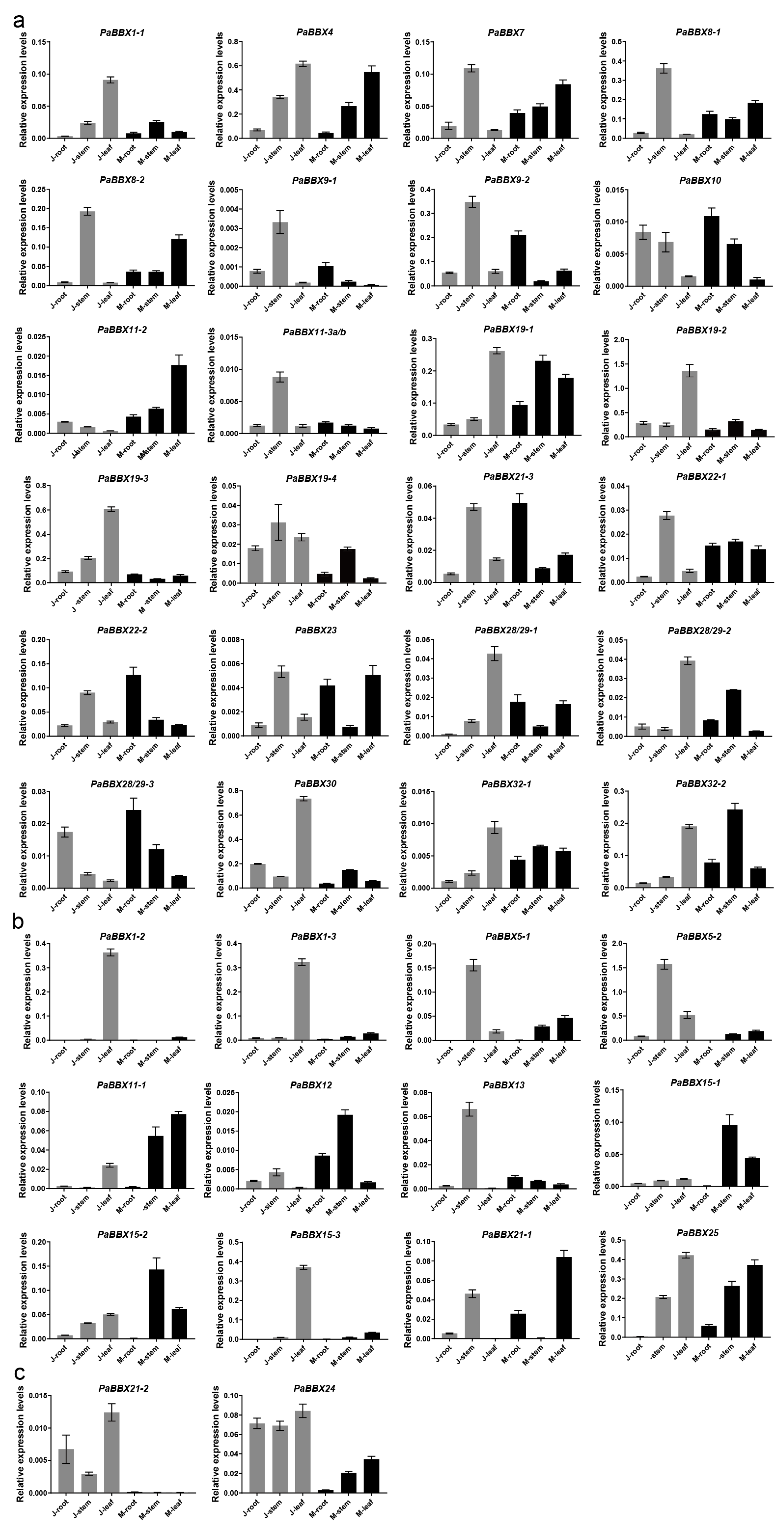
2.4. Expression Profiles of PaBBX Genes
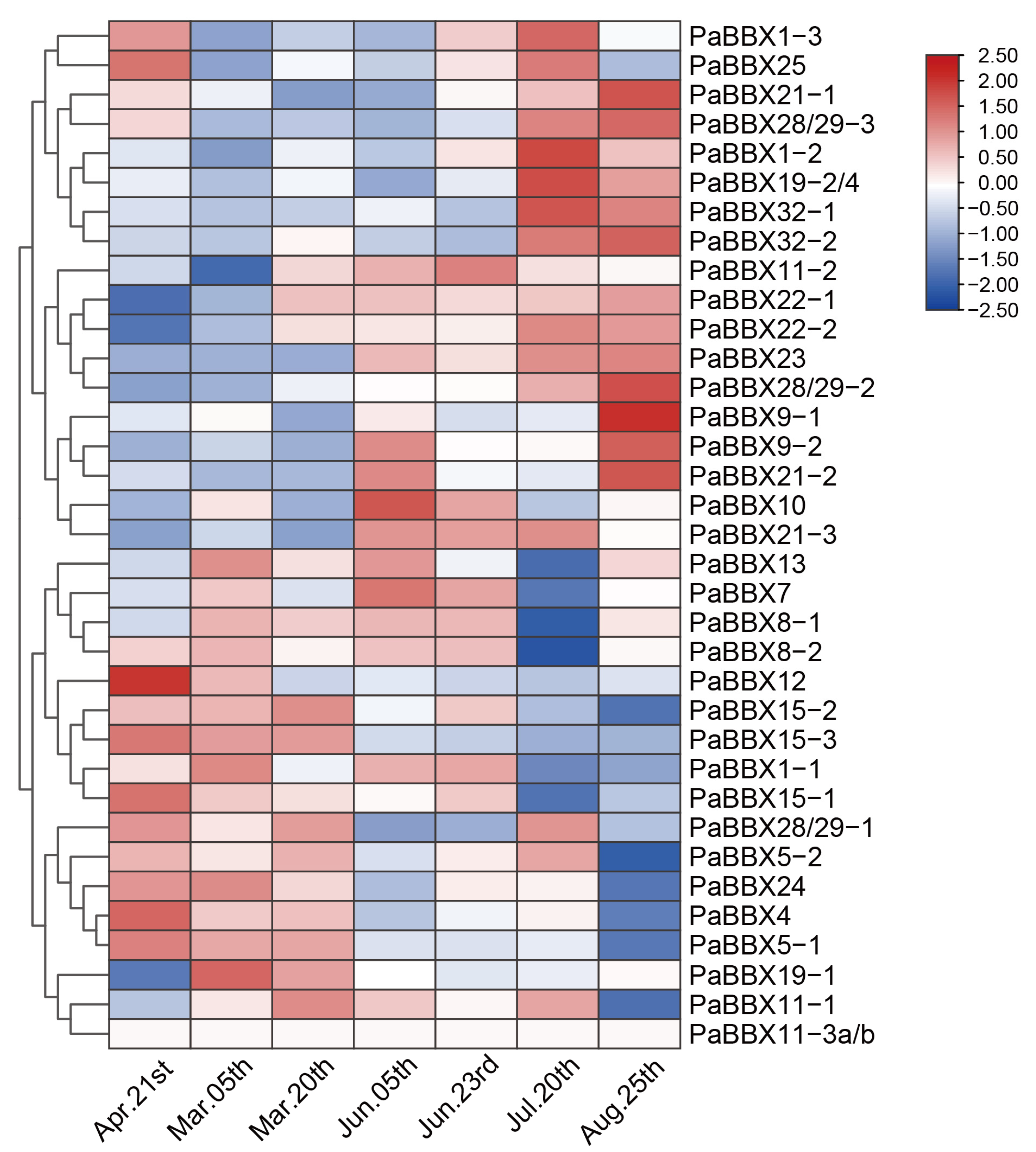
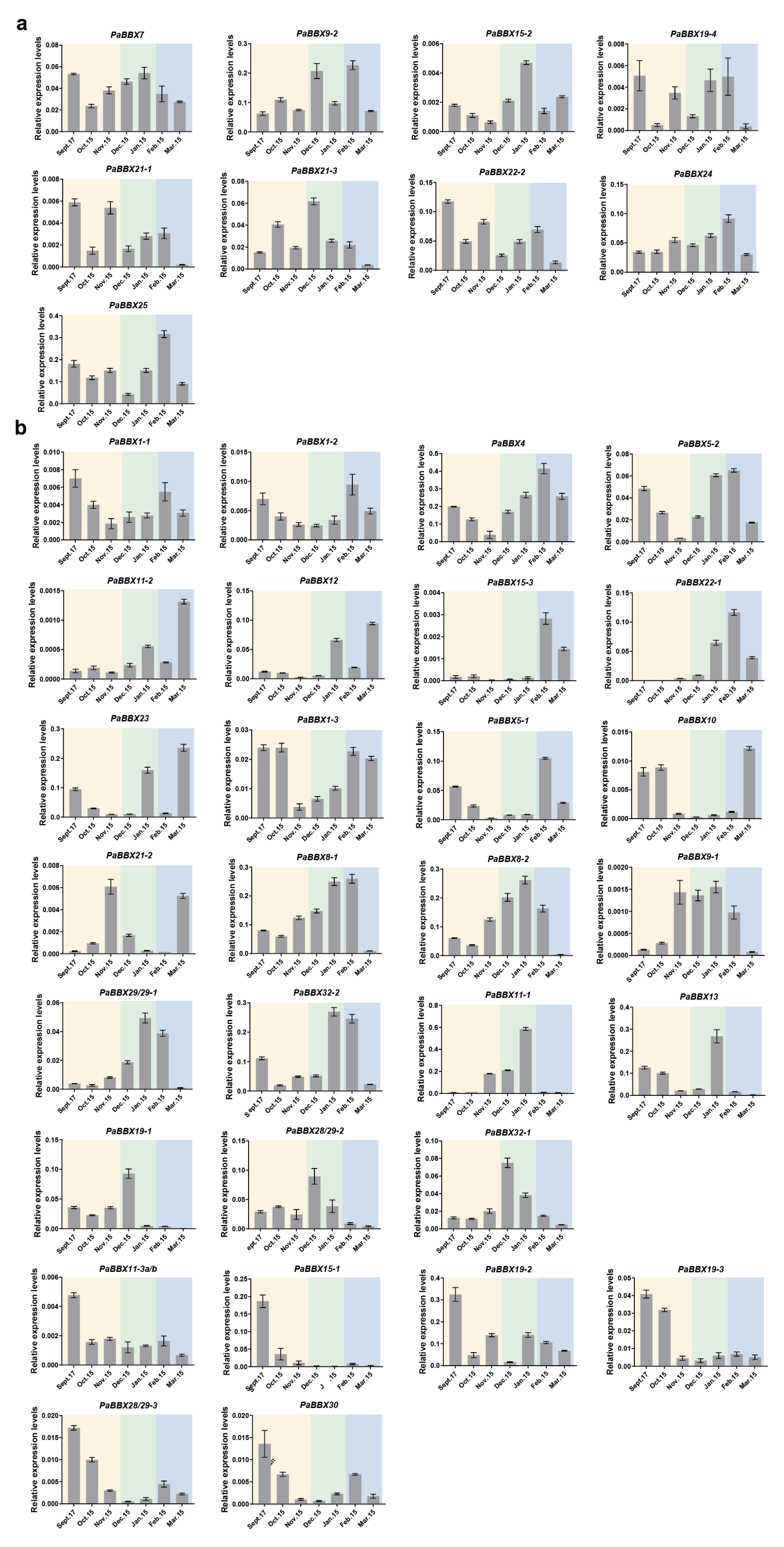
2.5. Transcriptional Profiles of PaBBX Genes during Annual
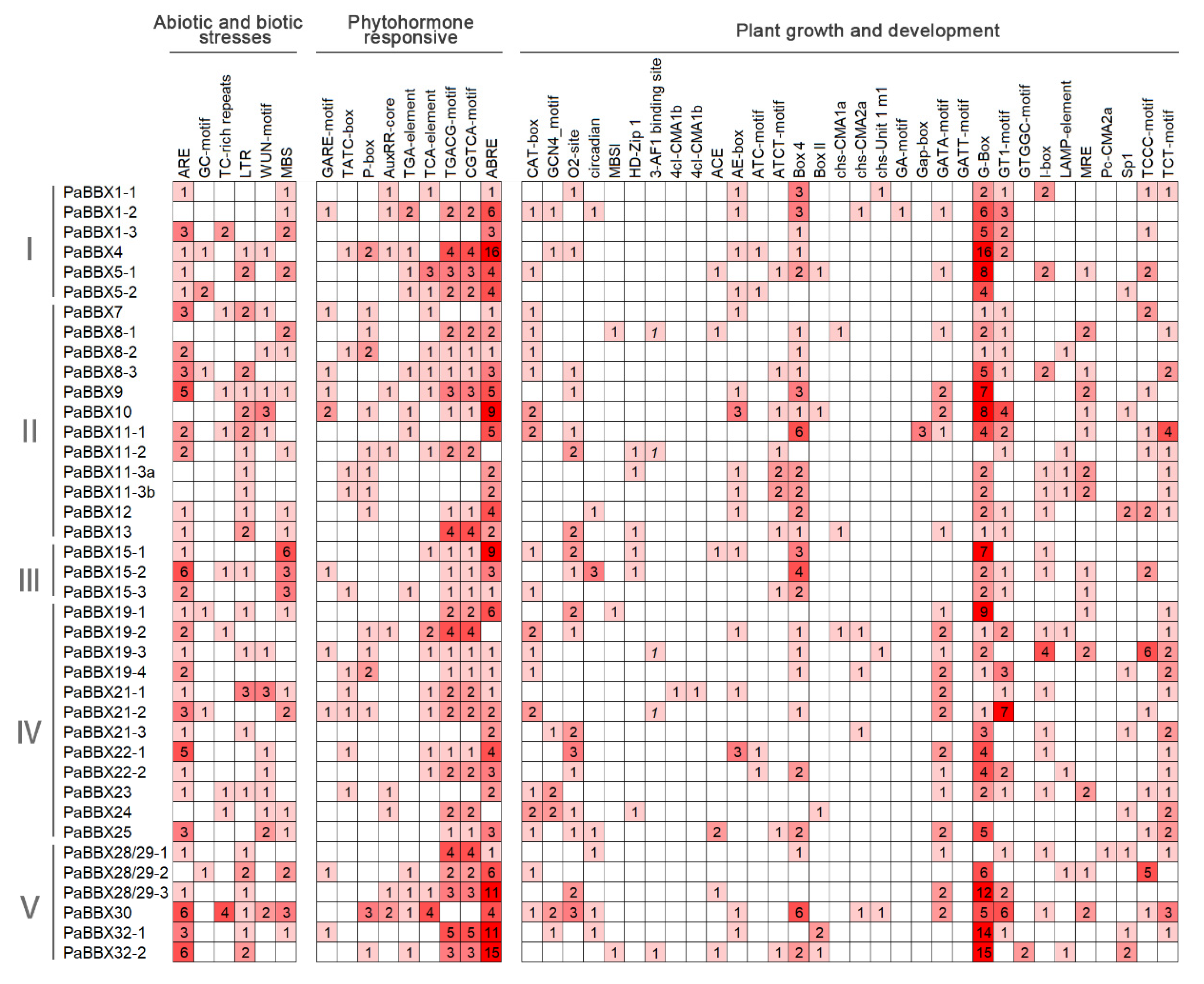
2.6. Identification of Cis-Elements in the Promoter of PaBBX Genes
3. Discussion
3.1. Structural and Evolutionary Analysis of BBX Genes in P. × acerifolia
3.2. Expression Patterns and Potential Functions of PaBBX Genes
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of BBX Genes from P. × acerifolia
4.3. Phylogenetic Analysis and Protein Sequence Alignments
4.4. Gene Structure and Conserved Motif Analysis
4.5. Putative Cis-Element Analysis of PaBBX Genes Promoters and Intron
4.6. Cloning PaBBX22-2 and PaBBX23
4.7. Analysis Expression Pattern of PaBBX Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kumagai, T.; Ito, S.; Nakamichi, N.; Niwa, Y.; Murakami, M.; Yamashino, T.; Mizuno, T. The common function of a novel subfamily of B-Box zinc finger proteins with reference to circadian-associated events in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2008, 72, 1539–1549. [Google Scholar] [CrossRef]
- Khanna, R.; Kronmiller, B.; Maszle, D.R.; Coupland, G.; Holm, M.; Mizuno, T.; Wu, S.-H. The Arabidopsis B-box zinc finger family. Plant Cell 2009, 21, 3416–3420. [Google Scholar] [CrossRef]
- Kluska, K.; Adamczyk, J.; Krężel, A. Metal binding properties, stability and reactivity of zinc fingers. Coord. Chem. Rev. 2018, 367, 18–64. [Google Scholar] [CrossRef]
- Crocco, C.D.; Botto, J.F. BBX proteins in green plants: Insights into their evolution, structure, feature and functional diversification. Gene 2013, 531, 44–52. [Google Scholar] [CrossRef]
- Wang, C.-Q.; Guthrie, C.; Sarmast, M.K.; Dehesh, K. BBX19 interacts with CONSTANS to repress FLOWERING LOCUS T transcription, defining a flowering time checkpoint in Arabidopsis. Plant Cell 2014, 26, 3589–3602. [Google Scholar] [CrossRef]
- Xu, D.; Jiang, Y.; Li, J.; Holm, M.; Deng, X.W. The B-Box Domain Protein BBX21 Promotes Photomorphogenesis. Plant Physiol. 2018, 176, 2365–2375. [Google Scholar] [CrossRef]
- Datta, S.; Johansson, H.; Hettiarachchi, C.; Irigoyen, M.L.; Desai, M.; Rubio, V.; Holm, M. LZF1/SALT TOLERANCE HOMOLOG3, an Arabidopsis B-box protein involved in light-dependent development and gene expression, undergoes COP1-mediated ubiquitination. Plant Cell 2008, 20, 2324–2338. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, G.; Yin, C.; Fang, Y. B-box transcription factor 28 regulates flowering by interacting with constans. Sci. Rep. 2020, 10, 17789. [Google Scholar] [CrossRef]
- Robson, F.; Costa, M.M.; Hepworth, S.R.; Vizir, I.; Piñeiro, M.; Reeves, P.H.; Putterill, J.; Coupland, G. Functional importance of conserved domains in the flowering-time gene CONSTANS demonstrated by analysis of mutant alleles and transgenic plants. Plant J. 2001, 28, 619–631. [Google Scholar] [CrossRef]
- Yadukrishnan, P.; Job, N.; Johansson, H.; Datta, S. Opposite roles of group IV BBX proteins: Exploring missing links between structural and functional diversity. Plant Signal. Behav. 2018, 13, e1462641. [Google Scholar] [CrossRef]
- Datta, S.; Hettiarachchi, G.H.C.M.; Deng, X.-W.; Holm, M. Arabidopsis CONSTANS-LIKE3 is a positive regulator of red light signaling and root growth. Plant Cell 2006, 18, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Hoecker, U. The activities of the E3 ubiquitin ligase COP1/SPA, a key repressor in light signaling. Curr. Opin. Plant Biol. 2017, 37, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Xu, D. COP1 and BBXs-HY5-mediated light signal transduction in plants. New Phytol. 2020, 228, 1748–1753. [Google Scholar] [CrossRef]
- Song, Z.; Bian, Y.; Liu, J.; Sun, Y.; Xu, D. B-box proteins: Pivotal players in light-mediated development in plants. J. Integr. Plant Biol. 2020, 62, 1293–1309. [Google Scholar] [CrossRef]
- Zhou, S.; Zhu, S.; Cui, S.; Hou, H.; Wu, H.; Hao, B.; Cai, L.; Xu, Z.; Liu, L.; Jiang, L.; et al. Transcriptional and post-transcriptional regulation of heading date in rice. New Phytol. 2021, 230, 943–956. [Google Scholar] [CrossRef]
- Campoli, C.; Drosse, B.; Searle, I.; Coupland, G.; von Korff, M. Functional characterisation of HvCO1, the barley (Hordeum vulgare) flowering time ortholog of CONSTANS. Plant J. 2012, 69, 868–880. [Google Scholar] [CrossRef]
- González-Schain, N.D.; Díaz-Mendoza, M.; Żurczak, M.; Suárez-López, P. Potato CONSTANS is involved in photoperiodic tuberization in a graft-transmissible manner. Plant J. 2012, 70, 678–690. [Google Scholar] [CrossRef]
- Abelenda, J.A.; Cruz-Oró, E.; Franco-Zorrilla, J.M.; Prat, S. Potato StCONSTANS-like1 suppresses storage organ formation by directly activating the FT-like StSP5G repressor. Curr. Biol. 2016, 26, 872–881. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, C.; Xu, Y.; Wei, Q.; Imtiaz, M.; Lan, H.; Gao, S.; Cheng, L.; Wang, M.; Fei, Z.; et al. A Zinc Finger Protein Regulates Flowering Time and Abiotic Stress Tolerance in Chrysanthemum by Modulating Gibberellin Biosynthesis. Plant Cell 2014, 26, 2038–2054. [Google Scholar] [CrossRef]
- Dally, N.; Xiao, K.; Holtgräwe, D.; Jung, C. The B2 flowering time locus of beet encodes a zinc finger transcription factor. Proc. Natl. Acad. Sci. USA 2014, 111, 10365–10370. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Tao, R.; Yin, L.; Ni, J.; Yang, Q.; Yan, X.; Yang, F.; Guo, X.; Li, H.; Teng, Y. Two B-box proteins, PpBBX18 and PpBBX21, antagonistically regulate anthocyanin biosynthesis via competitive association with Pyrus pyrifolia ELONGATED HYPOCOTYL 5 in the peel of pear fruit. Plant J. 2019, 100, 1208–1223. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhou, F.; Huang, X.-Q.; Chen, W.-C.; Kong, M.-J.; Zhou, C.-F.; Zhuang, Z.; Li, L.; Lu, S. ORANGE Represses Chloroplast Biogenesis in Etiolated Arabidopsis Cotyledons via Interaction with TCP14. Plant Cell 2019, 31, 2996–3014. [Google Scholar] [CrossRef]
- Job, N.; Datta, S. PIF3/HY5 module regulates BBX11 to suppress protochlorophyllide levels in dark and promote photomorphogenesis in light. New Phytol. 2021, 230, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Vaishak, K.P.; Yadukrishnan, P.; Bakshi, S.; Kushwaha, A.K.; Ramachandran, H.; Job, N.; Babu, D.; Datta, S. The B-box bridge between light and hormones in plants. J. Photochem. Photobiol. B Biol. 2019, 191, 164–174. [Google Scholar] [CrossRef]
- Bohlenius, H.; Huang, T.; Charbonnel-Campaa, L.; Brunner, A.M.; Jansson, S.; Strauss, S.H.; Nilsson, O. CO/FT Regulatory Module Controls Timing of Flowering and Seasonal Growth Cessation in Trees. Science 2006, 312, 1040–1043. [Google Scholar] [CrossRef]
- Cai, F.; Shao, C.; Zhang, Y.; Bao, Z.; Li, Z.; Shi, G.; Bao, M.; Zhang, J. Identification and characterisation of a novel FT orthologous gene in London plane with a distinct expression response to environmental stimuli compared to PaFT. Plant Biol. 2019, 21, 1039–1051. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, J.; Liu, G.; Li, X.; Lu, C.; Zhang, J.; Bao, M. Phylogenetic and evolutionary analysis of A-, B-, C- and E-class MADS-box genes in the basal eudicot Platanus acerifolia. J. Plant Res. 2012, 125, 381–393. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, G.; Guo, C.; He, Y.; Li, Z.; Ning, G.; Shi, X.; Bao, M. The FLOWERING LOCUS T orthologous gene of Platanus acerifolia is expressed as alternatively spliced forms with distinct spatial and temporal patterns. Plant Biol. 2011, 13, 809–820. [Google Scholar] [CrossRef]
- Cai, F.; Jin, X.; Tian, Y.; Huang, Z.; Wang, X.; Zhang, Y.; Sun, Y.; Shao, C. Molecular regulation of bud dormancy in perennial plants. Plant Growth Regul. 2023, 1–11. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jiao, C.; Jin, Q.; Lin, Y.; Cai, Y. B-BOX genes: Genome-wide identification, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri Rehd.). BMC Plant Biol. 2017, 17, 156. [Google Scholar] [CrossRef] [PubMed]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Zalabák, D.; Ikeda, Y. First Come, First Served: Sui Generis Features of the First Intron. Plants 2020, 9, 911. [Google Scholar] [CrossRef] [PubMed]
- Samach, A.; Onouchi, H.; Gold, S.E.; Ditta, G.S.; Schwarz-Sommer, Z.; Yanofsky, M.F.; Coupland, G. Distinct roles of CONSTANS target genes in reproductive development of Arabidopsis. Science 2000, 288, 1613–1616. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, X.; Weng, X.; Wang, L.; Xie, W. The rice B-box zinc finger gene family: Genomic identification, characterization, expression profiling and diurnal analysis. PloS ONE 2012, 7, e48242. [Google Scholar] [CrossRef]
- Ouyang, Y.; Pan, X.; Wei, Y.; Wang, J.; Xu, X.; He, Y.; Zhang, X.; Li, Z.; Zhang, H. Genome-wide identification and characterization of the BBX gene family in pineapple reveals that candidate genes are involved in floral induction and flowering. Genomics 2022, 114, 110397. [Google Scholar] [CrossRef]
- Zhao, J.; Li, H.; Huang, J.; Shi, T.; Meng, Z.; Chen, Q.; Deng, J. Genome-wide analysis of BBX gene family in Tartary buckwheat (Fagopyrum tataricum). PeerJ 2021, 9, e11939. [Google Scholar] [CrossRef]
- Liu, X.; Li, R.; Dai, Y.; Chen, X.; Wang, X. Genome-wide identification and expression analysis of the B-box gene family in the Apple (Malus domestica Borkh.) genome. Mol. Genet. Genom. 2018, 293, 303–315. [Google Scholar] [CrossRef]
- Wen, S.; Zhang, Y.; Deng, Y.; Chen, G.; Yu, Y.; Wei, Q. Genomic identification and expression analysis of the BBX transcription factor gene family in Petunia hybrida. Mol. Biol. Rep. 2020, 47, 6027–6041. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Liu, Q.; Zhang, T.; Chong, X.; Yuan, H. Genome-Wide Identification and Expression Analysis of BBX Transcription Factors in Iris germanica L. Int. J. Mol. Sci. 2021, 22, 8793. [Google Scholar] [CrossRef]
- Graeff, M.; Straub, D.; Eguen, T.; Dolde, U.; Rodrigues, V.; Brandt, R.; Wenkel, S. MicroProtein-Mediated Recruitment of CONSTANS into a TOPLESS Trimeric Complex Represses Flowering in Arabidopsis. PLoS Genet. 2016, 12, e1005959. [Google Scholar] [CrossRef] [PubMed]
- Holm, M.; Hardtke, C.S.; Gaudet, R.; Deng, X.W. Identification of a structural motif that confers specific interaction with the WD40 repeat domain of Arabidopsis COP1. EMBO J. 2001, 20, 118–127. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.; Ji, M.; Wu, Y.; Zhang, S.; Zhu, Y.; Yao, J.; Li, Z.; Gao, H.; Wang, X. Genome-wide identification and expression analysis of the B-box transcription factor gene family in grapevine (Vitis vinifera L.). BMC Genom. 2021, 22, 221. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.M.; Cui, L.; Wall, P.K.; Zhang, Q.; Zhang, X.; Leebens-Mack, J.; Ma, H.; Altman, N.; DePamphilis, C.W. Expression pattern shifts following duplication indicative of subfunctionalization and neofunctionalization in regulatory genes of Arabidopsis. Mol. Biol. Evol. 2006, 23, 469–478. [Google Scholar] [CrossRef]
- Bai, M.; Sun, J.; Liu, J.; Ren, H.; Wang, K.; Wang, Y.; Wang, C.; Dehesh, K. The B-box protein BBX19 suppresses seed germination via induction of ABI5. Plant J. 2019, 99, 1192–1202. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Adams, J.P.; No, K.; Liang, H.; Meilan, R.; Pechánová, O.; Barakat, A.; Carlson, J.E.; Page, G.P.; Yuceer, C. Overexpression of CONSTANS homologs CO1 and CO2 fails to alter normal reproductive onset and fall bud set in woody perennial poplar. PLoS ONE 2012, 7, e45448. [Google Scholar] [CrossRef]
- Gao, X.-R.; Zhang, H.; Li, X.; Bai, Y.-W.; Peng, K.; Wang, Z.; Dai, Z.-R.; Bian, X.-F.; Zhang, Q.; Jia, L.-C.; et al. The B-box transcription factor IbBBX29 regulates leaf development and flavonoid biosynthesis in sweet potato. Plant Physiol. 2023, 191, 496–514. [Google Scholar] [CrossRef]
- Yadav, A.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. BBX31 promotes hypocotyl growth, primary root elongation and UV-B tolerance in Arabidopsis. Plant Signal. Behav. 2019, 14, e1588672. [Google Scholar] [CrossRef]
- Conti, L. Hormonal control of the floral transition: Can one catch them all? Dev. Biol. 2017, 430, 288–301. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gao, Z.; Du, P.; Xiao, W.; Tan, Q.; Chen, X.; Li, L.; Gao, D. Expression of ABA Metabolism-Related Genes Suggests Similarities and Differences Between Seed Dormancy and Bud Dormancy of Peach (Prunus persica). Front. Plant Sci. 2016, 6, 1248. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Niu, Q.; Tang, Y.; Ma, Y.; Yan, X.; Li, J.; Tian, J.; Bai, S.; Teng, Y. PpyGAST1 is potentially involved in bud dormancy release by integrating the GA biosynthesis and ABA signaling in ‘Suli’ pear (Pyrus pyrifolia White Pear Group). Environ. Exp. Bot. 2019, 162, 302–312. [Google Scholar] [CrossRef]
- Hao, X.; Tang, H.; Wang, B.; Wang, L.; Cao, H.; Wang, Y.; Zeng, J.; Fang, S.; Chu, J.; Yang, Y.; et al. Gene Characterization and Expression Analysis Reveal the Importance of Auxin Signaling in Bud Dormancy Regulation in Tea Plant. J. Plant Growth Regul. 2018, 38, 225–240. [Google Scholar] [CrossRef]
- Xu, D.; Li, J.; Gangappa, S.N.; Hettiarachchi, C.; Lin, F.; Andersson, M.X.; Jiang, Y.; Deng, X.W.; Holm, M. Convergence of Light and ABA signaling on the ABI5 promoter. PLoS Genet. 2014, 10, e1004197. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, X.; Aiwaili, P.; Mu, X.; Zhao, M.; Zhao, J.; Cheng, L.; Ma, C.; Gao, J.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Zhu, L.; Guan, Y.; Liu, Y.; Zhang, Z.; Jaffar, M.A.; Song, A.; Chen, S.; Jiang, J.; Chen, F. Regulation of flowering time in chrysanthemum by the R2R3 MYB transcription factor CmMYB2 is associated with changes in gibberellin metabolism. Hortic. Res. 2020, 7, 96. [Google Scholar] [CrossRef]
- Lee, J.; He, K.; Stolc, V.; Lee, H.; Figueroa, P.; Gao, Y.; Tongprasit, W.; Zhao, H.; Lee, I.; Deng, X.W. Analysis of transcription factor HY5 genomic binding sites revealed its hierarchical role in light regulation of development. Plant Cell 2007, 19, 731–749. [Google Scholar] [CrossRef]
- Maurya, J.P.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of growth cessation and dormancy in trees: A molecular perspective. Ann. Bot. 2017, 120, 351–360. [Google Scholar] [CrossRef]
- Talar, U.; Kielbowicz-Matuk, A. Beyond Arabidopsis: BBX Regulators in Crop Plants. Int. J. Mol. Sci. 2021, 22, 2906. [Google Scholar] [CrossRef]
- Susila, H.; Nasim, Z.; Gawarecka, K.; Jung, J.-Y.; Jin, S.; Youn, G.; Ahn, J.H. Chloroplasts prevent precocious flowering through a GOLDEN2-LIKE-B-BOX DOMAIN PROTEIN module. Plant Commun. 2023, 2, 100515. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xu, J.; Yuan, C.; Chen, Q.; Liu, Q.; Wang, X.; Qin, C. BBX17 Interacts with CO and Negatively Regulates Flowering Time in Arabidopsis thaliana. Plant Cell Physiol. 2022, 63, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, Y.S.; Ma, C.; Strauss, S.H.; Busov, V.B. EARLY BUD-BREAK 1 (EBB1) is a regulator of release from seasonal dormancy in poplar trees. Proc. Natl. Acad. Sci. USA 2014, 111, 10001–10006. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; He, Y.; Xia, R. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, G.; Zhang, J.; Zhang, J.; Bao, M. Extraction of high-quality tissue-specific RNA from Londonplane trees (Platanus acerifolia), permitting the construction of a female inflorescence cDNA library. Funct. Plant Biol. 2008, 35, 159–165. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Identifier | Chromosomes Position | AA | MW (kDa) | pI |
|---|---|---|---|---|---|
| PaBBX1-1 | PaC3G126980 | 53,321,272–53,324,171 | 383 | 42.81 | 6.04 |
| PaBBX1-2 | PaB3G200600 | 57,602,006–57,605,538 | 377 | 41.98 | 5.68 |
| PaBBX1-3 | PaA3G246120 | 51,710,999–51,713,790 | 383 | 42.22 | 5.93 |
| PaBBX4 | PaA6G276070 | 27,421,637–27,423,559 | 374 | 41.07 | 5.33 |
| PaBBX5-1 | PaB6G308930 | 27,080,769–27,082,589 | 376 | 41.39 | 5.38 |
| PaBBX5-2 | PaC6G408130 | 17,664,745–17,666,673 | 377 | 41.29 | 5.56 |
| PaBBX7 | PaC2G374890 | 68,135,516–68,144,895 | 436 | 47.57 | 5.25 |
| PaBBX8-1 | PaA3G242790 | 61,010,653–61,017,031 | 405 | 44.36 | 5.59 |
| PaBBX8-2 | PaB2G072670 | 91,892,447–91,898,521 | 405 | 44.44 | 5.01 |
| PaBBX9-1 | PaA2G259070 | 66,033,109–66,043,294 | 419 | 45.96 | 5.69 |
| PaBBX9-2 | PaC2G370070 | 56,914,445–56,942,510 | 439 | 48.10 | 5.47 |
| PaBBX10 | PaC3G123980 | 36,237,251–36,289,053 | 520 | 58.57 | 5.88 |
| PaBBX11-1 | PaA5G347500 | 12,207,675–12,212,906 | 400 | 45.51 | 5.93 |
| PaBBX11-2 | PaC5G216800 | 12,060,240–12,064,960 | 409 | 45.22 | 5.16 |
| PaBBX11-3a | PaB1G042960 | 71,884,034–71,889,372 | 311 | 35.02 | 5.04 |
| PaBBX11-3b | PaUnG429150-RA | 43,527–48,865 | 311 | 35.02 | 5.04 |
| PaBBX12 | PaC3G139600 | 87,665,264–87,668,302 | 485 | 53.53 | 6.12 |
| PaBBX13 | PaA3G234590 | 79,284,256–79,287,445 | 492 | 54.23 | 6.8 |
| PaBBX15-1 | PaB3G199040 | 51,735,509–51,737,376 | 432 | 48.20 | 4.97 |
| PaBBX15-2 | PaA3G247750 | 46,315,091–46,315,360 | 437 | 48.34 | 4.95 |
| PaBBX15-3 | PaC3G125090 | 42,957,754–42,959,478 | 417 | 46.43 | 5.11 |
| PaBBX19-1 | PaB4G179910 | 64,824,309–64,835,187 | 204 | 23.14 | 5.34 |
| PaBBX19-2 | PaA4G084960 | 64,411,679–64,420,184 | 223 | 24.95 | 6.19 |
| PaBBX19-3 | NA | 58,602,036–58,606,617 | 197 | 21.99 | 6.38 |
| PaBBX19-4 | PaUnG529740-RA | 2505–3689 | 179 | 20.12 | 6.22 |
| PaBBX21-1 | PaB4G180130 | 65,601,829–65,602,921 | 297 | 32.81 | 6.36 |
| PaBBX21-2 | PaA4G084640 | 65,243,318–65,245,509 | 284 | 31.62 | 6.2 |
| PaBBX21-3 | PaC4G152800 | 59,354,451–59,356,546 | 188 | 20.76 | 6.28 |
| PaBBX22-1 | PaB2G060740 | 48,920,313–48,948,818 | 299 | 32.77 | 5.33 |
| PaBBX22-2 | PaC2G363350 | 33,447,397–33,470,603 | 297 | 32.56 | 5.84 |
| PaBBX23 | PaB4G165570 | 3,297,234–3,310,372 | 206 | 22.34 | 6.43 |
| PaBBX24 | PaC5G232000 | 76,674,127–76,682,184 | 244 | 26.85 | 4.9 |
| PaBBX25 | PaB5G340870 | 61,197,235–61,199,987 | 238 | 26.37 | 4.87 |
| PaBBX28/29-1 | PaB2G067580 | 76,340,288–76,341,894 | 322 | 35.13 | 4.61 |
| PaBBX28/29-2 | PaA2G260070 | 61,929,831–61,932,394 | 206 | 22.36 | 8.63 |
| PaBBX28/29-3 | PaC2G369370 | 55,193,130–55,195,276 | 278 | 29.96 | 4.44 |
| PaBBX30 | NA | 56,910,595–56,910,981 | 130 | 14.90 | 8.06 |
| PaBBX32-1 | PaB6G307570 | 13,158,377–13,159,201 | 258 | 28.71 | 9.35 |
| PaBBX32-2 | PaA6G274390 | 14,746,991–14,747,818 | 264 | 29.09 | 8.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, G.; Ai, K.; Yan, X.; Zhou, Z.; Cai, F.; Bao, M.; Zhang, J. Genome-Wide Analysis of the BBX Genes in Platanus × acerifolia and Their Relationship with Flowering and/or Dormancy. Int. J. Mol. Sci. 2023, 24, 8576. https://doi.org/10.3390/ijms24108576
Shi G, Ai K, Yan X, Zhou Z, Cai F, Bao M, Zhang J. Genome-Wide Analysis of the BBX Genes in Platanus × acerifolia and Their Relationship with Flowering and/or Dormancy. International Journal of Molecular Sciences. 2023; 24(10):8576. https://doi.org/10.3390/ijms24108576
Chicago/Turabian StyleShi, Gehui, Kangyu Ai, Xu Yan, Zheng Zhou, Fangfang Cai, Manzhu Bao, and Jiaqi Zhang. 2023. "Genome-Wide Analysis of the BBX Genes in Platanus × acerifolia and Their Relationship with Flowering and/or Dormancy" International Journal of Molecular Sciences 24, no. 10: 8576. https://doi.org/10.3390/ijms24108576