
Dihydroceramides Derived from Bacteroidetes Species Sensitize TRPV1 Channels
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
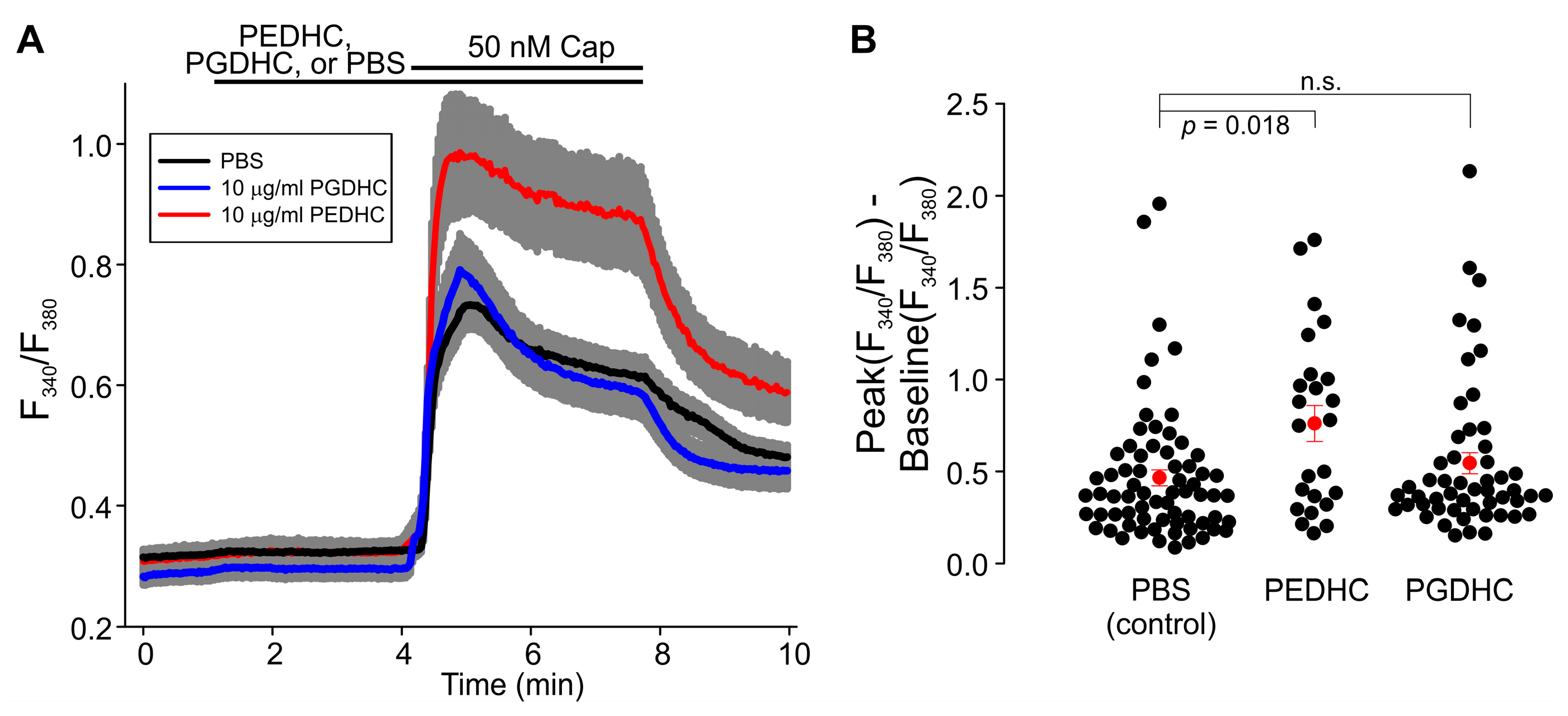
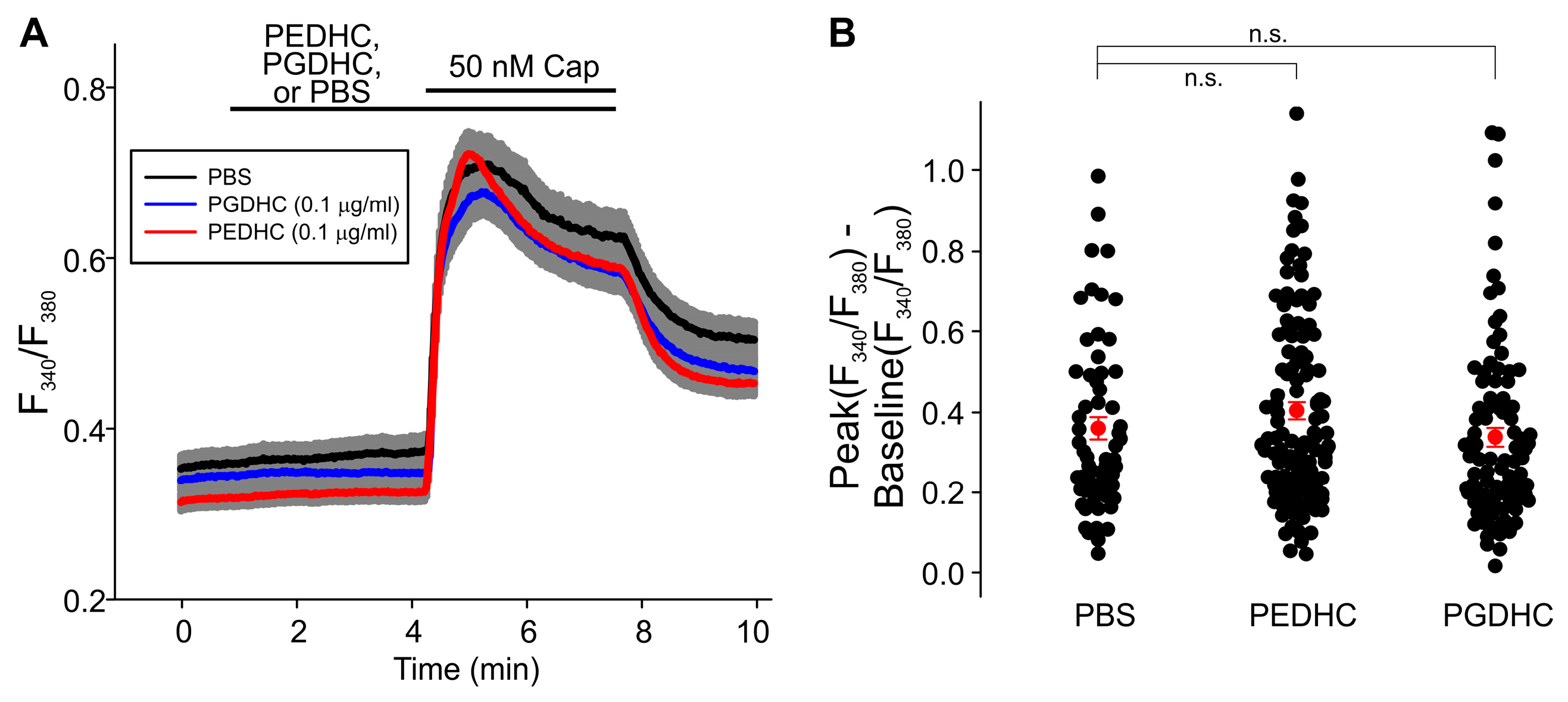
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Fluorescence Imaging
4.3. Electrophysiological Recordings
4.4. Materials
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munjuluri, S.; Wilkerson, D.A.; Sooch, G.; Chen, X.; White, F.A.; Obukhov, A.G. Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation. Cells 2021, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, J.L.; Melnyk, J.L.; Basbaum, A.I. Differential TRPV1 and TRPV2 channel expression in dental pulp. J. Dent. Res. 2011, 90, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.; Yamada, C.; Garcia, C.; Akkaoui, J.; Ho, A.; Nichols, F.; Movila, A. Crosstalk between dihydroceramides produced by Porphyromonas gingivalis and host lysosomal cathepsin B in the promotion of osteoclastogenesis. J. Cell. Mol. Med. 2022, 26, 2841–2851. [Google Scholar] [CrossRef] [PubMed]
- Cutting, K.F.; White, R.J.; Mahoney, P. Wound infection, dressings and pain, is there a relationship in the chronic wound? Int. Wound J. 2013, 10, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound microbiology and associated approaches to wound management. Clin. Microbiol. Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef] [Green Version]
- Salyers, A.A.; Shoemaker, N.B.; Guthrie, E.P. Recent advances in Bacteroides genetics. Crit. Rev. Microbiol. 1987, 14, 49–71. [Google Scholar] [CrossRef]
- Cao, H.; Liu, M.C.; Tong, M.K.; Jiang, S.; Chow, K.H.; To, K.K.; Tse, C.W.; Ho, P.L. Comprehensive investigation of antibiotic resistance gene content in cfiA-harboring Bacteroides fragilis isolates of human and animal origins by whole genome sequencing. Int. J. Med. Microbiol. 2022, 312, 151559. [Google Scholar] [CrossRef]
- Leigh, D.A. Wound infections due to Bacteroides fragilis following intestinal surgery. Br. J. Surg. 1975, 62, 375–378. [Google Scholar] [CrossRef]
- Alexiou, K.; Drikos, I.; Terzopoulou, M.; Sikalias, N.; Ioannidis, A.; Economou, N. A prospective randomised trial of isolated pathogens of surgical site infections (SSI). Ann. Med. Surg. (Lond.) 2017, 21, 25–29. [Google Scholar] [CrossRef]
- Shenoy, P.A.; Vishwanath, S.; Nagaraj, R.T.; Banerjee, B.; Krishna, M.S. Diabetic Foot Infection with Bacteroides pyogenes. J. Glob. Infect Dis. 2021, 13, 186–188. [Google Scholar] [CrossRef]
- Hsieh, C.T.; Lee, Y.J.; Dai, X.; Ojeda, N.B.; Lee, H.J.; Tien, L.T.; Fan, L.W. Systemic Lipopolysaccharide-Induced Pain Sensitivity and Spinal Inflammation Were Reduced by Minocycline in Neonatal Rats. Int. J. Mol. Sci. 2018, 19, 2947. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Song, S.; Zhang, Y.; Ge, Y.; Fang, X.; Huang, T.; Du, J.; Gao, J. Inhibition of the Rho/Rho kinase pathway prevents lipopolysaccharide-induced hyperalgesia and the release of TNF-alpha and IL-1beta in the mouse spinal cord. Sci. Rep. 2015, 5, 14553. [Google Scholar] [CrossRef]
- Diogenes, A.; Ferraz, C.C.; Akopian, A.N.; Henry, M.A.; Hargreaves, K.M. LPS sensitizes TRPV1 via activation of TLR4 in trigeminal sensory neurons. J. Dent. Res. 2011, 90, 759–764. [Google Scholar] [CrossRef]
- Nichols, F.C.; Yao, X.; Bajrami, B.; Downes, J.; Finegold, S.M.; Knee, E.; Gallagher, J.J.; Housley, W.J.; Clark, R.B. Phosphorylated dihydroceramides from common human bacteria are recovered in human tissues. PLoS ONE 2011, 6, e16771. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.L.; Heaver, S.L.; Waters, J.L.; Kim, B.I.; Bretin, A.; Goodman, A.L.; Gewirtz, A.T.; Worgall, T.S.; Ley, R.E. Sphingolipids produced by gut bacteria enter host metabolic pathways impacting ceramide levels. Nat. Commun. 2020, 11, 2471. [Google Scholar] [CrossRef]
- Nichols, F.C. Novel ceramides recovered from Porphyromonas gingivalis: Relationship to adult periodontitis. J. Lipid Res. 1998, 39, 2360–2372. [Google Scholar] [CrossRef]
- Kanzaki, H.; Movila, A.; Kayal, R.; Napimoga, M.H.; Egashira, K.; Dewhirst, F.; Sasaki, H.; Howait, M.; Al-Dharrab, A.; Mira, A.; et al. Phosphoglycerol dihydroceramide, a distinctive ceramide produced by Porphyromonas gingivalis, promotes RANKL-induced osteoclastogenesis by acting on non-muscle myosin II-A (Myh9), an osteoclast cell fusion regulatory factor. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Nativel, B.; Couret, D.; Giraud, P.; Meilhac, O.; d'Hellencourt, C.L.; Viranaicken, W.; Da Silva, C.R. Porphyromonas gingivalis lipopolysaccharides act exclusively through TLR4 with a resilience between mouse and human. Sci. Rep. 2017, 7, 15789. [Google Scholar] [CrossRef] [Green Version]
- Holland, W.L.; Bikman, B.T.; Wang, L.P.; Yuguang, G.; Sargent, K.M.; Bulchand, S.; Knotts, T.A.; Shui, G.; Clegg, D.J.; Wenk, M.R.; et al. Lipid-induced insulin resistance mediated by the proinflammatory receptor TLR4 requires saturated fatty acid-induced ceramide biosynthesis in mice. J. Clin. Investig. 2011, 121, 1858–1870. [Google Scholar] [CrossRef] [Green Version]
- Darveau, R.P.; Pham, T.T.; Lemley, K.; Reife, R.A.; Bainbridge, B.W.; Coats, S.R.; Howald, W.N.; Way, S.S.; Hajjar, A.M. Porphyromonas gingivalis lipopolysaccharide contains multiple lipid A species that functionally interact with both toll-like receptors 2 and 4. Infect. Immun. 2004, 72, 5041–5051. [Google Scholar] [CrossRef]
- Medvedev, A.E.; Vogel, S.N. Overexpression of CD14, TLR4, and MD-2 in HEK 293T cells does not prevent induction of in vitro endotoxin tolerance. J. Endotoxin Res. 2003, 9, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Young, D.W.; Gusovsky, F.; Chow, J.C. Cellular events mediated by lipopolysaccharide-stimulated toll-like receptor 4. MD-2 is required for activation of mitogen-activated protein kinases and Elk-1. J. Biol. Chem. 2000, 275, 20861–20866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saghy, E.; Szoke, E.; Payrits, M.; Helyes, Z.; Borzsei, R.; Erostyak, J.; Janosi, T.Z.; Setalo, G., Jr.; Szolcsanyi, J. Evidence for the role of lipid rafts and sphingomyelin in Ca2+-gating of Transient Receptor Potential channels in trigeminal sensory neurons and peripheral nerve terminals. Pharmacol. Res. 2015, 100, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Cao, E.; Julius, D.; Cheng, Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action. Nature 2016, 534, 347–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ufret-Vincenty, C.A.; Klein, R.M.; Hua, L.; Angueyra, J.; Gordon, S.E. Localization of the PIP2 sensor of TRPV1 ion channels. J. Biol. Chem. 2011, 286, 9688–9698. [Google Scholar] [CrossRef] [Green Version]
- Prescott, E.D.; Julius, D. A modular PIP2 binding site as a determinant of capsaicin receptor sensitivity. Science 2003, 300, 1284–1288. [Google Scholar] [CrossRef]
- Kim, A.Y.; Tang, Z.; Liu, Q.; Patel, K.N.; Maag, D.; Geng, Y.; Dong, X. Pirt, a phosphoinositide-binding protein, functions as a regulatory subunit of TRPV1. Cell 2008, 133, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Sattler, K.; El-Battrawy, I.; Cyganek, L.; Lang, S.; Lan, H.; Li, X.; Zhao, Z.; Utikal, J.; Wieland, T.; Borggrefe, M.; et al. TRPV1 activation and internalization is part of the LPS-induced inflammation in human iPSC-derived cardiomyocytes. Sci. Rep. 2021, 11, 14689. [Google Scholar] [CrossRef]
- Sen, C.K. Human Wounds and Its Burden: An Updated Compendium of Estimates. Adv. Wound Care (New Rochelle) 2019, 8, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Oboukhova, E.A.; Kumar, S.; Sturek, M.; Obukhov, A.G. Canonical transient receptor potential channels expression is elevated in a porcine model of metabolic syndrome. Mol. Endocrinol. 2009, 23, 689–699. [Google Scholar] [CrossRef]
- Chen, X.; Sun, W.; Gianaris, N.G.; Riley, A.M.; Cummins, T.R.; Fehrenbacher, J.C.; Obukhov, A.G. Furanocoumarins are a novel class of modulators for the transient receptor potential vanilloid type 1 (TRPV1) channel. J. Biol. Chem. 2014, 289, 9600–9610. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, W.; Riley, A.M.; Soliman, M.; Chakraborty, S.; Stamatkin, C.W.; Obukhov, A.G. Molecular determinants of the sensitivity to Gq/11-phospholipase C-dependent gating, Gd3+ potentiation, and Ca2+ permeability in the transient receptor potential canonical type 5 (TRPC5) channel. J. Biol. Chem. 2017, 292, 898–911. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Duan, Y.; Riley, A.M.; Welch, M.A.; White, F.A.; Grant, M.B.; Obukhov, A.G. Long-term diabetic microenvironment augments the decay rate of capsaicin-induced currents in mouse dorsal root ganglion neurons. Molecules 2019, 24, 775. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ludwig, N.; Demaree, I.S.; Yamada, C.; Nusbaum, A.; Nichols, F.C.; White, F.A.; Movila, A.; Obukhov, A.G. Dihydroceramides Derived from Bacteroidetes Species Sensitize TRPV1 Channels. Int. J. Mol. Sci. 2023, 24, 877. https://doi.org/10.3390/ijms24010877
Ludwig N, Demaree IS, Yamada C, Nusbaum A, Nichols FC, White FA, Movila A, Obukhov AG. Dihydroceramides Derived from Bacteroidetes Species Sensitize TRPV1 Channels. International Journal of Molecular Sciences. 2023; 24(1):877. https://doi.org/10.3390/ijms24010877
Chicago/Turabian StyleLudwig, Nora, Isaac S. Demaree, Chiaki Yamada, Amilia Nusbaum, Frank C. Nichols, Fletcher A. White, Alexandru Movila, and Alexander G. Obukhov. 2023. "Dihydroceramides Derived from Bacteroidetes Species Sensitize TRPV1 Channels" International Journal of Molecular Sciences 24, no. 1: 877. https://doi.org/10.3390/ijms24010877