
Isolation of a Novel QTL, qSCM4, Associated with Strong Culm Affects Lodging Resistance and Panicle Branch Number in Rice
 , ,
, ,
Abstract
:1. Introduction
2. Results
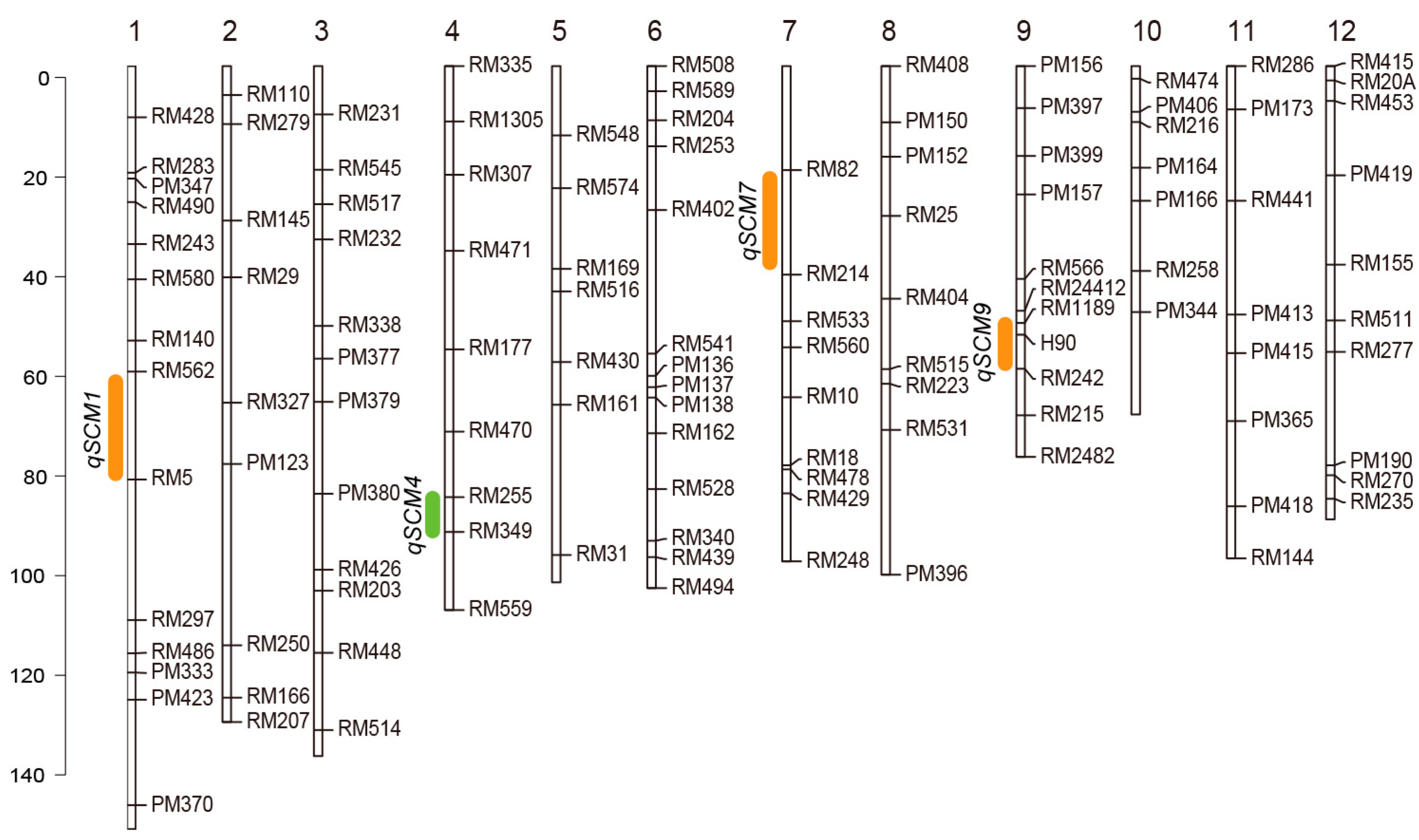
2.1. QTL Mapping of Yield and Lodging Related Traits on Chromosome 4 in Different Environments
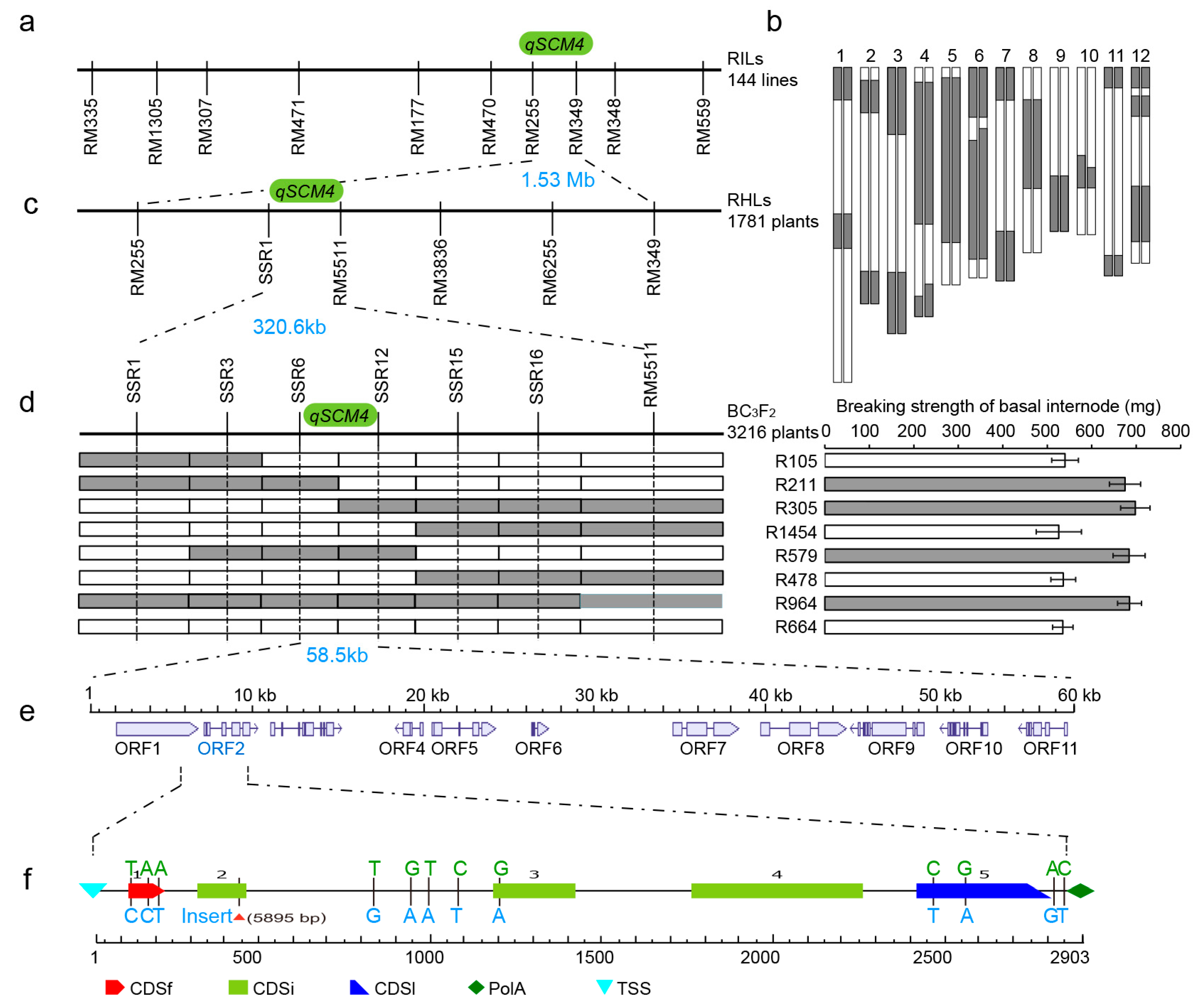
2.2. Fine Mapping and Candidate Gene Analysis of the Strong Culm and Lodging Resistance QTL qSCM4
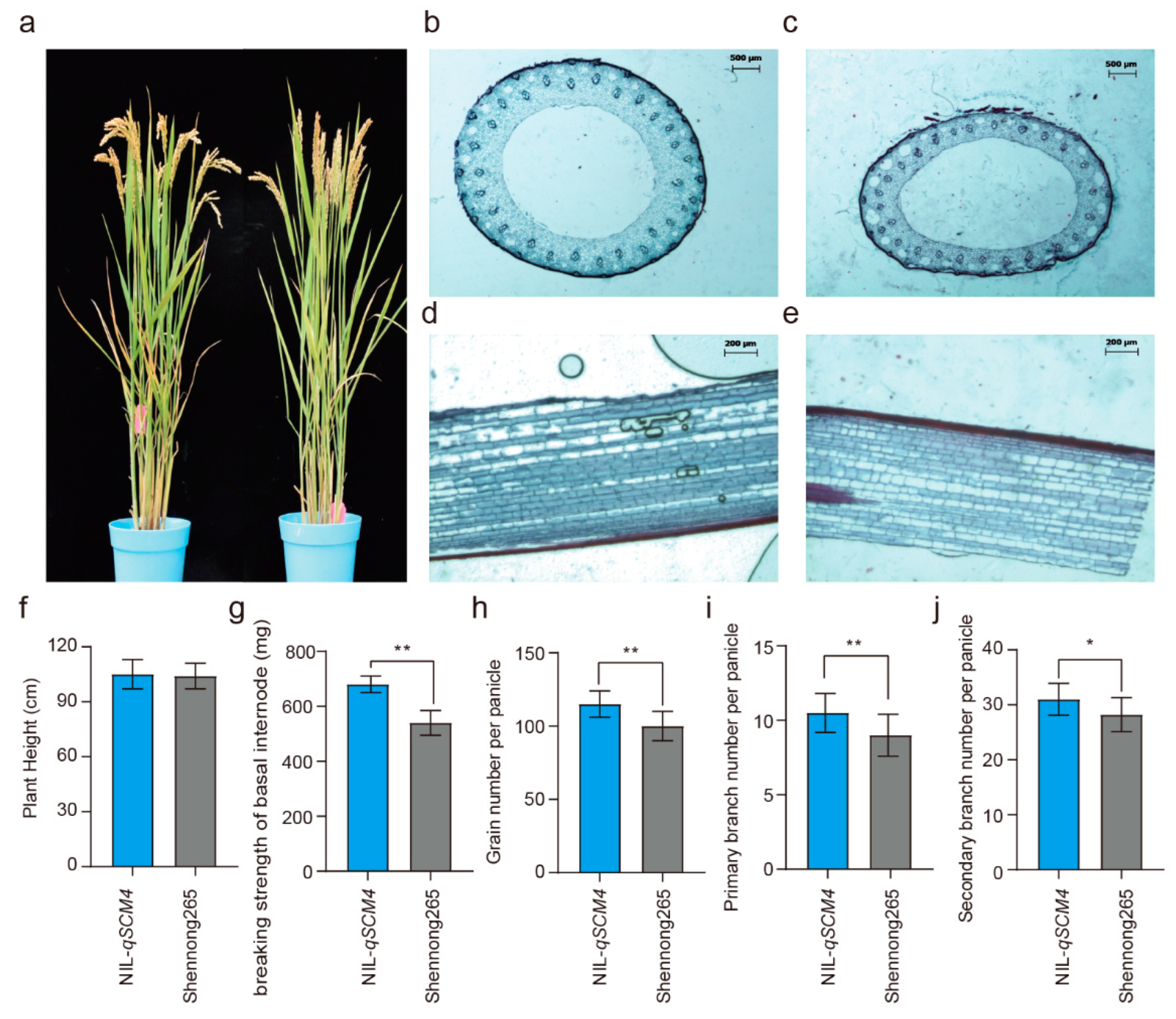
2.3. Effect of qSCM4 on the Traits Related to Lodging and Panicle and Anatomical Structure Analysis
3. Discussion
4. Materials and Methods
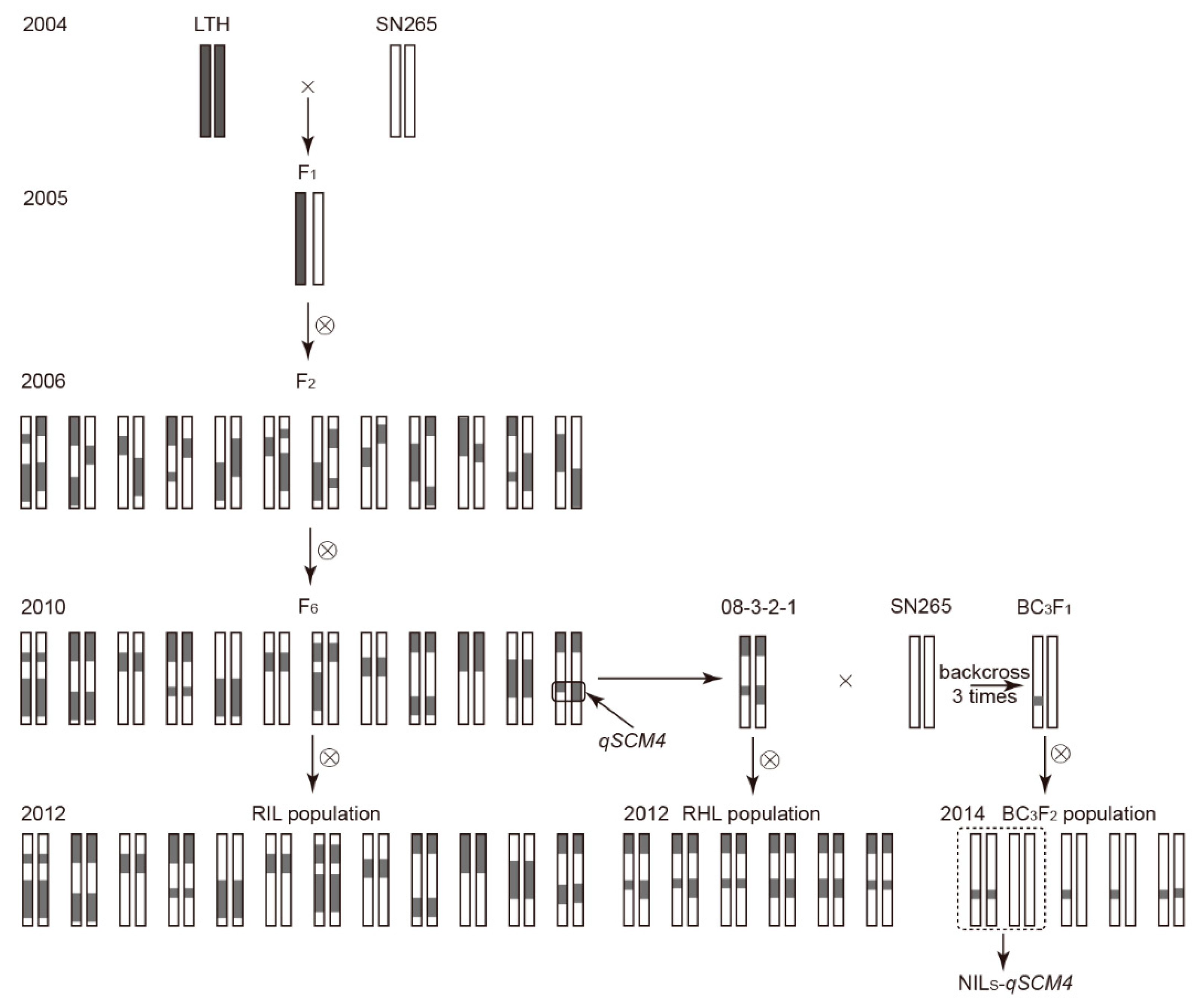
4.1. Mapping Population and Its Linkage Map
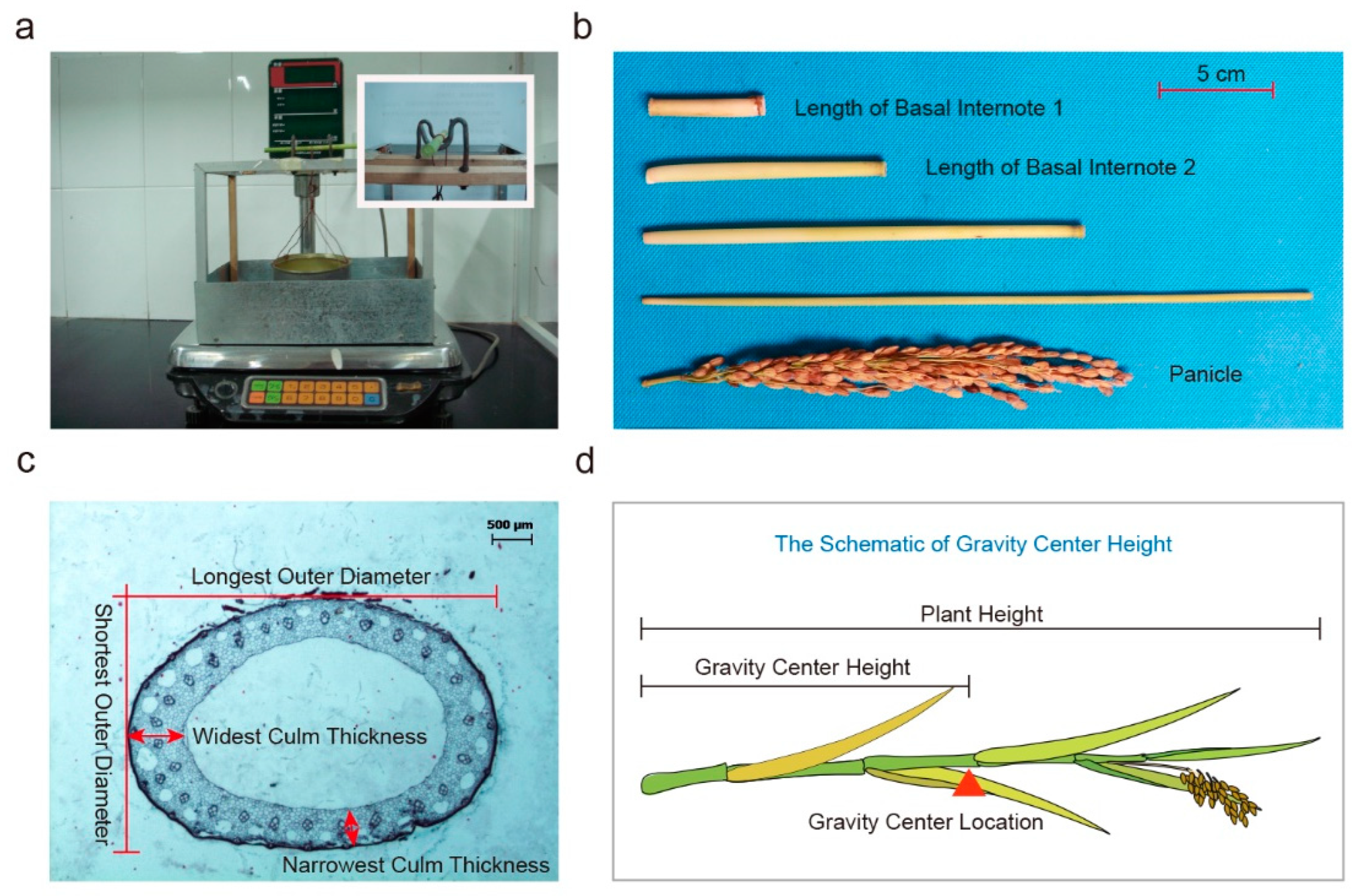
4.2. Cultivation, Evaluation of Lodging and Yield Related Traits
4.3. QTL Analysis, Fine Mapping of qSCM4 and Candidate Gene Analysis
4.4. DNA Sequence of Os04g0615000
4.5. Determination of Stalk Anatomical Structure
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rajkumara, S. Lodging in Cereals—A Review. Agric. Rev. 2008, 29, 55–60. [Google Scholar]
- Setter, T.L.; Laureles, E.V.; Mazaredo, A.M. Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis. Field Crops Res. 1997, 49, 95–106. [Google Scholar] [CrossRef]
- Nakajima, T.; Yoshida, M.; Tomimura, K. Effect of lodging on the level of mycotoxins in wheat, barley, and rice infected with the Fusarium graminearum species complex. J. Gen. Plant Pathol. 2008, 74, 289. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Hirotsu, N.; Madoka, Y.; Ookawa, T.; Ishimaru, K. Improvement of Resistance to Bending-Type Lodging in Rice. Jpn. J. Crop Sci. 2007, 76, 1–9. [Google Scholar] [CrossRef]
- Ishimaru, K.; Togawa, E.; Ookawa, T.; Kashiwaga, T.; Madoka, Y.; Hirotsu, N. New target for rice lodging resistance and its effect in a typhoon. Planta 2008, 227, 601–609. [Google Scholar] [CrossRef]
- Cui, Y.T.; Hu, X.M.; Liang, G.H.; Feng, A.H.; Wang, F.M.; Ruan, S.; Dong, G.J.; Shen, L.; Zhang, B.; Chen, D.D.; et al. Production of novel beneficial alleles of a rice yield-related QTL by CRISPR/Cas9. Plant Biotechnol. J. 2020, 18, 1987–1989. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Sasaki, H.; Ishimaru, K. Factors responsible for decreasing sturdiness of the lower part in lodging of rice (Oryza sativa L.). Plant Prod. Sci. 2005, 8, 166–172. [Google Scholar] [CrossRef]
- Kg, C. Ecological intensification of cereal production systems: Yield potential, soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 1999, 96, 5952–9595. [Google Scholar]
- Ma, G.H.; Yuan, L.P. Hybrid rice achievements, development and prospect in China. J. Integr. Agric. 2015, 14, 197–205. [Google Scholar] [CrossRef]
- Nomura, T.; Arakawa, N.; Yamamoto, T.; Ueda, T.; Adachi, S.; Yonemaru, J.; Abe, A.; Takagi, H.; Yokoyama, T.; Ookawa, T. Next generation long-culm rice with superior lodging resistance and high grain yield, Monster Rice 1. PLoS ONE 2019, 14, e0221424. [Google Scholar] [CrossRef]
- Liu, S.T.; Huang, Y.W.; Xu, H.; Zhao, M.Z.; Xu, Q.; Li, F.C. Genetic enhancement of lodging resistance in rice due to the key cell wall polymer lignin, which affects stem characteristics. Breed. Sci. 2018, 68, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.J.; Wu, L.M.; Wu, X.R.; Ding, Y.F.; Li, G.H.; Li, J.R.; Weng, F.; Liu, Z.H.; Tang, S.; Ding, C.Q.; et al. Lodging Resistance of Japonica Rice (Oryza Sativa L.): Morphological and Anatomical Traits due to top-Dressing Nitrogen Application Rates. Rice 2016, 9, 31. [Google Scholar] [CrossRef]
- Duan, C.N.; Wang, B.C.; Wang, P.Q.; Wang, D.H.; Cai, S.X. Relationship between the minute structure and thelodging resistance of rice stems. Colloids Surf. B Biointerfaces 2004, 35, 155–158. [Google Scholar]
- Hirano, K.; Ordonio, R.; Matsuoka, M. Engineering the lodging resistance mechanism of post-Green Revolution rice to meet future demands. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 220–233. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, T.; Togawa, E.; Hirotsu, N.; Ishimaru, K. Improvement of lodging resistance with QTLs for stem diameter in rice (Oryza sativa L.). Thero. Appl. Genet. 2008, 117, 749–757. [Google Scholar] [CrossRef]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, S.; Gui, J.S.; Fu, C.J.; Yu, H.; Song, D.L.; Shen, J.H.; Qin, P.; Liu, X.M.; Han, B.; et al. Shortened Basal Internodes Encodes a Gibberellin 2-Oxidase and Contributes to Lodging Resistance in Rice. Mol. Plant 2018, 11, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Sunohara, H.; Miura, K.; Wu, X.; Saeda, T.; Mizuno, S.; Ashikari, M.; Matsuoka, M.; Kitano, H. Effects of Ssi1 Gene Controlling dm-type Internode Elongation Pattern on Lodging Resistance and Panicle Characters in Rice. Breed. Sci. 2006, 56, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.C.; Zhou, Y.H. Rice Brittleness Mutants: A Way to Open the ‘Black Box’ of Monocot Cell Wall Biosynthesis. J. Integr. Plant Biol. 2011, 53, 136–142. [Google Scholar] [CrossRef]
- Li, F.C.; Liu, S.T.; Xu, H.; Xu, Q. A novel FC17/CESA4 mutation causes increased biomass saccharifcation and lodging resistance by remodeling cell wall in rice. Biotechnol. Biofuels 2018, 11, 298. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.F.; Qin, Y.L.; Fang, J.J.; Yuan, S.J.; Peng, L.X.; Zhao, J.F.; Li, X.Y. A Missense Mutation in the Zinc Finger Domain of OsCESA7 Deleteriously Affects Cellulose Biosynthesis and Plant Growth in Rice. PLoS ONE 2016, 11, e0153993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.C.; Xie, G.S.; Huang, J.F.; Zhang, R.; Li, Y.; Zhang, M.M.; Wang, Y.T.; Li, A.; Li, X.K.; Xia, T.; et al. OsCESA9 conserved-site mutation leads to largely enhanced plant lodging resistance and biomass enzymatic saccharification by reducing cellulose DP and crystallinity in rice. Plant Biotechnol. J. 2017, 15, 1093–1104. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zeng, X.; Li, Y.; Li, J.; Huang, X.; Zhao, D. BRITTLE CULM17, a Novel Allele of TAC4, Affects the Mechanical Properties of Rice Plants. Int. J. Mol. Sci. 2022, 23, 5305. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Kotake, T.; Kamihara, K.; Tsuna, K.; Aobhara, T.; Kaneko, Y.; Takatsuji, H.; Tsumuraya, Y.; Kawasaki, S. Rice BRITTLE CULM 3 (BC3) encodes a classical dynamin OsDRP2B essential for proper secondary cell wall synthesis. Planta 2010, 232, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.Y.; Li, R.; Qian, Q.; Song, X.Q.; Liu, X.L.; Yu, Y.C.; Zeng, D.L.; Wan, J.M.; Li, J.Y.; Zhou, Y.H. The rice dynamin-related protein DRP2B mediates membrane trafficking, and thereby plays a critical role in secondary cell wall cellulose biosynthesis. Plant J. 2010, 64, 56–70. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Li, S.B.; Qian, Q.; Zeng, D.L.; Zhang, M.; Guo, L.B.; Liu, X.L.; Zhang, B.C.; Deng, L.W.; Liu, X.F.; et al. BC10, a DUF266-containing and Golgi-located type II membrane protein, is required for cell-wall biosynthesis in rice (Oryza sativa L.). Plant J. 2009, 57, 446–462. [Google Scholar] [CrossRef]
- Wu, B.; Zhang, B.C.; Dai, Y.; Zhang, L.; Shang-Guan, K.K.; Peng, Y.G.; Zhou, Y.H.; Zhu, Z. Brittle Culm15 encodes a membrane-associated chitinase-like protein required for cellulose biosynthesis in rice. Plant Physiol. 2012, 159, 1440–1452. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.F.; Li, Y.; Hu, Z.; Hu, H.Z.; Wang, G.Y.; Li, A.; Wang, Y.M.; Tu, Y.Y.; Xia, T.; Peng, L.C.; et al. Ectopic expression of a novel OsExtensin-like gene consistently enhances plant lodging resistance by regulating cell elongation and cell wall thickening in rice. Plant Biotechnol. J. 2018, 16, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, T.; Munakata, J.; Ishimaru, K. Functional analysis of the lodging resistance QTL BSUC11 on morphological and chemical characteristics in upper culms of rice. Euphytica 2016, 210, 233–243. [Google Scholar] [CrossRef]
- Jiao, Y.Q.; Wang, Y.H.; Xue, D.W.; Wang, J.; Yan, W.X.; Liu, G.F.; Dong, G.J.; Zeng, D.L.; Lu, Z.F.; Zhu, X.D.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Ookawa, T.; Inoue, K.; Matsuoka, M.; Ebitani, T.; Takarada, T.; Yamamoto, T.; Ueda, T.; Yokoyama, T.; Sugiyama, C.; Nakaba, S.; et al. Increased lodging resistance in long-culm, low-lignin gh2 rice for improved feed and bioenergy production. Sci. Rep. 2014, 4, 6567. [Google Scholar] [CrossRef] [Green Version]
- Yano, K.; Ookawa, T.; Aya, K.; Ochiai, Y.; Hirasawa, T.; Ebitani, T.; Takarada, T.; Yano, M.; Yamamoto, T.; Fukuoka, S.; et al. Isolation of a Novel Lodging Resistance QTL Gene Involved in Strigolactone Signaling and Its Pyramiding with a QTL Gene Involved in Another Mechanism. Mol. Plant 2015, 8, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Ookawa, T.; Nomura, T.; Kamahora, E.; Jiang, M.; Ochiai, Y.; Samadi, A.; Yamaguchi, T.; Adachi, S.; Katsura, K.; Motobayashi, T. Pyramiding of multiple strong-culm genes originating from indica and tropical japonica to the temperate japonica rice. Sci. Rep. 2022, 12, 15400. [Google Scholar] [CrossRef]
- Ookawa, T.; Hobo, T.; Yano, M.; Murata, K.; Ando, T.; Miura, H.; Asano, K.; Ochiai, Y.; Ikeda, M.; Nishitani, R.; et al. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 2010, 1, 132. [Google Scholar] [CrossRef] [Green Version]
- Mulsanti, I.W.; Yamamoto, T.; Ueda, T.; Samadi, A.F.; Kamahora, E.; Rumanti, I.A.; Thanh, V.C.; Adachi, S.; Suzuki, S.; Kanekatsu, M.; et al. Finding the superior allele of japonica-type for increasing stem lodging resistance in indica rice varieties using chromosome segment substitution lines. Rice 2018, 11, 25. [Google Scholar] [CrossRef]
- Hirano, K.; Okuno, A.; Hobo, T.; Ordonio, R.; Shinozaki, Y.; Asano, K.; Kitano, H.; Matsuoka, M. Utilization of Stiff Culm Trait of Rice smos1 Mutant for Increased Lodging Resistance. PLoS ONE 2014, 9, e96009. [Google Scholar] [CrossRef]
- Tu, B.; Tao, Z.; Wang, S.; Zhou, L.; Zheng, L.; Zhang, C.; Li, X.; Zhang, X.; Yin, J.; Zhu, X.; et al. Loss of Gn1a/OsCKX2 confers heavy-panicle rice with excellent lodging resistance. J. Integr. Plant Biol. 2022, 64, 23–38. [Google Scholar] [CrossRef]
- Jiang, S.; Yang, C.; Xu, Q.; Wang, L.; Yang, X.; Song, X.; Wang, J.; Zhang, X.; Li, B.; Li, H.; et al. Genetic Dissection of Germinability under Low Temperature by Building a Resequencing Linkage Map in japonica Rice. Int. J. Mol. Sci. 2020, 21, 1284. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, T.; Ishimaru, K. Identification and Functional Analysis of a Locus for Improvement of Lodging Resistance in Rice. Plant Physiol. 2004, 134, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Ookawa, T.; Yasuda, K.; Kato, H.; Sakai, M.; Seto, M.; Sunaga, K.; Motobayashi, T.; Tojo, S.; Hirasawa, T. Biomass Production and Lodging Resistance in ‘Leaf Star’, a New Long-Culm Rice Forage Cultivar. Plant Prod. Sci. 2010, 13, 58–66. [Google Scholar] [CrossRef]
- Lin, L.; Zhao, Y.; Liu, F.; Chen, Q.; Qi, J. Narrow leaf 1 (NAL1) regulates leaf shape by affecting cell expansion in rice (Oryza sativa L.). Biochem. Biophys. Res. Commun. 2019, 516, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Takai, T.; Adachi, S.; Taguchi-Shiobara, F.; Sanoh-Arai, Y.; Iwasawa, N.; Yoshinaga, S.; Hirose, S.; Taniguchi, Y.; Yamanouchi, U.; Wu, J.; et al. A natural variant of NAL1, selected in high-yield rice breeding programs, pleiotropically increases photosynthesis rate. Sci. Rep. 2013, 3, 2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Li, S.; Wang, L.; Ye, W.; Zeng, D.; Rao, Y.; Peng, Y.; Hu, J.; Yang, Y.; Xu, J.; et al. LSCHL4 from Japonica Cultivar, which is allelic to NAL1, increases yield of indica super rice 93-11. Mol. Plant 2014, 7, 1350–1364. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Zhang, F.; Wu, Y.; Zheng, T.; Wang, Y.; Zhao, X.; Cui, Y.; Chen, K.; Zhang, Q.; et al. SS1 (NAL1)- and SS2-Mediated Genetic Networks Underlying Source-Sink and Yield Traits in Rice (Oryza sativa L.). PLoS ONE 2015, 10, e0132060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, D.; Trijatmiko, K.; Tagle, A.; Sapasap, M.; Koide, Y.; Sasaki, K.; Tsakirpaloglou, N.; Gannaban, R.; Nishimura, T.; Yanagihara, S.; et al. NAL1 allele from a rice landrace greatly increases yield in modern indica cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 20431–20436. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Weng, X.; Yan, W.; Bai, X.; Xing, Y. Ghd7, a pleiotropic gene controlling flag leaf area in rice. Yi Chuan 2012, 34, 901–906. [Google Scholar] [CrossRef]
- Huang, X.; Qian, Q.; Liu, Z.; Sun, H.; He, S.; Luo, D.; Xia, G.; Chu, C.; Li, J.; Fu, X. Natural variation at the DEP1 locus enhances grain yield in rice. Nat. Genet. 2009, 41, 494–497. [Google Scholar] [CrossRef]
- Wang, B.; Wang, H. IPA1: A New “Green Revolution” Gene? Mol. Plant 2017, 10, 779–781. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z. Precision mapping of quantitative trait loci. Genetics 1994, 136, 1457–1468. [Google Scholar] [CrossRef]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef]
- Jiang, S.; Sun, S.; Bai, L.; Ding, G.; Wang, T.; Xia, T.; Jiang, H.; Zhang, X.; Zhang, F. Resequencing and variation identification of whole genome of the japonica rice variety “Longdao24” with high yield. PLoS ONE 2017, 12, e0181037. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Shenyang | Harbin | ||
|---|---|---|---|---|
| SN265 | LTH | SN265 | LTH | |
| Number of grains per panicle (GPP) | 162.07 | 126.43 | 161.07 | 128.32 |
| Number of primary branches (PBN) | 12.20 | 9.67 | 12.10 | 9.41 |
| Number of secondary branches (SBN) | 33.03 | 24.24 | 31.98 | 24.13 |
| Breaking strength of the basal internode (BS)/g | 883.09 | 366.50 | 879.25 | 367.22 |
| Basal culm diameter (CD)/cm | 4.54 | 4.10 | 4.38 | 4.06 |
| Basal culm thickness of the basal culm (CT)/cm | 1.40 | 0.58 | 1.38 | 0.59 |
| Groups | Location | Trait | Year | Chr. a | Interval | LOD b | Add. c | Var (%) d |
|---|---|---|---|---|---|---|---|---|
| F2 | Shenyang | BS | 2008 | 4 | RM470–RM559 | 4.80 | −160 | 23.4 |
| F2 | Shenyang | CT | 2008 | 4 | RM470–RM559 | 3.19 | −0.11 | 20.8 |
| F2 | Shenyang | CD | 2008 | 4 | RM470–RM559 | 3.34 | −0.15 | 16.2 |
| F2 | Shenyang | GPP | 2008 | 4 | RM470–RM559 | 3.21 | −6.1 | 16.8 |
| F2 | Shenyang | PBN | 2008 | 4 | RM470–RM559 | 5.90 | −1.6 | 26.4 |
| F2 | Shenyang | SBN | 2008 | 4 | RM470–RM559 | 3.40 | −2.5 | 16.1 |
| RILs | Shenyang | BS | 2010 | 4 | RM225–RM349 | 5.21 | −135 | 31.5 |
| RILs | Shenyang | CT | 2010 | 4 | RM225–RM349 | 4.12 | −0.18 | 24.5 |
| RILs | Shenyang | CD | 2010 | 4 | RM225–RM349 | 4.36 | −0.11 | 20.5 |
| RILs | Shenyang | GPP | 2010 | 4 | RM225–RM349 | 3.21 | −12.12 | 14.6 |
| RILs | Shenyang | PBN | 2010 | 4 | RM225–RM349 | 13.34 | −1. 1 | 43.2 |
| RILs | Shenyang | SBN | 2010 | 4 | RM225–RM349 | 4.56 | −5.47 | 26.6 |
| RILs | Harbin | BS | 2012 | 4 | RM225–RM349 | 3.92 | −157 | 28.7 |
| RILs | Harbin | CT | 2012 | 4 | RM225–RM349 | 4.12 | −0.18 | 24.5 |
| RILs | Harbin | CD | 2012 | 4 | RM225–RM349 | 3.76 | −0.14 | 22.7 |
| RILs | Harbin | GPP | 2012 | 4 | RM225–RM349 | 3.37 | −8.72 | 20.4 |
| RILs | Harbin | PBN | 2012 | 4 | RM225–RM349 | 3.19 | −0.98 | 19.4 |
| RILs | Harbin | SBN | 2012 | 4 | RM470–RM559 | 3.56 | −2.76 | 19.5 |
| Marker | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Primers 1 | CGCTTTCGGCATTCGTTATC | AGGTTGGATGTCGGCCATAA |
| Primers 2 | CATAGCAAGATATTGCGGCGTTT | TGCTTAATGCACATGGTATTTTTGC |
| Primers 3 | GGCTCAAACACCTAAAGAGCAAA | AACTCCTTCCAATCTCCGATCA |
| Primers 4 | TCTTGTGCAGATTAAAGCTTCTGG | GGGTTTGTTCCAGCATAGATTCC |
| Primers 5 | TGCAAAAAGTGCTGGTTCTGAATTT | TCCGGCAATGGTGTATATCAGGT |
| Primers 6 | CATGGCCCTGAAAACTGGAC | GATGCCTCCTCCCCTTCAGT |
| Primers 7 | CGCCATGTCGCTCCATCT | TCATGAGCACAGCAAAACTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Lai, Y.; Wang, L.; Zhao, M.; Wang, J.; Li, M.; Chi, L.; Lv, G.; Liu, Y.; Cui, Z.; et al. Isolation of a Novel QTL, qSCM4, Associated with Strong Culm Affects Lodging Resistance and Panicle Branch Number in Rice. Int. J. Mol. Sci. 2023, 24, 812. https://doi.org/10.3390/ijms24010812
Yang X, Lai Y, Wang L, Zhao M, Wang J, Li M, Chi L, Lv G, Liu Y, Cui Z, et al. Isolation of a Novel QTL, qSCM4, Associated with Strong Culm Affects Lodging Resistance and Panicle Branch Number in Rice. International Journal of Molecular Sciences. 2023; 24(1):812. https://doi.org/10.3390/ijms24010812
Chicago/Turabian StyleYang, Xianli, Yongcai Lai, Lizhi Wang, Minghui Zhao, Jiayu Wang, Mingxian Li, Liyong Chi, Guoyi Lv, Youhong Liu, Zhibo Cui, and et al. 2023. "Isolation of a Novel QTL, qSCM4, Associated with Strong Culm Affects Lodging Resistance and Panicle Branch Number in Rice" International Journal of Molecular Sciences 24, no. 1: 812. https://doi.org/10.3390/ijms24010812