
Analgesic Action of Acetaminophen via Kv7 Channels
 , , and
, , and 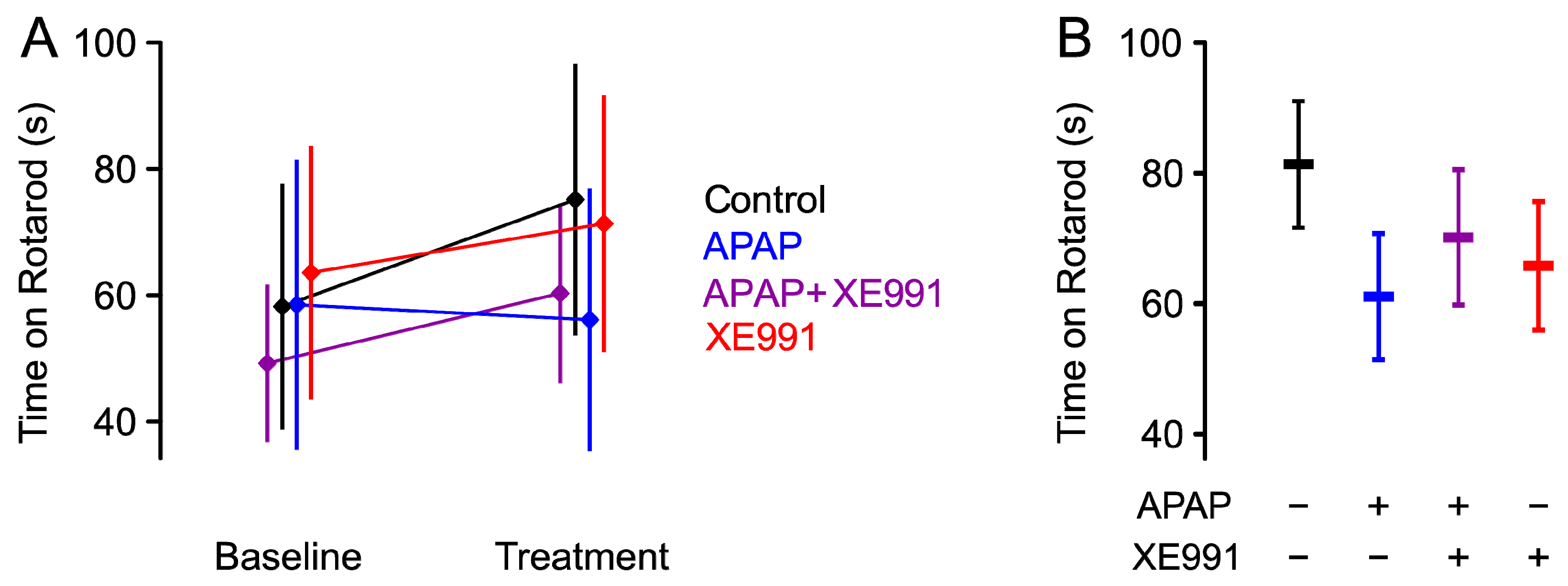{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Motor Coordination
2.2. Naive Condition
2.3. Inflamed State
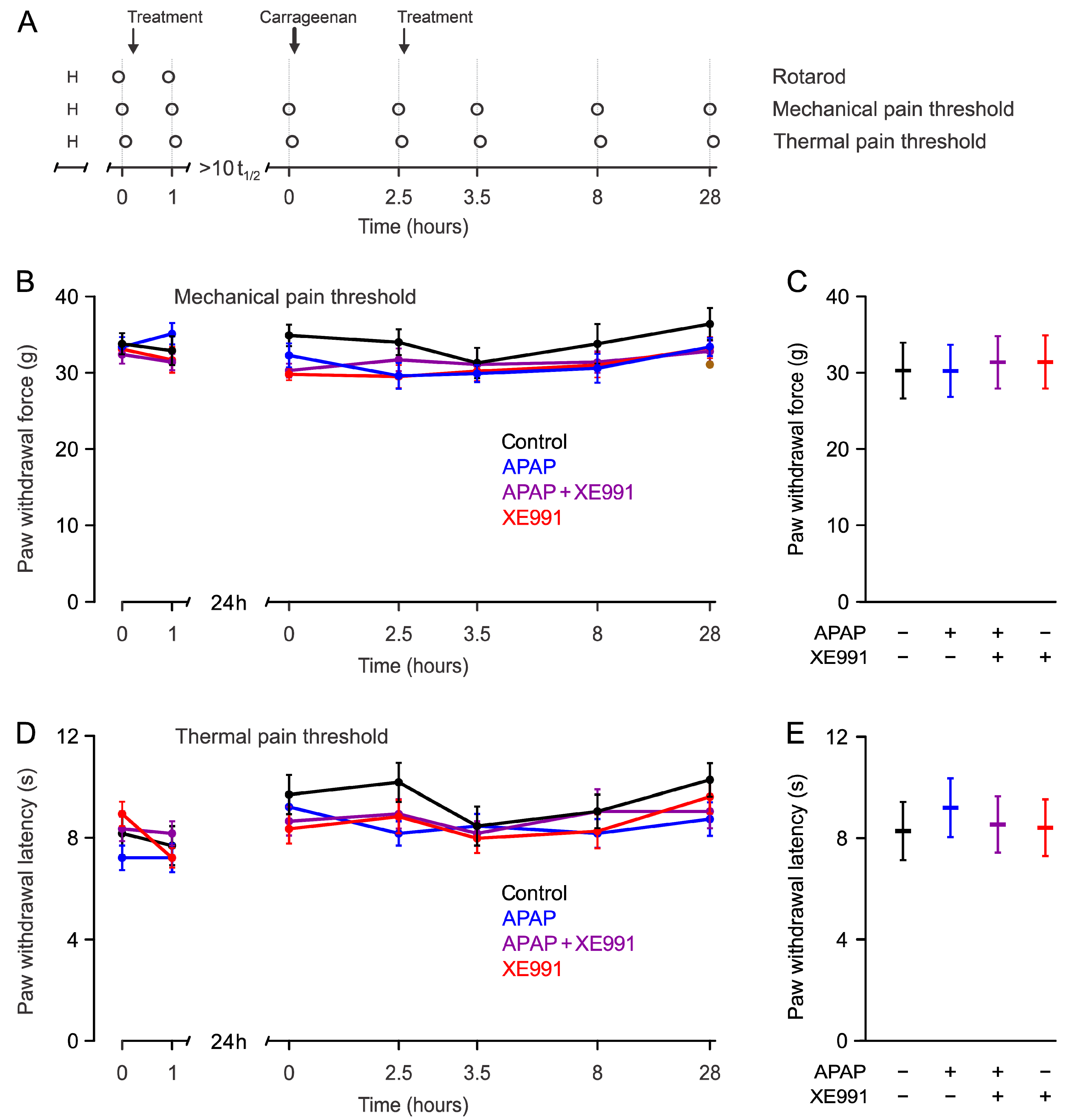
2.3.1. Mechanical Pain Thresholds during Hyperalgesia
2.3.2. Thermal Pain Thresholds during Hyperalgesia
2.4. Site of Action
2.5. CSF and Tissue Analysis
3. Discussion
3.1. Naive Condition
3.2. Inflamed Condition
3.3. XE991
3.4. Site of Action
3.5. Tissues Analysis
3.6. Limitations
3.7. Conclusion
4. Materials and Methods
4.1. Chemicals and Solutions
4.2. Preparation of Substances/Mixtures for Behavioral Experiments
4.3. Animals
4.4. Behavioral Testing
4.5. Experiment 1—APAP s.c. and Systemic Kv7 Inhibition
4.6. Experiment 2—APAP s.c. and Spinal Kv7 Inhibition
4.7. Pharmacokinetic Experiments
4.7.1. CSF Sample Preparation
4.7.2. Liver and Spinal Cord Sample Preparation
4.7.3. HPLC-UV-MS/MS Analysis
4.8. Sample Size Calculation and Statistical Analysis
4.8.1. Sample Size Calculation of Experiment 1—APAP s.c. and Systemic Kv7 Inhibition
4.8.2. Analysis of Experiment 1
4.8.3. Sample Size Calculation of Experiment 2—APAP s.c. and Spinal Kv7 Inhibition
4.8.4. Analysis of Experiment 2
4.8.5. Additional Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Model List of Essential Medicines—22nd List. 2021. Available online: https://www.who.int/publications-detail-redirect/WHO-MHP-HPS-EML-2021.02 (accessed on 19 July 2022).
- Przybyła, G.W.; Szychowski, K.A.; Gmiński, J. Paracetamol—An old drug with new mechanisms of action. Clin. Exp. Pharmacol. Physiol. 2020, 48, 3–19. [Google Scholar] [CrossRef]
- Larson, A.M. Acetaminophen hepatotoxicity. Clin. Liver Dis. 2007, 11, 525–548. [Google Scholar] [CrossRef] [PubMed]
- Rumack, B.H. Acetaminophen hepatotoxicity: The first 35 years. J. Toxicol. Clin. Toxicol. 2002, 40, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Laine, J.E.; Auriola, S.; Pasanen, M.; Juvonen, R.O. Acetaminophen bioactivation by human cytochrome P450 enzymes and animal microsomes. Xenobiotica 2009, 39, 11–21. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R.; Jaeschke, H. Metabolism and disposition of acetaminophen: Recent advances in relation to hepatotoxicity and diagnosis. Pharm. Res. 2013, 30, 2174–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz Ochoa, J.G.; Bucher, J.; Péry, A.R.R.; Zaldivar Comenges, J.M.; Niklas, J.; Mauch, K. A multi-scale modeling framework for individualized, spatiotemporal prediction of drug effects and toxicological risk. Front. Pharmacol. 2012, 3, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holme, J.A.; Dahlin, D.C.; Nelson, S.D.; Dybing, E. Cytotoxic effects of N-acetyl-p-benzoquinone imine, a common arylating intermediate of paracetamol and N-hydroxyparacetamol. Biochem. Pharmacol. 1984, 33, 401–406. [Google Scholar] [CrossRef]
- Andersson, D.A.; Gentry, C.; Alenmyr, L.; Killander, D.; Lewis, S.E.; Andersson, A.; Bucher, B.; Galzi, J.-L.; Sterner, O.; Bevan, S.; et al. TRPA1 mediates spinal antinociception induced by acetaminophen and the cannabinoid Δ(9)-tetrahydrocannabiorcol. Nat. Commun. 2011, 2, 551. [Google Scholar] [CrossRef] [Green Version]
- Jaeschke, H.; McGill, M.R.; Ramachandran, A. Oxidant stress, mitochondria, and cell death mechanisms in drug-induced liver injury: Lessons learned from acetaminophen hepatotoxicity. Drug Metab. Rev. 2012, 44, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Rotundo, L.; Pyrsopoulos, N. Liver injury induced by paracetamol and challenges associated with intentional and unintentional use. World J. Hepatol. 2020, 12, 125–136. [Google Scholar] [CrossRef]
- Sahoo, N.; Hoshi, T.; Heinemann, S.H. Oxidative modulation of voltage-gated potassium channels. Antioxid. Redox Signal. 2014, 21, 933–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.; Salzer, I.; Kronschläger, M.T.; Boehm, S. The paracetamol metabolite N-acetylp-benzoquinone imine reduces excitability in first- and second-order neurons of the pain pathway through actions on KV7 channels. Pain 2019, 160, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.L.; Hoshi, N. Modulation of Kv7 channels and excitability in the brain. Cell. Mol. Life Sci. 2017, 74, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Liu, Y.; Hu, F.; Yang, J.; Guo, X.; Hou, X.; Ju, C.; Wang, K. Activation of Neuronal Voltage-Gated Potassium Kv7/KCNQ/M-Current by a Novel Channel Opener SCR2682 for Alleviation of Chronic Pain. J. Pharmacol. Exp. Ther. 2021, 377, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Iwata, M.; Tsuchimori, N.; Matsumoto, T. Activation of peripheral KCNQ channels attenuates inflammatory pain. Mol. Pain 2014, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Løkken, P.; Skjelbred, P. Analgesic and anti-inflammatory effects of paracetamol evaluated by bilateral oral surgery. Br. J. Clin. Pharmacol. 1980, 10 (Suppl. S2), 253S–260S. [Google Scholar] [CrossRef] [Green Version]
- Bjørnsson, G.A.; Haanaes, H.R.; Skoglund, L.A. A randomized, double-blind crossover trial of paracetamol 1000 mg four times daily vs ibuprofen 600 mg: Effect on swelling and other postoperative events after third molar surgery. Br. J. Clin. Pharmacol. 2003, 55, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, X.; McNaughton, P.A. Inflammatory pain: The cellular basis of heat hyperalgesia. Curr. Neuropharmacol. 2006, 4, 197–206. [Google Scholar] [CrossRef]
- Kratzer, I.; Strazielle, N.; Saudrais, E.; Mönkkönen, K.; Malleval, C.; Blondel, S.; Ghersi-Egea, J.-F. Glutathione Conjugation at the Blood-CSF Barrier Efficiently Prevents Exposure of the Developing Brain Fluid Environment to Blood-Borne Reactive Electrophilic Substances. J. Neurosci. 2018, 38, 3466–3479. [Google Scholar] [CrossRef] [Green Version]
- Barrière, D.A.; Boumezbeur, F.; Dalmann, R.; Cadeddu, R.; Richard, D.; Pinguet, J.; Daulhac, L.; Sarret, P.; Whittingstall, K.; Keller, M.; et al. Paracetamol is a centrally acting analgesic using mechanisms located in the periaqueductal grey. Br. J. Pharmacol. 2020, 177, 1773–1792. [Google Scholar] [CrossRef]
- Boreus, L.O.; Sandberg, F. A comparison of some pharmacological effects of acetophenetidin and N-acetyl p-aminophenol. Acta Physiol. Scand. 1953, 28, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Kerckhove, N.; Mallet, C.; François, A.; Boudes, M.; Chemin, J.; Voets, T.; Bourinet, E.; Alloui, A.; Eschalier, A. Ca(v)3.2 calcium channels: The key protagonist in the supraspinal effect of paracetamol. Pain 2014, 155, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Mallet, C.; Barrière, D.A.; Ermund, A.; Jönsson, B.A.G.; Eschalier, A.; Zygmunt, P.M.; Högestätt, E.D. TRPV1 in Brain Is Involved in Acetaminophen-Induced Antinociception. PLoS ONE 2010, 5, e12748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinger-Gratz, P.P.; Ralvenius, W.T.; Neumann, E.; Kato, A.; Nyilas, R.; Lele, Z.; Katona, I.; Zeilhofer, H.U. Acetaminophen Relieves Inflammatory Pain through CB1 Cannabinoid Receptors in the Rostral Ventromedial Medulla. J. Neurosci. 2018, 38, 322–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshijima, H.; Hunt, M.; Nagasaka, H.; Yaksh, T. Systematic Review of Systemic and Neuraxial Effects of Acetaminophen in Preclinical Models of Nociceptive Processing. J. Pain Res. 2021, 14, 3521–3552. [Google Scholar] [CrossRef]
- Chen, Z.; Wei, H.; Pertovaara, A.; Wang, J.; Carlson, S. Anxiety- and activity-related effects of paracetamol on healthy and neuropathic rats. Pharmacol. Res. Perspect. 2018, 6, e00367. [Google Scholar] [CrossRef] [Green Version]
- Arendt-Nielsen, L.; Nielsen, J.C.; Bjerring, P. Double-blind, placebo controlled comparison of paracetamol and paracetamol plus codeine—A quantitative evaluation by laser induced pain. Eur. J. Clin. Pharmacol. 1991, 40, 241–247. [Google Scholar] [CrossRef]
- Flinn, F.B.; Brodie, B.B. The effect on the pain threshold of N-acetyl p-aminophenol, a product derived in the body from acetanilide. J. Pharmacol. Exp. Ther. 1948, 94, 76. [Google Scholar]
- van Amerongen, G.; Siebenga, P.; de Kam, M.L.; Hay, J.L.; Groeneveld, G.J. Effect profile of paracetamol, Δ9-THC and promethazine using an evoked pain test battery in healthy subjects. Eur. J. Pain 2018, 22, 1331–1342. [Google Scholar] [CrossRef]
- Tiippana, E.; Hamunen, K.; Kontinen, V.; Kalso, E. The effect of paracetamol and tropisetron on pain: Experimental studies and a review of published data. Basic Clin. Pharmacol. Toxicol. 2013, 112, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Pickering, G.; Creveaux, I.; Macian, N.; Pereira, B. Paracetamol and Pain Modulation by TRPV1, UGT2B15, SULT1A1 Genotypes: A Randomized Clinical Trial in Healthy Volunteers. Pain Med. 2020, 21, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Meller, S.T. Thermal and mechanical hyperalgesia: A distinct role for different excitatory amino acid receptors and signal transduction pathways? APS J. 1994, 3, 215–231. [Google Scholar] [CrossRef]
- Lewin, G.R.; Rueff, A.; Mendell, L.M. Peripheral and central mechanisms of NGF-induced hyperalgesia. Eur. J. Neurosci. 1994, 6, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Tal, M.; Bennett, G.J. Dextrorphan relieves neuropathic heat-evoked hyperalgesia in the rat. Neurosci. Lett. 1993, 151, 107–110. [Google Scholar] [CrossRef]
- Gierthmühlen, J.; Böhmer, J.; Attal, N.; Bouhassira, D.; Freynhagen, R.; Haanpää, M.; Hansson, P.; Jensen, T.S.; Kennedy, J.; Maier, C.; et al. Association of sensory phenotype with quality of life, functionality, and emotional well-being in patients suffering from neuropathic pain. Pain 2022, 163, 1378–1387. [Google Scholar] [CrossRef]
- Staud, R.; Weyl, E.E.; Price, D.D.; Robinson, M.E. Mechanical and Heat Hyperalgesia Highly Predict Clinical Pain Intensity in Patients With Chronic Musculoskeletal Pain Syndromes. J. Pain 2012, 13, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Bian, X.; Qu, Z.; Ma, Z.; Zhou, Y.; Wang, K.; Jiang, H.; Xie, J. Peptide hormone ghrelin enhances neuronal excitability by inhibition of Kv7/KCNQ channels. Nat. Commun. 2013, 4, 1435. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.L.; Kang, S.; Hoshi, N. XE991 and Linopirdine Are State-Dependent Inhibitors for Kv7/KCNQ Channels that Favor Activated Single Subunits. J. Pharmacol. Exp. Ther. 2017, 362, 177–185. [Google Scholar] [CrossRef]
- Passmore, G.M.; Selyanko, A.A.; Mistry, M.; Al-Qatari, M.; Marsh, S.J.; Matthews, E.A.; Dickenson, A.H.; Brown, T.A.; Burbidge, S.A.; Main, M.; et al. KCNQ/M currents in sensory neurons: Significance for pain therapy. J. Neurosci. 2003, 23, 7227–7236. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Li, L.; Liu, H.; Li, H.; Liu, Z.; Li, Z. KCNQ2/3/5 channels in dorsal root ganglion neurons can be therapeutic targets of neuropathic pain in diabetic rats. Mol. Pain 2018, 14, 1744806918793229. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Gao, H.; Jaffe, D.; Zhang, H.; Gamper, N. M-type K+ channels in peripheral nociceptive pathways. Br. J. Pharmacol. 2018, 175, 2158–2172. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, N.; Kohno, T. Analgesic Effect of Acetaminophen: A Review of Known and Novel Mechanisms of Action. Front. Pharmacol. 2020, 11, 580289. [Google Scholar] [CrossRef] [PubMed]
- Verneuil, J.; Brocard, C.; Trouplin, V.; Villard, L.; Peyronnet-Roux, J.; Brocard, F. The M-current works in tandem with the persistent sodium current to set the speed of locomotion. PLoS Biol. 2020, 18, e3000738. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Fang, D.; Liu, X.-D.; Li, S.; Ren, J.; Xing, G.-G. Suppression of KCNQ/M (Kv7) potassium channels in the spinal cord contributes to the sensitization of dorsal horn WDR neurons and pain hypersensitivity in a rat model of bone cancer pain. Oncol. Rep. 2015, 33, 1540–1550. [Google Scholar] [CrossRef] [Green Version]
- Cornish, B.H.; Ward, L.C.; Thomas, B.J. Measurement of extracellular and total body water of rats using multiple frequency bioelectrical impedance analysis. Nutr. Res. 1992, 12, 657–666. [Google Scholar] [CrossRef]
- Earl, R.A.; Zaczek, R.; Teleha, C.A.; Fisher, B.N.; Maciag, C.M.; Marynowski, M.E.; Logue, A.R.; Tam, S.W.; Tinker, W.J.; Huang, S.M.; et al. 2-Fluoro-4-pyridinylmethyl analogues of linopirdine as orally active acetylcholine release-enhancing agents with good efficacy and duration of action. J. Med. Chem. 1998, 41, 4615–4622. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.W.; Marion, C.J. Exotic Animal Formulary, 5th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018; ISBN 978-0-323-49803-6. [Google Scholar]
- Flecknell, P.A. Post-operative analgesia in rabbits and rodents. Lab Anim. 1992, 20, 34–37. [Google Scholar]
- Djouhri, L.; Malki, M.I.; Zeidan, A.; Nagi, K.; Smith, T. Activation of Kv7 channels with the anticonvulsant retigabine alleviates neuropathic pain behaviour in the streptozotocin rat model of diabetic neuropathy. J. Drug Target. 2019, 27, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.; Miller, M.C.; Caralopoulos, I.N.; Worden, M.S.; Brinker, T.; Gordon, Z.N.; Johanson, C.E.; Silverberg, G.D. Temporal course of cerebrospinal fluid dynamics and amyloid accumulation in the aging rat brain from three to thirty months. Fluids Barriers CNS 2012, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Ince, I.; Aksoy, M.; Ahiskalioglu, A.; Comez, M.; Dostbil, A.; Celik, M.; Yilmaz, I.; Mammadov, R.; Dogan, H.; Boztok Ozgermen, B.; et al. A Comparative Investigation of the Analgesic Effects of Metamizole and Paracetamol in Rats. J. Investig. Surg. 2015, 28, 173–180. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stampf, J.-L.; Ciotu, C.I.; Heber, S.; Boehm, S.; Fischer, M.J.M.; Salzer, I. Analgesic Action of Acetaminophen via Kv7 Channels. Int. J. Mol. Sci. 2023, 24, 650. https://doi.org/10.3390/ijms24010650
Stampf J-L, Ciotu CI, Heber S, Boehm S, Fischer MJM, Salzer I. Analgesic Action of Acetaminophen via Kv7 Channels. International Journal of Molecular Sciences. 2023; 24(1):650. https://doi.org/10.3390/ijms24010650
Chicago/Turabian StyleStampf, Jan-Luca, Cosmin I. Ciotu, Stefan Heber, Stefan Boehm, Michael J. M. Fischer, and Isabella Salzer. 2023. "Analgesic Action of Acetaminophen via Kv7 Channels" International Journal of Molecular Sciences 24, no. 1: 650. https://doi.org/10.3390/ijms24010650