
Growth Cessation and Dormancy Induction in Micropropagated Plantlets of Rheum rhaponticum ‘Raspberry’ Influenced by Photoperiod and Temperature

Abstract
:1. Introduction
2. Results
2.1. The Effect of Photoperiod and Temperature (Experiment 1)
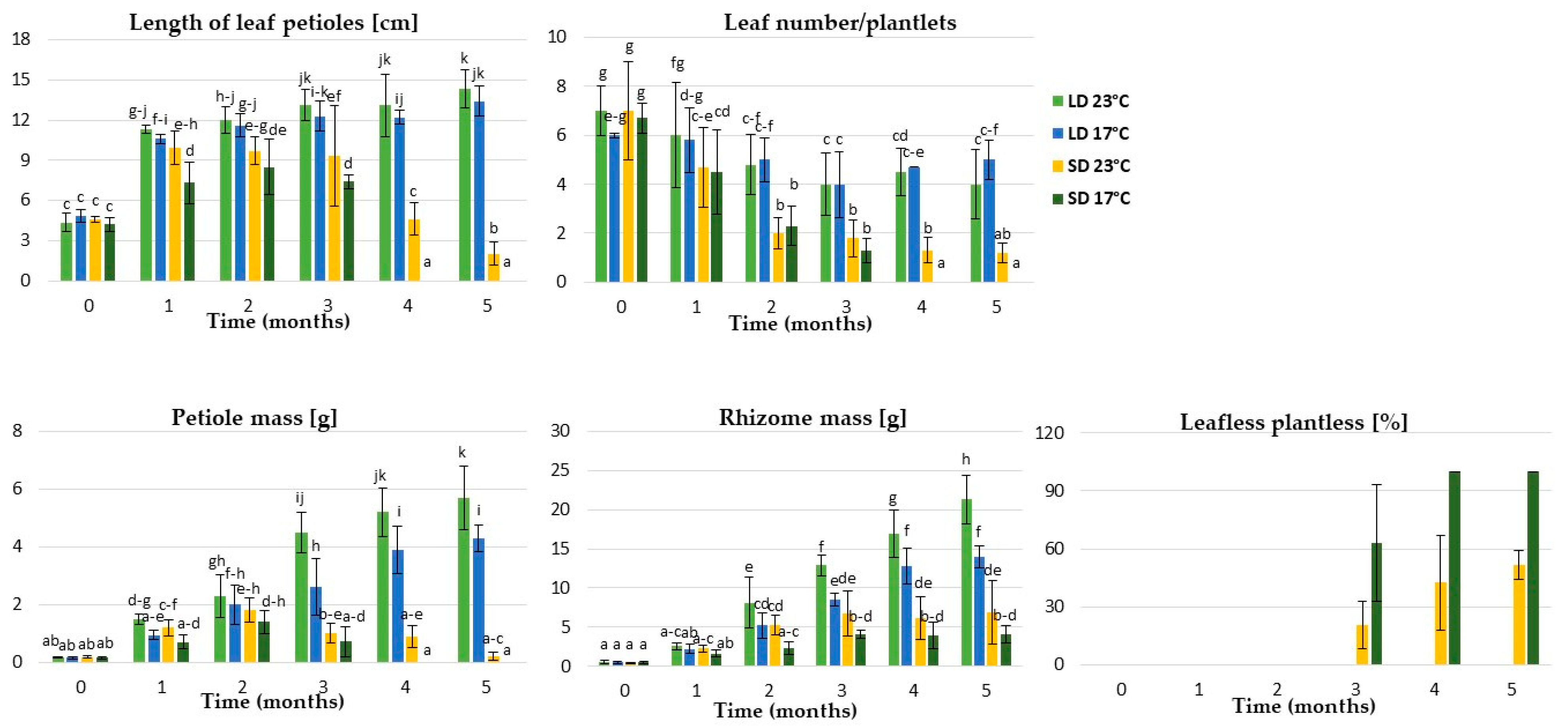
2.1.1. Physiological Responses of the Underground Buds during Dormancy Induction
2.1.2. Expression Analysis of Dormancy-Related Genes in Plantlets during Ex Vitro Growth under Different Photoperiods and Temperatures
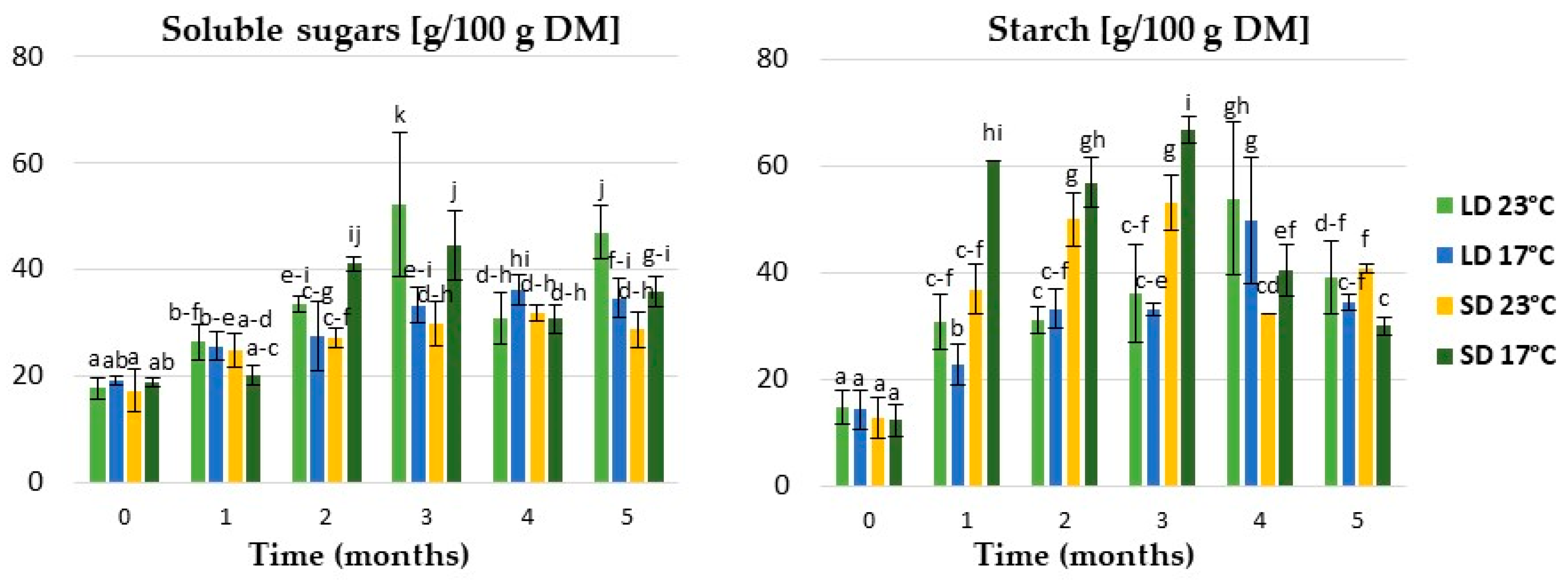
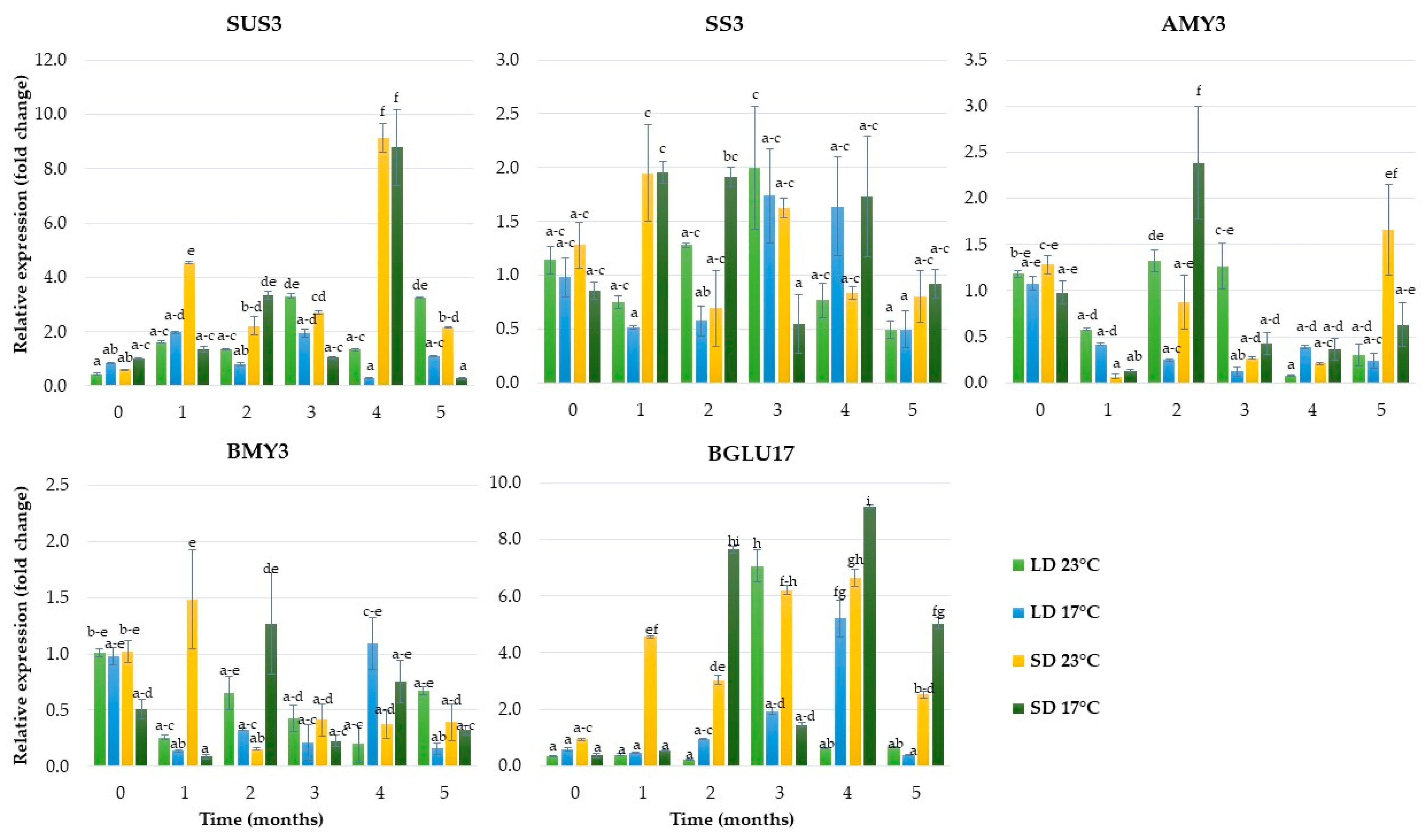
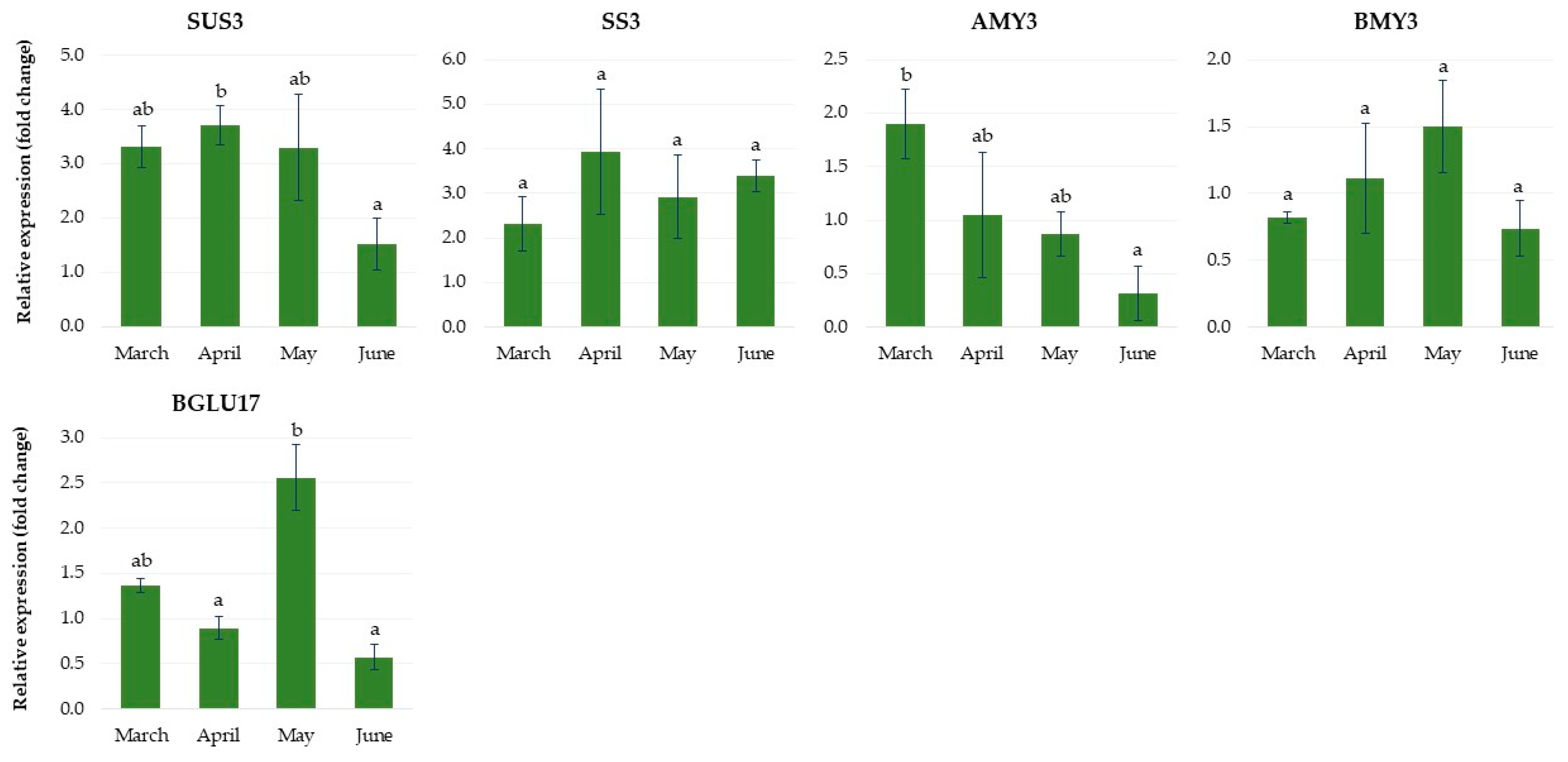
2.1.3. Expression of Genes Related to Carbohydrate Metabolism
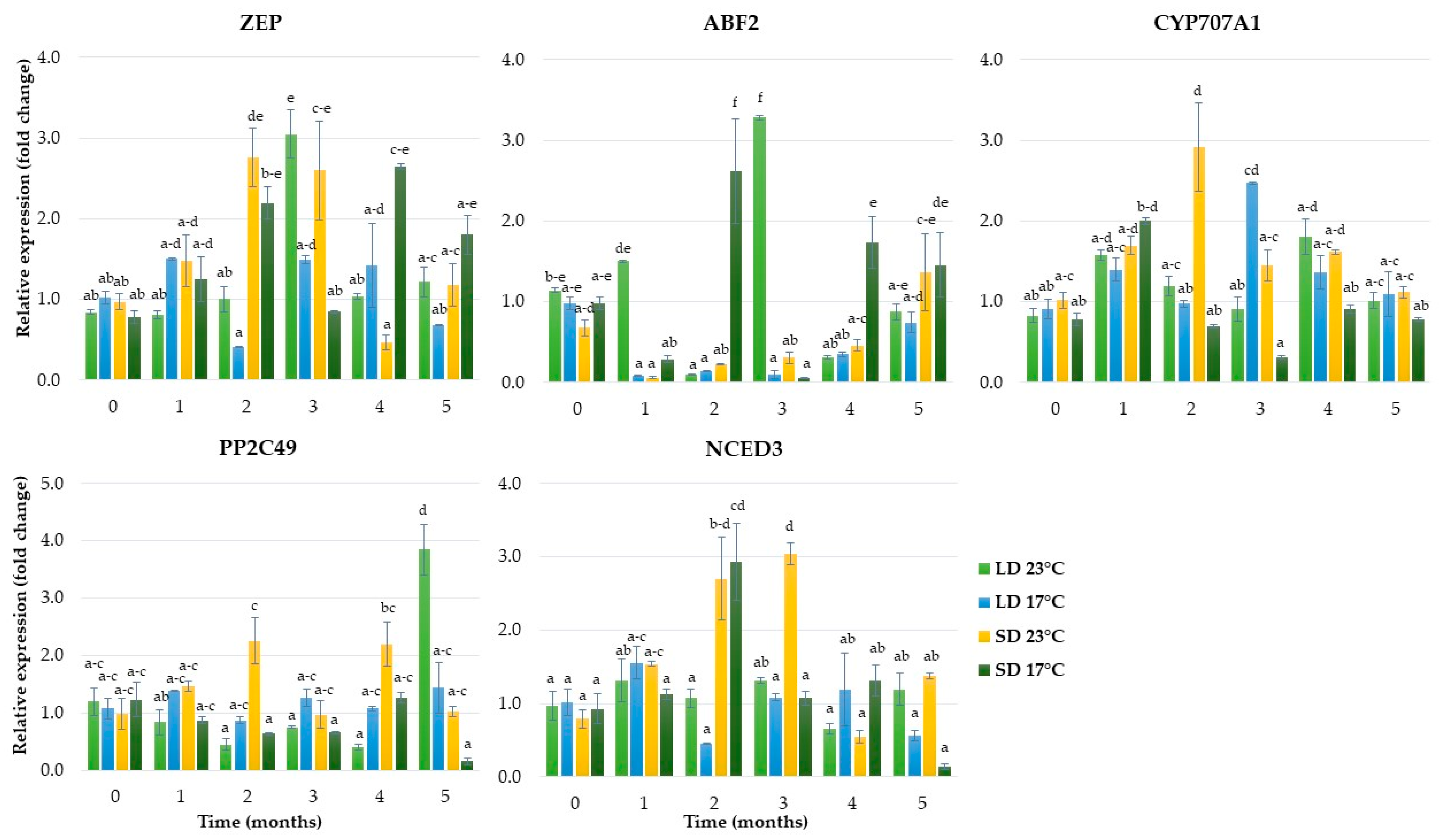
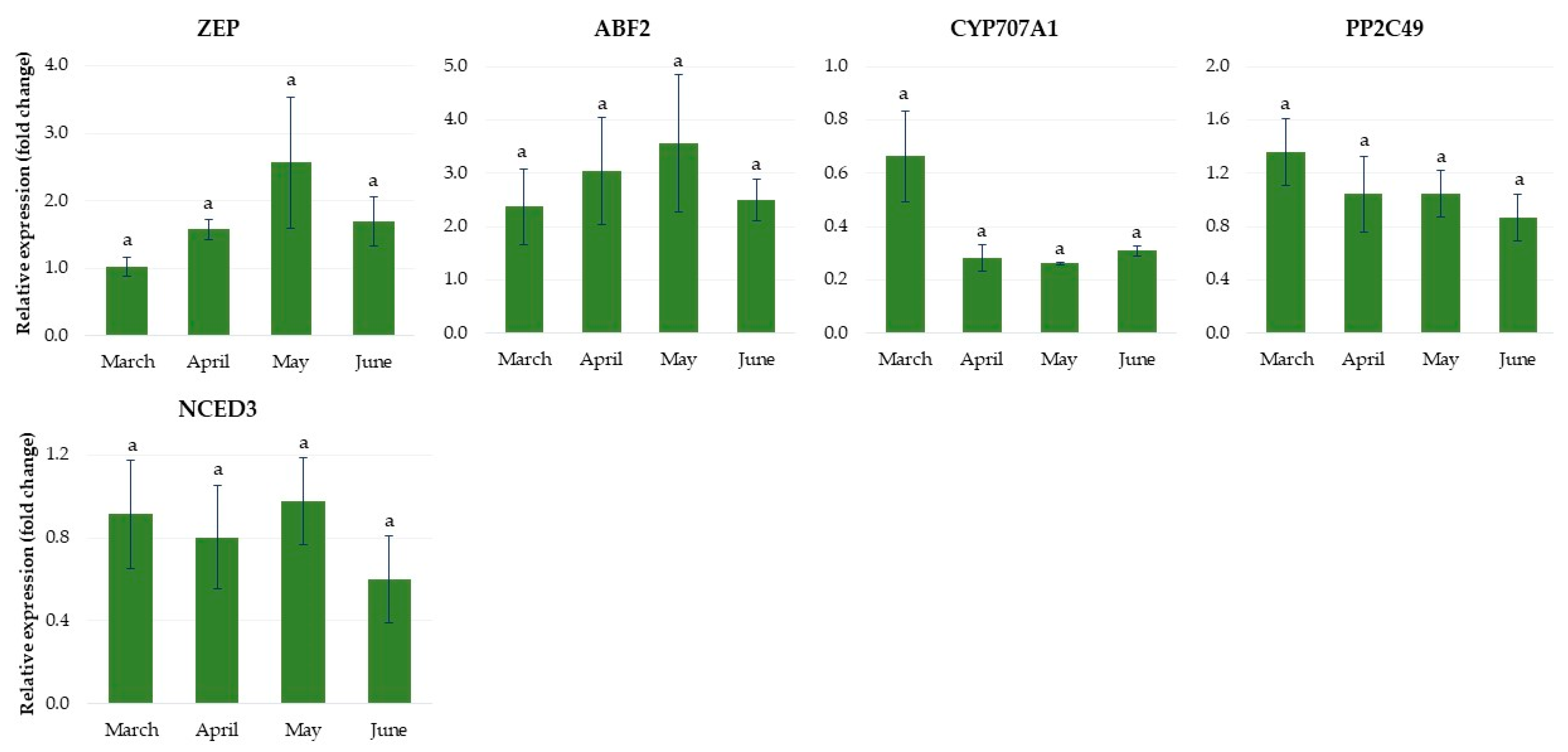
2.1.4. Expression of Genes Related to Abscisic Acid Metabolism
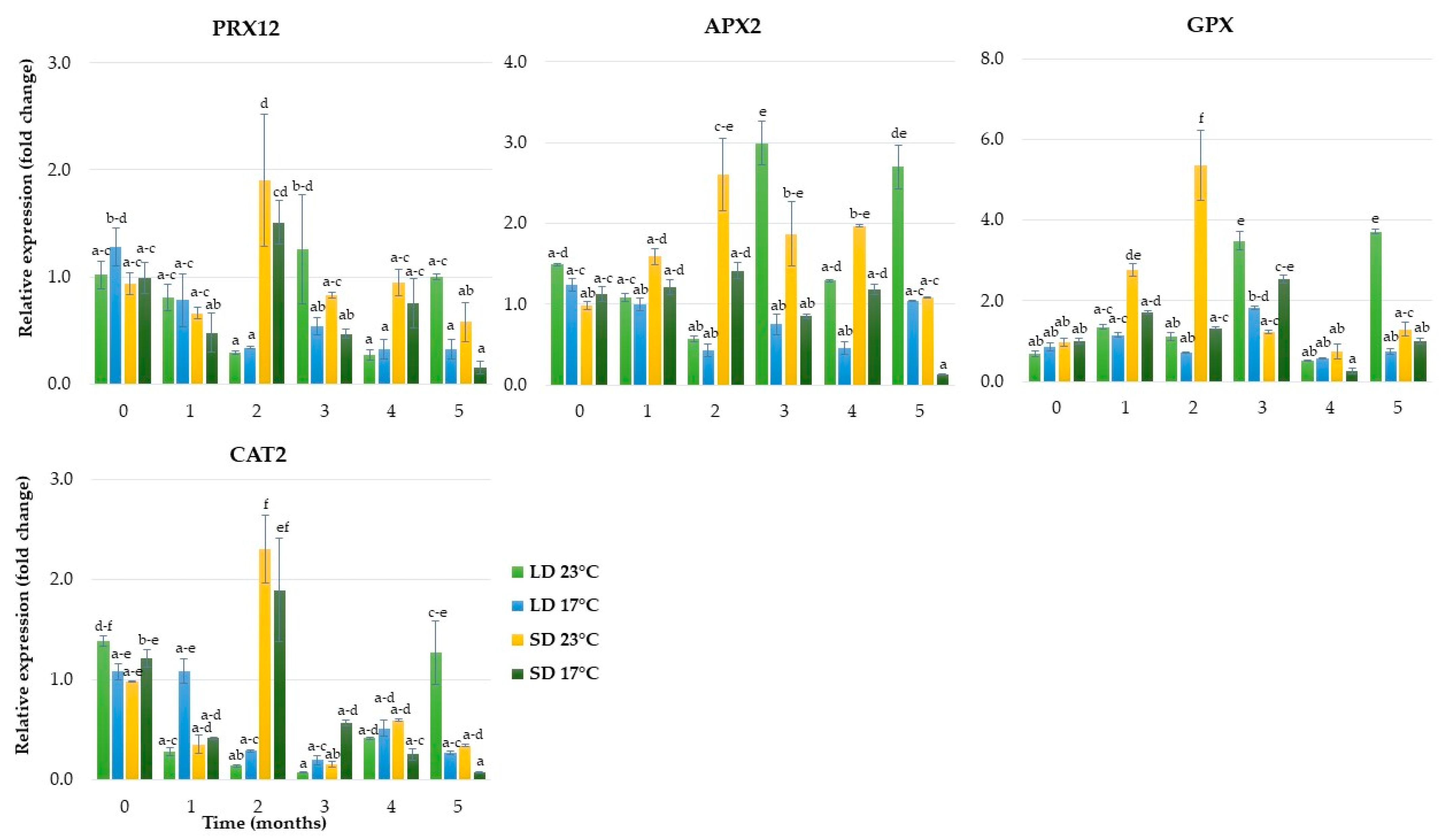
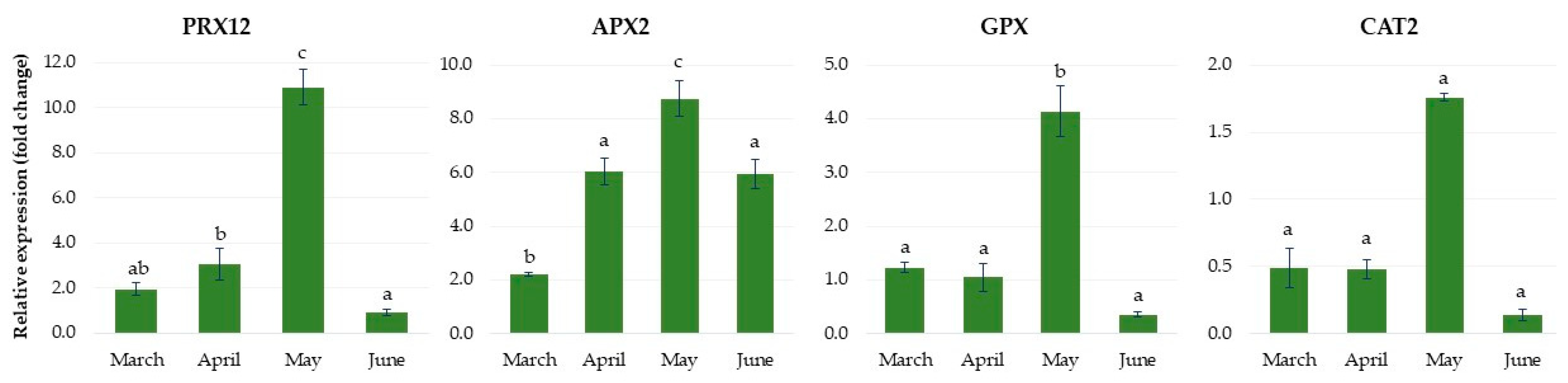
2.1.5. Expression of Genes Related to Antioxidant Metabolism
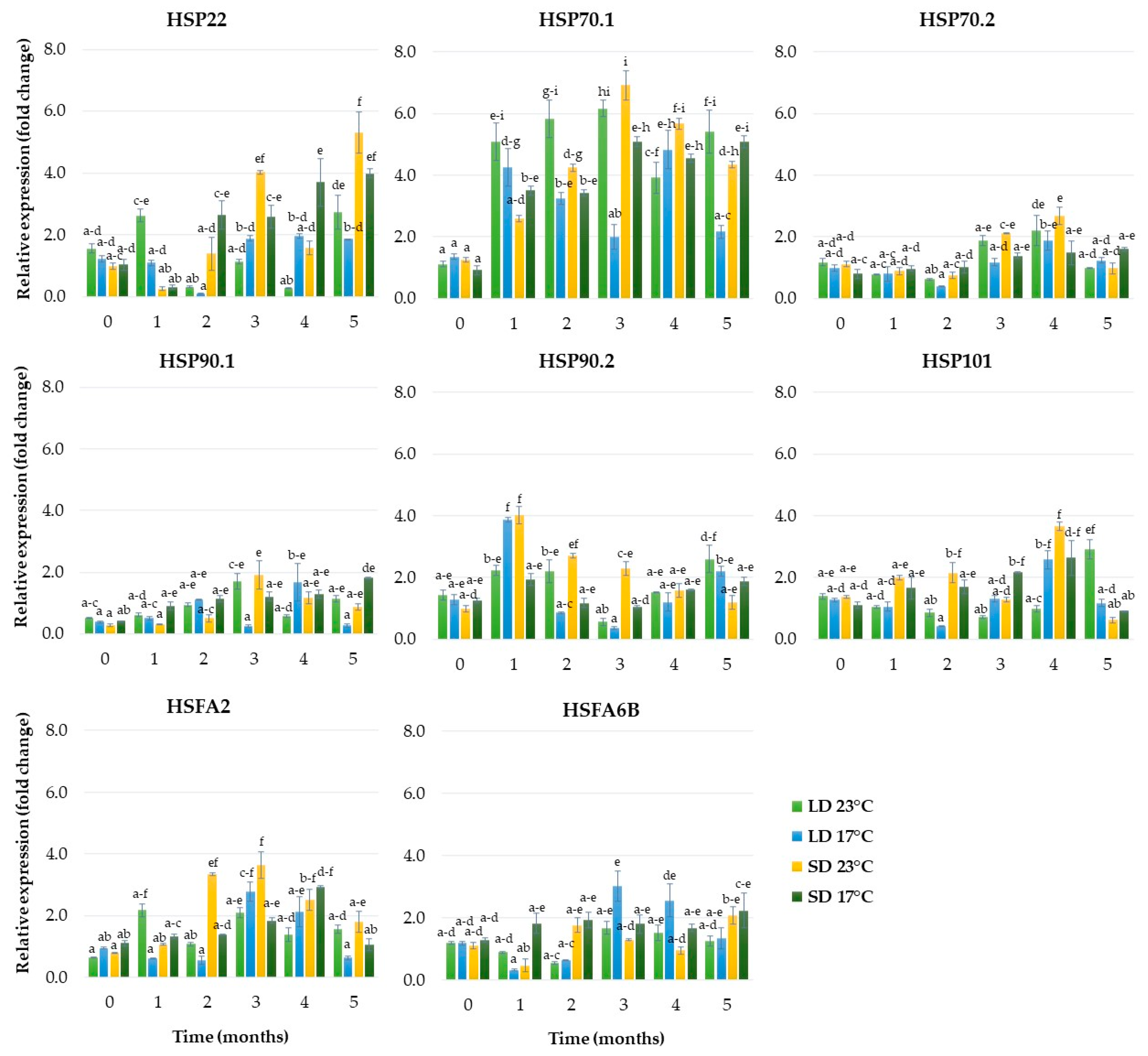
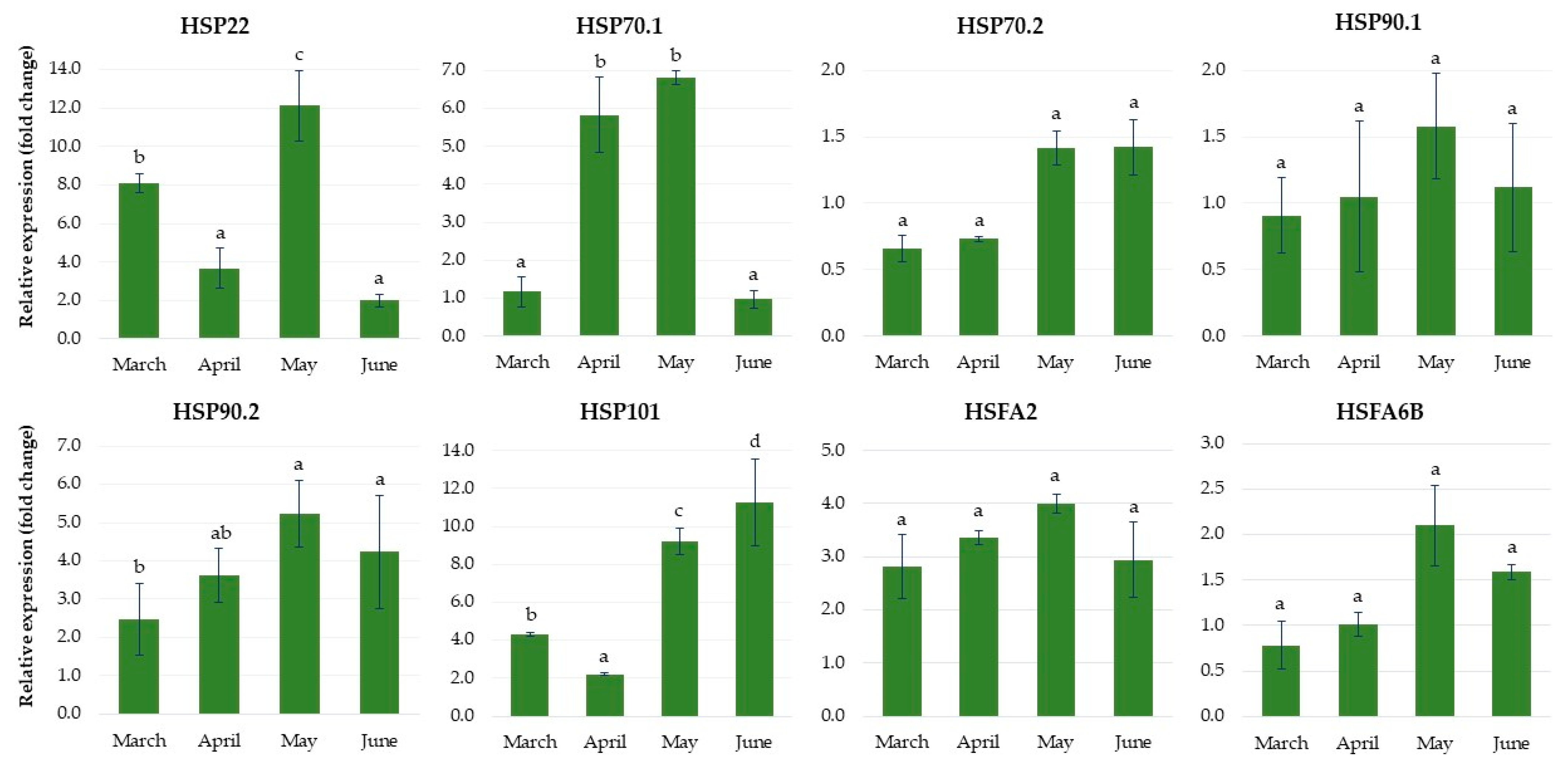
2.1.6. Expression of Genes Related to Heat Stress
2.2. The Effect of LD and Increased Temperature (Experiment 2)
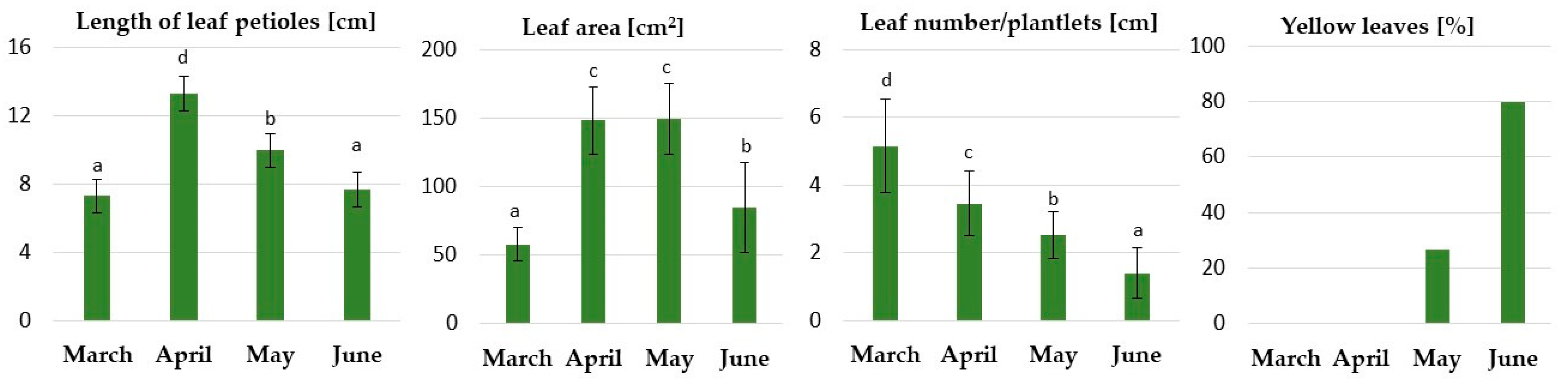
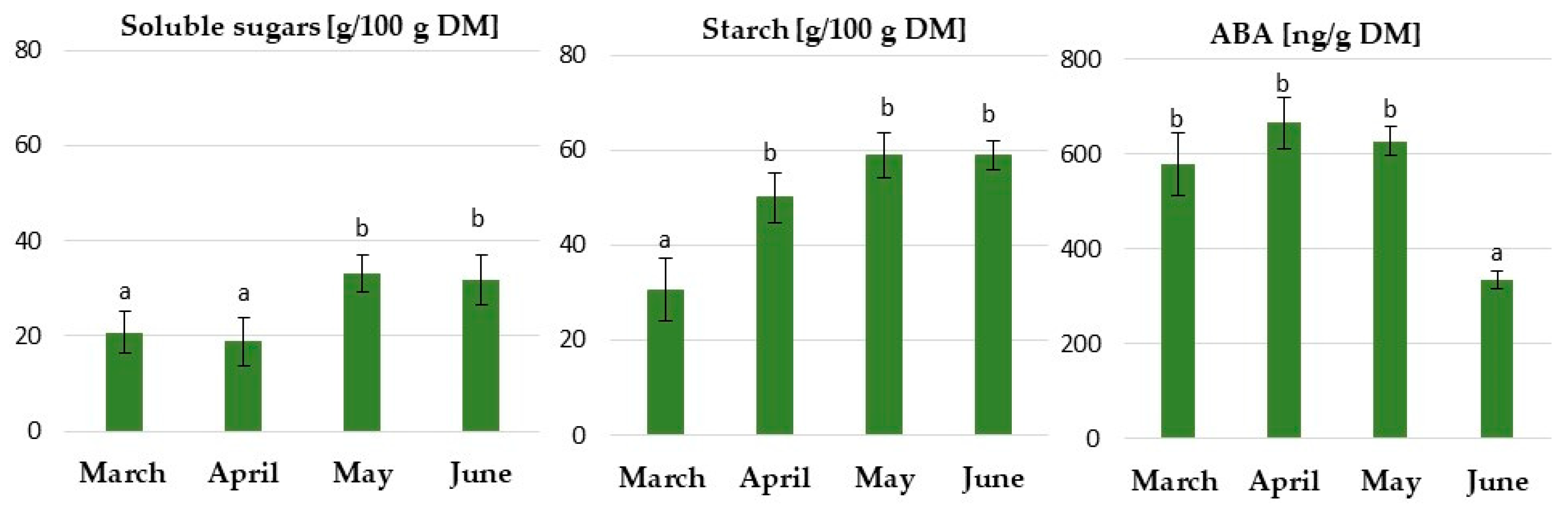
2.2.1. Physiological Responses of the Underground Buds during Dormancy Induction
2.2.2. Expression Analysis of Dormancy-Related Genes in Plantlets during Ex Vitro Growth in the Greenhouse
2.2.3. Expression of Genes Related to Carbohydrate Metabolism
2.2.4. Expression of Genes Related to Abscisic Acid Metabolism
2.2.5. Expression of Genes Related to Antioxidant Metabolism
2.2.6. Expression of Genes Related to Heat Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Ex Vitro Growth and Dormancy Induction of Plantlets
4.3. Measurements of Soluble Sugar Content
4.4. Measurements of Starch Content
4.5. Quantification of Abscisic Acid
4.6. Molecular Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krafczyk, N.; Kötke, M.; Lehnert, N.; Glomb, M.A. Phenolic composition of rhubarb. Eur. Food Res. Technol. 2008, 228, 187–196. [Google Scholar] [CrossRef]
- Takeoka, G.R.; Dao, L.; Harden, L.; Pantoja, A.; Kuhl, J.C. Antioxidant activity, phenolic and anthocyanin contents of various rhubarb (Rheum spp.) varieties. Int. J. Food Sci. Technol. 2013, 48, 172–178. [Google Scholar] [CrossRef]
- Kalisz, S.; Oszmiański, J.; Kolniak-Ostek, J.; Grobelna, A.; Kieliszek, M.; Cendrowski, A. Effect of a variety of polyphenols compounds and antioxidant properties of rhubarb (Rheum rhabarbarum). Food Sci. Technol. 2020, 118, 1–8. [Google Scholar] [CrossRef]
- Lassus, C.; Voipio, I. Micropropagation of rhubarb with special reference to weaning stage and subsequent growth. Agric. Sci. Finl. 1994, 3, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Kozak, D.; Sałata, A. Effect of cytokinins on in vitro multiplication of rhubarb (Rheum rhaponticum L.) ‘Karpov Lipskiego’ shoots and ex vitro acclimatization and growth. Acta Sci. Pol. Hortorum Cultus 2011, 10, 75–87. [Google Scholar]
- Thomas, J. Virus Identification and Development of Long-Term Management Strategies for the Rhubarb Industry; Final Report Hal Project VG05053; Department of Agriculture, Forestry and Fisheries: Pretoria, South Africa, 2011; 93p. [Google Scholar]
- Wojtania, A.; Mieszczakowska-Frąc, M. In vitro propagation method for production of phenolic-rich planting material of culinary rhubarb ‘Malinowy’. Plants 2021, 10, 1768. [Google Scholar] [CrossRef]
- Wojtania, A.; Markiewicz, M.; Waligórski, P. Regulation of the bud dormancy development and release in micropropagated rhubarb ‘Malinowy’. Int. J. Mol. Sci. 2022, 23, 1480. [Google Scholar] [CrossRef]
- Anderson, J.V.; Horvath, D.P.; Chao, W.S.; Foley, M.E. Bud dormancy in perennial plants: A mechanism for survival. In Dormancy and Resistance in Harsh Environments; Topics in Current Genetics 21; Lubzens, E., Cerda, J., Clark, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 69–90. [Google Scholar]
- Yang, Q.; Gao, Y.; Wu, X.; Moriguchi, T.; Bai, S.; Teng, Y. Bud endodormancy in deciduous fruit trees: Advances and prospects. Hortic. Res. 2021, 8, 139. [Google Scholar] [CrossRef]
- Langens-Gerrits, M.M.; Miller, W.B.M.; Croes, A.F.; de Klerk, G.J. Effect of low temperature on dormancy breaking and growth after planting in lily bulblets regenerated in vitro. Plant Growth Regul. 2003, 40, 267–275. [Google Scholar] [CrossRef]
- Podwyszyńska, M. The mechanisms of in vitro storage organ formation in ornamental geophytes. Floric. Ornam. Plant Biotechnol. 2012, 6, 9–23. [Google Scholar]
- Marković, M.; Trifunović Momčilow, M.; Uzelac, B.; Jevremović, S.; Subotić, A. Bulb dormancy in vitro—Fritilaria meleagris: Initiation, release and physiological parameters. Plants 2021, 10, 902. [Google Scholar] [CrossRef]
- Vahdati, K.; Leslie, C.; Zamani, Z.; McGranahan, G. Rooting and acclimatization of in vitro-grown shoots from mature trees of three persian walnut cultivars. Hortic. Sci. 2004, 39, 324–327. [Google Scholar] [CrossRef]
- Gabryszewska, E. Effect of temperature on the growth and dormancy of tissue-cultured herbaceus peony shoots. Acta Hortic. 2006, 725, 471–475. [Google Scholar] [CrossRef]
- Wojtania, A.; Dziurka, M.; Skrzypek, E. In vitro rooting response of yellow-flowered magnolia in relations to phenolic acid contents. Agronomy 2020, 10, 1880. [Google Scholar] [CrossRef]
- Gabryszewska, E.; Kawa-Miszczak, L. Rooting in vitro and acclimatisation in the greenhouse of herbaceous peony plantlets. Biotechnologia 2010, 2, 172–179, (English Abstract). [Google Scholar]
- Fu, Z.; Xu, M.; Wang, H.; Wang, E.; Li, Y.; Wang, L.; Gao, J.; Zhang, J.; Yuan, X.; Zhang, H. Analysis of the transcriptome and related physiological indicators of tree peony (Paeonia suffruticosa Andr.) plantlets before and after rooting in vitro. Plant Cell Tissue Organ Cultlure 2021, 147, 529–543. [Google Scholar] [CrossRef]
- Fennel, A.; Hoover, E. Photoperiod influences growth, bud dormancy, and cold acclimation in Vitis labruscana and V. riparia. J. Am. Soc. Hortic. Sci. 1991, 116, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Junttila, O.; Ernstsen, A.; Heino, P.; Palva, E.T. Photoperiodic control of growth, cold acclimation and dormancy develpment in silver birch (Betula pendula) ecotypes. Physiol. Plant. 2003, 117, 206–212. [Google Scholar] [CrossRef]
- Kalcsits, L.; Silim, S.; Tanino, K. Warm temperature accelerates short-photoperiod-induced growth cessation and dormancy induction in hybrid poplar (Populus × spp.). Trees 2009, 23, 971–979. [Google Scholar] [CrossRef]
- Rohne, A.; Bastien, C.; Boerjan, W. Temperature signals contribute to the timing of photoperiodic growth cessation and bud set in polplar. Tree Physiol. 2011, 31, 472–482. [Google Scholar]
- Heide, O.M.; Prestrud, A.K. Low temperature, but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiol. 2005, 25, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heide, O.M. Temperature rather than photoperiod controls growth cessation and dormancy in Sorbus species. J. Exp. Bot. 2011, 62, 5397–5404. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.J.; Tisdall, L. Acclimatization strategies for micropropagated plants. In Micropropagation of Woody Plants; Ahuja, M.R., Ed.; Springer: Dordrecht, The Netherlands, 1993; pp. 153–166. [Google Scholar]
- Pospíšilová, J.; Tichá, I.; Kadleček, P.; Haisel, D.; Plzáková, Š. Acclimatization of micropropagated plants to ex vitro conditions. Biol. Plant. 1999, 42, 481–497. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Hernández, J.A.; Díaz-Vivancoos, P. Antioxidant system: The hub of bud dormancy regulation in Prunus sp. Sci. Hortic. 2022, 305, 111396. [Google Scholar] [CrossRef]
- Doğramaci, M.; Horvath, D.P.; Anderson, J.V. Meta-analysis identifies molecular markers for endodormancy in crown buds of leafy spurge: A herbaceous perennial. In Advances in Plant Dormancy; Anderson, J.V., Ed.; Springer: Cham, Switzerland, 2015; pp. 197–219. [Google Scholar]
- Anderson, J.V.; Gesch, R.W.; Jia, Y.; Chao, W.S.; Horvath, D.P. Seasonal shifts in dormancy status carbohydrate metabolism and related gene expression in crown buds of leafy spurge. Plant Cell Environ. 2005, 28, 1567–1578. [Google Scholar] [CrossRef]
- Hemmati, H.; Gupta, D.; Basu, C. Molecular physiology of heat stress responses in plants. In Elucidation of Abiotic Stress Signaling in Plants; Pandey, G.K., Ed.; Springer: New York, NY, USA, 2015; pp. 109–129. [Google Scholar]
- Wang, L.; Ma, K.B.; Lu, Z.G.; Ren, S.X.; Jiang, H.R.; Cui, J.W.; Chen, G.; Teng, N.J.; Lam, H.M.; Jin, B. Differential physiological, transcriptomic and metabolomic responses of Arabidopsis leaves under prolonged warming and heat shock. BMC Plant Biol. 2020, 20, 86. [Google Scholar] [CrossRef]
- Schrader, W.L. Rhubarb Production in California; UCANR Publications: San Diego, CA, USA, 2000. [Google Scholar]
- Chao, W.S.; Foley, M.E.; Horvath, D.P.; Anderson, J.V. Signals regulating dormancy in vegetative buds. Int. J. Plant Dev. Biol. 2007, 1, 49–56. [Google Scholar]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar signals and the control of plant growth and development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef]
- Gibon, Y.; Pyl, E.T.; Sulpice, R.; Lunn, J.E.; Höhne, M.; Günther, M.; Stitt, M. Adjusment of growth, starch turnover, protein content and centralmetabolism to a decrease of the carbon supply when Arabidopsis is grown in very short photoperiod. Plant Cell Environ. 2009, 32, 859–874. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome profiles reveal the crucial roles of hormone and sugar in the bud dormancy of Prunus mume. Sci. Rep. 2018, 8, 5090. [Google Scholar] [CrossRef] [Green Version]
- Wind, J.; Smeekens, S.; Hanson, J. Sucrose: Metabolite and signaling molecule. Phytochemistry 2010, 71, 1610–1614. [Google Scholar] [CrossRef]
- Chao, W.S.; Serpe, M.D. Changes in the expression of carbohydrate metabolism genes during three phases of bud dormancy in leafy spurge. Plant Mol. Biol. 2010, 73, 227–239. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Shi, X.; Shao, L.; Xu, T.; Xia, Y.; Li, D.; Zhang, J. Chilling requirement validation and physiological and molecular responses of the bud endodormancy release in Paeonia lactiflora ‘Meiju’. Pall. Int. J. Mol. Sci. 2021, 22, 8382. [Google Scholar] [CrossRef]
- Zheng, C.; Halaly, T.; Acheampong, A.K.; Takebayashi, Y.; Jikumaru, Y.; Kamiya, Y.; Or, E. Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism. J. Exp. Bot. 2015, 66, 1527–1542. [Google Scholar] [CrossRef]
- Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant hormone-mediated regulation of heat tolerance in response to global climate change. Front. Plant Sci. 2021, 11, 627969. [Google Scholar] [CrossRef]
- Pan, W.; Liang, J.; Sui, J.; Li, J.; Liu, C.; Xin, Y.; Zhand, Y.; Wang, S.; Zhao, Y.; Zhang, J.; et al. ABA and bud dormancy in perennials: Current knowledge and future perspective. Genes 2021, 12, 1635. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Z.; Du, P.; Xiao, W.; Tan, Q.; Chen, X.; Li, L.; Gao, D. Expression of ABA metabolism-related genes suggests similarities and differences between seed dormancy and bud dormancy of peach (Prunus persica). Front. Plant Sci. 2016, 6, 1248. [Google Scholar] [CrossRef] [Green Version]
- Tuan, P.A.; Bai, S.; Saito, T.; Ito, A.; Moriguchi, T. Dormancy-Associated MADS-Box (DAM) and the abscisic acid pathway regulate pear endodormancy through a feedback mechanism. Plant Cell Physiol. 2017, 58, 1378–1390. [Google Scholar]
- Rubio, S.; Noriega, X.; Pérez, F.J. ABA promotes starch synthesis and storage metabolism in dormant grapevine buds. J. Plant Physiol. 2019, 234, 1–8. [Google Scholar]
- Miller, G.; Shulaev, V.; Mittlera, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2008, 133, 481–489. [Google Scholar] [CrossRef]
- Sharma, M.; Pandey, G.K. Elucidation of Abiotic Stress Signaling in Plants; Springer: New York, NY, USA, 2015. [Google Scholar]
- Campbell, J.L.; Klueva, N.Y.; Zheng, H.G.; Nieto-Sotelo, J.; Ho, T.H.; Nguyen, H.T. Cloning of new members of heat shock protein HSP101 gene family in wheat (Triticum aestivum (L.) Moench) inducible by heat, dehydration, and ABA. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2001, 1517, 270–277. [Google Scholar] [CrossRef]
- Tian, F.; Hu, X.L.; Yao, T.; Yang, X.; Chen, J.G.; Lu, M.Z.; Zhang, J. Recent advances in the roles of HSFs and HSPs in heat stress response in woody plants. Front. Plant Sci. 2021, 1319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Pei, X.; Zhang, R.; Lu, Y.; Zheng, J.; Zheng, Y. Molecular characterization and expression analysis of small heat shock protein 17.3 gene from Sorbus pohuashanensis (Hance) Hedl. In Response to Abiotic Stress. Mol. Biol. Rep. 2020, 47, 9325–9335. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Sun, Y.; Sun, A.Q.; Yi, S.Y.; Qin, J.; Li, M.H.; Liu, J. The involvement of chloroplast HSP100/ClpB in the acquired thermotolerance in tomato. Plant Mol. Biol. 2006, 62, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Young, T.E.; Ling, J.; Geisler-Lee, C.J.; Tanguay, R.L.; Caldwell, C.; Gallie, D.R. Developmental and thermal regulation of the maize heat shock protein, HSP101. Plant Physiol. 2001, 127, 777–791. [Google Scholar] [CrossRef]
- Katiyar-Agarwal, S.; Agarwal, M.; Grover, A. Heat-tolerant basmati rice engineered by over-expression of hsp101. Plant Mol. Biol. 2003, 51, 677–686. [Google Scholar] [CrossRef]
- Bi, H.; Zhao, Y.; Li, H.; Liu, W. Wheat heat shock factor TaHsfA6f increases ABA levels and enhances tolerance to multiple abiotic stresses in transgenic plants. Int. J. Mol. Sci. 2020, 21, 3121. [Google Scholar] [CrossRef]
- Erdayani, E.; Nagarajan, R.; Grant, N.P.; Gill, K.S. Genome-wide analysis of the HSP101/CLPB gene family for heat tolerance in hexaploid wheat. Sci. Rep. 2020, 10, 3948. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, S.K.; Dalal, M.; Chinnusamy, V.; Bansal, K.C. Differential regulation of genes coding for organelle and cytosolic ClpATPases under biotic and abiotic stresses in wheat. Front. Plant Sci. 2016, 7, 929. [Google Scholar] [CrossRef] [Green Version]
- Albertos, P.; Dündar, G.; Schenk, P.; Carrera, S.; Cavelius, P.; Sieberer, T.; Poppenberger, B. Transcription factor BES1 interacts with HSFA1 to promote heat stress resistance of plants. EMBO J. 2022, 41, e108664. [Google Scholar] [CrossRef]
- Firmansyah, A.N.; Argosubekti, N. A review of heat stress signaling in plants. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 484. [Google Scholar]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Medina, E.; Kim, S.H.; Yun, M.; Choi, W.G. Recapitulation of the function and role of ROS generated in response to heat stress in plants. Plants 2021, 10, 371. [Google Scholar] [CrossRef]
- Soengas, P.; Rodríguez, V.M.; Velasco, P.; Cartea, M.E. Effect of temperature stress on antioxidant defenses in Brassica oleracea. ACS Omega 2018, 3, 5237–5243. [Google Scholar] [CrossRef]
- Mala, D.; Awasthi, S.; Sharma, N.K.; Swarnkar, M.K.; Shankar, R.; Kumar, S. Comparative transcriptome analysis of Rheum australe, an endangered medicinal herb, growing in its natural habitat and those grown in controlled growth chambers. Sci. Rep. 2021, 11, 3702. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Dobrev, P.I.; Kamınek, M. Fast and efficient separation of cytokinins from auxin and abscisic acid and their purification using mixed-mode solid-phase extraction. J. Chromatogr. A 2002, 950, 21–29. [Google Scholar] [CrossRef]
- Stefancic, M.; Stampar, F.; Veberic, R.; Osterc, G. The levels of IAA, IAAsp and some phenolics in cherry rootstock ‘GiSelA 5’ leafy cuttings pretreated with IAA and IBA. Sci. Hortic. 2007, 112, 399–405. [Google Scholar] [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Beatrice, P.; Chiatante, D.; Scippa, G.S.; Montagnoli, A. Photoreceptors’ gene expression of Arabidopsis thaliana grown with biophilic LED-sourced lighting systems. PLoS ONE 2022, 17, e0269868. [Google Scholar] [CrossRef]
- Łabuz, J.; Sztatelman, O.; Banaś, A.K.; Gabryś, H. The expression of phototropins in Arabidopsis leaves: Developmental and light regulation. J. Exp. Bot. 2012, 63, 1763–1771. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| SUS3 | 5′-TCGAAATTGGAGCGTCGTGA-3′ |
| 5′-CAGTTTTCACCAAGTCGCGG-3′ | |
| SS3 | 5′-GGCTCGGCTTGTTCTAACCT-3′ |
| 5′-TGTGTCAGTCCACATGGCTC-3′ | |
| AMY3 | 5′-CAGCGGTCTTCTTCGACCAT-3′ |
| 5′-GCCCTGGTCCGATCTTCATT-3′ | |
| BMY3 | 5′-CAGGTACGAGGCTATCGCAG-3′ |
| 5′-TCAGGTGATTGGTGCTCGTC-3′ | |
| BGLU17 | 5′-GAACTCAGCCACTGAGCCAT-3′ |
| 5′-GAGTTGGACTGTAGCGGCAT-3′ | |
| ZEP | 5′-GGCACAAGGGATCACGAACT-3′ |
| 5′-CCTTGGAGGAGAATCGAATGG-3′ | |
| ABF2 | 5′-TCGTTGACTCTGCCTCGAAC-3′ |
| 5′-CCTGAGCCACCTGAGACAAG-3′ | |
| CYP707A1 | 5′-CACTGAAGAGCAAGAGGCTATA-3′ |
| 5′-TTCTTGGTATCTGCCCAACTC-3′ | |
| PP2C49 | 5′-GATCGACGACCTATCCATGCA-3′ |
| 5′-GGTCCTCCATGGCCATCA-3′ | |
| NCED3 | 5′-TCGAAGCAGGGATGGTCAAC-3′ |
| 5′-CCTGAGACTTTAGGCCACGG-3′ | |
| PRX12 | 5′-ATTGCTTCGTTCAGGGATGTG-3′ |
| 5′-TCGATCGCTTCCTGTCTCAA-3′ | |
| APX2 | 5′-GGTGCCACAAGGAGCGTTCAG-3′ |
| 5′-AAGAGCCTTGTCGGTTGGTAGTTG-3′ | |
| GPX | 5′-CAGCCTGAGGTTCGAGCATT-3′ |
| 5′-CACATCATTGCCACGAGCAT-3′ | |
| CAT2 | 5′-CCGGTGTTCAGACTCCTGTC-3′ |
| 5′-AAGAGCGTGGACCATGTCAG-3′ | |
| HSP22 | 5′-TGCTATCCGATCTCTGGCTAGACC-3′ |
| 5′-GGAGACAGAGCCACGCTTGTG-3′ | |
| HSP70.1 | 5′-TGTTGGACATTGACCTCTCTCT-3′ |
| 5′-CGTCATCGTAGCTAAACTGGT-3′ | |
| HSP70.2 | 5′-TCATTGGTGACCCCTTTCTCT-3′ |
| 5′-TCACATTTCTTCGAAGCTTTGTT-3′ | |
| HSP90.1 | 5′-TGGTTCTGAAAACTTCTAATATGTCG-3′ |
| 5′-TGACACAAACCCAACCCTAGA-3′ | |
| HSP90.2 | 5′-GGACTCACCGTGCTGTCTTGTAAC-3′ |
| 5′-ACTTGTCGTTCTTGTCTGCGTCAG-3′ | |
| HSP101 | 5′-AGGCAGGACAGTCGATTTCA-3′ |
| 5′-CCACAATCTCGTCAAGCCTG-3′ | |
| HSFA2 | 5′-ATCATGGTGTGCTTGTAGCTGAGG-3′ |
| 5′-AACGTCATCATCTGCTGCTGTCTC-3′ | |
| HSFA6B | 5′-ATCGAAGAGGCGATCAGCA-3′ |
| 5′-TGAGGATGAGGCTGCAACA-3′ | |
| GAPDH | 5′-CTCAATGACGGCCACACAGA-3′ |
| 5′-ACCAGTGCTGCTGGGAATG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtania, A.; Markiewicz, M.; Waligórski, P. Growth Cessation and Dormancy Induction in Micropropagated Plantlets of Rheum rhaponticum ‘Raspberry’ Influenced by Photoperiod and Temperature. Int. J. Mol. Sci. 2023, 24, 607. https://doi.org/10.3390/ijms24010607
Wojtania A, Markiewicz M, Waligórski P. Growth Cessation and Dormancy Induction in Micropropagated Plantlets of Rheum rhaponticum ‘Raspberry’ Influenced by Photoperiod and Temperature. International Journal of Molecular Sciences. 2023; 24(1):607. https://doi.org/10.3390/ijms24010607
Chicago/Turabian StyleWojtania, Agnieszka, Monika Markiewicz, and Piotr Waligórski. 2023. "Growth Cessation and Dormancy Induction in Micropropagated Plantlets of Rheum rhaponticum ‘Raspberry’ Influenced by Photoperiod and Temperature" International Journal of Molecular Sciences 24, no. 1: 607. https://doi.org/10.3390/ijms24010607