

Structure and Function of the Zinc Binding Protein ZrgA from Vibrio cholerae

Abstract
:1. Introduction
2. Results
2.1. ZrgA Is Essential to Zinc Import through zrgABCDE and Does Not Require the Flexible Loop
2.2. Expression, Purification and Zinc Binding of WT and Loop Deletion Mutants of ZrgA
2.3. Crystal Structure of V. Cholerae ZrgA Δ124-184
3. Discussion
4. Materials and Methods
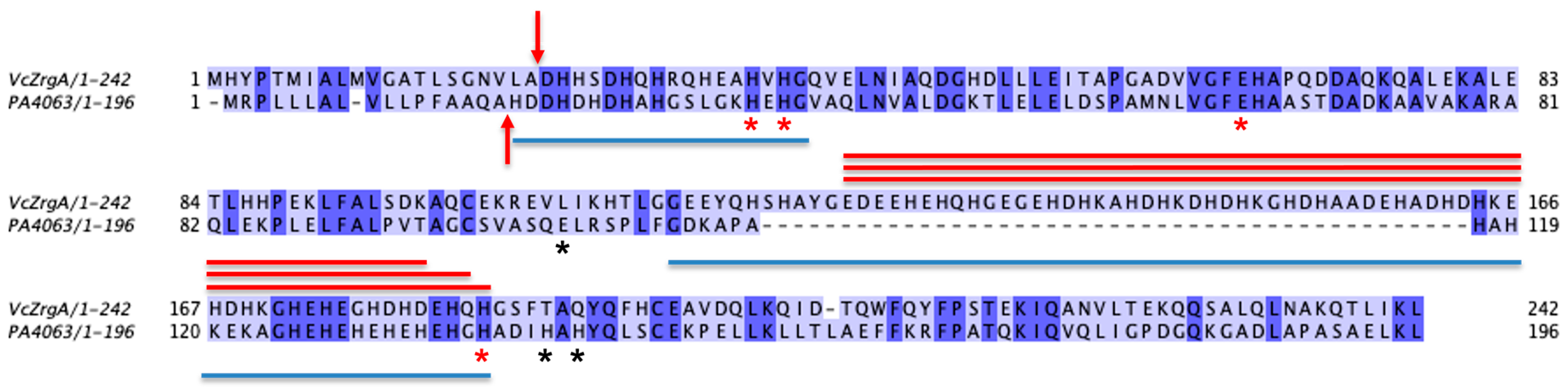
4.1. Multiple Sequence Alignments
4.2. Strains and Media
4.3. Construction of V. cholerae Deletion Mutant Strains
4.4. Cell Growth Determination
4.5. Colony Forming Units
4.6. Expression and Purification of ZrgA and Mutant Variants
4.7. Generation of Apo Proteins and Metal Quantitation
4.8. Mag-Fura 2 Competition Assay
4.9. Equilibrium Zinc Binding Assay
4.10. Crystallization and Structure Determination
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SBP | Substrate binding proteins |
| ABC | ATP binding cassette |
| CFU | Colony Forming Units |
| MF2 | Mag Fura-2 |
References
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.L.; Dassa, E.; Orelle, C.; Chen, J. Structure, function, and evolution of bacterial ATP-binding cassette systems. Microbiol. Mol. Biol. Rev. 2008, 72, 317–364, table of contents. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heide, T.; Poolman, B. ABC transporters: One, two or four extracytoplasmic substrate-binding sites? EMBO Rep. 2002, 3, 938–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khare, D.; Oldham, M.L.; Orelle, C.; Davidson, A.L.; Chen, J. Alternating access in maltose transporter mediated by rigid-body rotations. Mol. Cell 2009, 33, 528–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berntsson, R.P.; Smits, S.H.; Schmitt, L.; Slotboom, D.J.; Poolman, B. A structural classification of substrate-binding proteins. FEBS Lett. 2010, 584, 2606–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheepers, G.H.; Lycklama, A.N.J.A.; Poolman, B. An updated structural classification of substrate-binding proteins. FEBS Lett. 2016, 590, 4393–4401. [Google Scholar] [CrossRef] [PubMed]
- Kehl-Fie, T.E.; Skaar, E.P. Nutritional immunity beyond iron: A role for manganese and zinc. Curr. Opin. Chem. Biol. 2010, 14, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Yatsunyk, L.A.; Easton, J.A.; Kim, L.R.; Sugarbaker, S.A.; Bennett, B.; Breece, R.M.; Vorontsov, I.; Tierney, D.L.; Crowder, M.W.; Rosenzweig, A.C. Structure and metal binding properties of ZnuA, a periplasmic zinc transporter from Escherichia coli. J. Biol. Inorg. Chem. 2008, 13, 271–288. [Google Scholar] [CrossRef] [Green Version]
- Ilari, A.; Alaleona, F.; Tria, G.; Petrarca, P.; Battistoni, A.; Zamparelli, C.; Verzili, D.; Falconi, M.; Chiancone, E. The Salmonella enterica ZinT structure, zinc affinity and interaction with the high-affinity uptake protein ZnuA provide insight into the management of periplasmic zinc. Biochim. Biophys. Acta 2014, 1840, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Pederick, V.G.; Eijkelkamp, B.A.; Begg, S.L.; Ween, M.P.; McAllister, L.J.; Paton, J.C.; McDevitt, C.A. ZnuA and zinc homeostasis in Pseudomonas aeruginosa. Sci. Rep. 2015, 5, 13139. [Google Scholar] [CrossRef] [Green Version]
- Desrosiers, D.C.; Sun, Y.C.; Zaidi, A.A.; Eggers, C.H.; Cox, D.L.; Radolf, J.D. The general transition metal (Tro) and Zn2+ (Znu) transporters in Treponema pallidum: Analysis of metal specificities and expression profiles. Mol. Microbiol. 2007, 65, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Boyd, B.; Lingwood, C.A. Identification of the Key Protein for Zinc Uptake in Hemophilus influenzae. J. Biol. Chem. 1997, 272, 29033–29038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.; Randich, A.M.; Bhattacharyya-Pakrasi, M.; Pakrasi, H.B.; Smith, T.J. Possible regulatory role for the histidine-rich loop in the zinc transport protein, ZnuA. Biochemistry 2007, 46, 8734–8743. [Google Scholar] [CrossRef] [PubMed]
- Neupane, D.P.; Kumar, S.; Yukl, E.T. Two ABC Transporters and a Periplasmic Metallochaperone Participate in Zinc Acquisition in Paracoccus denitrificans. Biochemistry 2018, 58, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.; Fan, F.; Jensen, O.; Zhong, Z.; Kan, B.; Wang, H.; Zhu, J. Dual Zinc Transporter Systems in Vibrio cholerae Promote Competitive Advantages over Gut Microbiome. Infect. Immun. 2015, 83, 3902–3908. [Google Scholar] [CrossRef] [Green Version]
- Meléndez, A.B.; Valencia, D.; Yukl, E.T. Specificity of Interactions between Components of Two Zinc ABC Transporters in. Int. J. Mol. Sci. 2020, 21, 9098. [Google Scholar] [CrossRef]
- Fiorillo, A.; Battistoni, A.; Ammendola, S.; Secli, V.; Rinaldo, S.; Cutruzzolà, F.; Demitri, N.; Ilari, A. Structure and metal-binding properties of PA4063, a novel player in periplasmic zinc trafficking by Pseudomonas aeruginosa. Acta Cryst. D Struct. Biol. 2021, 77, 1401–1410. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Golynskiy, M.V.; Gunderson, W.A.; Hendrich, M.P.; Cohen, S.M. Metal binding studies and EPR spectroscopy of the manganese transport regulator MntR. Biochemistry 2006, 45, 15359–15372. [Google Scholar] [CrossRef]
- Abate, F.; Malito, E.; Cozzi, R.; Lo Surdo, P.; Maione, D.; Bottomley, M.J. Apo, Zn2+-bound and Mn2+-bound structures reveal ligand-binding properties of SitA from the pathogen Staphylococcus pseudintermedius. Biosci. Rep. 2014, 34, e00154. [Google Scholar] [CrossRef] [PubMed]
- Plumptre, C.D.; Eijkelkamp, B.A.; Morey, J.R.; Behr, F.; Counago, R.M.; Ogunniyi, A.D.; Kobe, B.; O′Mara, M.L.; Paton, J.C.; McDevitt, C.A. AdcA and AdcAII employ distinct zinc acquisition mechanisms and contribute additively to zinc homeostasis in Streptococcus pneumoniae. Mol. Microbiol. 2014, 91, 834–851. [Google Scholar] [CrossRef] [PubMed]
- Meni, A.; Yukl, E.T. Structural Features Mediating Zinc Binding and Transfer in the AztABCD Zinc Transporter System. Biomolecules 2020, 10, 1156. [Google Scholar] [CrossRef] [PubMed]
- Blindauer, C.A. Advances in the molecular understanding of biological zinc transport. Chem. Commun. 2015, 51, 4544–4563. [Google Scholar] [CrossRef] [Green Version]
- Udagedara, S.R.; La Porta, D.M.; Spehar, C.; Purohit, G.; Hein, M.J.A.; Fatmous, M.E.; Casas Garcia, G.P.; Ganio, K.; McDevitt, C.A.; Maher, M.J. Structural and functional characterizations of the C-terminal domains of CzcD proteins. J. Inorg. Biochem. 2020, 208, 111087. [Google Scholar] [CrossRef]
- Neville, S.L.; Sjöhamn, J.; Watts, J.A.; MacDermott-Opeskin, H.; Fairweather, S.J.; Ganio, K.; Carey Hulyer, A.; McGrath, A.P.; Hayes, A.J.; Malcolm, T.R.; et al. The structural basis of bacterial manganese import. Sci. Adv. 2021, 7, eabg3980. [Google Scholar] [CrossRef]
- Crow, A.; Greene, N.P.; Kaplan, E.; Koronakis, V. Structure and mechanotransmission mechanism of the MacB ABC transporter superfamily. Proc. Natl. Acad. Sci. USA 2017, 114, 12572–12577. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, H.; Kobayashi, H.; Takahashi, E.; Okamoto, K. MacAB is involved in the secretion of Escherichia coli heat-stable enterotoxin II. J. Bacteriol. 2008, 190, 7693–7698. [Google Scholar] [CrossRef] [Green Version]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef] [Green Version]
- Consortium, U. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, J.C.; Karplus, K.; Schoolnik, G.K.; Yildiz, F.H. Identification and characterization of RbmA, a novel protein required for the development of rugose colony morphology and biofilm structure in Vibrio cholerae. J. Bacteriol. 2006, 188, 1049–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Graichen, M.E.; Liu, A.; Pearson, A.R.; Wilmot, C.M.; Davidson, V.L. MauG, a novel diheme protein required for tryptophan tryptophylquinone biogenesis. Biochemistry 2003, 42, 7318–7325. [Google Scholar] [CrossRef]
- Handali, M.; Neupane, D.P.; Roychowdhury, H.; Yukl, E.T. Transcriptional Regulation, Metal Binding Properties and Structure of Pden1597, an Unusual Zinc Transport Protein from Paracoccus denitrificans. J. Biol. Chem. 2015, 290, 11878–11889. [Google Scholar] [CrossRef] [Green Version]
- Edelhoch, H. Spectroscopic determination of tryptophan and tyrosine in proteins. Biochemistry 1967, 6, 1948–1954. [Google Scholar] [CrossRef]
- Handali, M.; Roychowdhury, H.; Neupane, D.P.; Yukl, E.T. AztD, a Periplasmic Zinc Metallochaperone to an ATP-binding Cassette (ABC) Transporter System in Paracoccus denitrificans. J. Biol. Chem. 2015, 290, 29984–29992. [Google Scholar] [CrossRef] [Green Version]
- Kuzmic, P. Program DYNAFIT for the analysis of enzyme kinetic data: Application to HIV proteinase. Anal. Biochem. 1996, 237, 260–273. [Google Scholar] [CrossRef]
- Kuzmic, P. DynaFit—A software package for enzymology. Methods Enzym. 2009, 467, 247–280. [Google Scholar] [CrossRef]
- Luft, J.R.; Collins, R.J.; Fehrman, N.A.; Lauricella, A.M.; Veatch, C.K.; DeTitta, G.T. A deliberate approach to screening for initial crystallization conditions of biological macromolecules. J. Struct. Biol. 2003, 142, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Cryst. D Biol. Cryst. 2010, 66, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabsch, W. Xds. Acta Cryst. D Biol. Cryst. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Cryst. D Biol. Cryst. 2011, 67, 235–242. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Cryst. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Cryst. D Biol. Cryst. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. D Biol. Cryst. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Skubak, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Cryst. D Biol. Cryst. 2011, 67, 355–367. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Site | Kd ± S.D. (nM) |
|---|---|---|
| WT ZrgA | 1 | 0.1 ± 0.2 * |
| 2 | 5.7 ± 4.0 | |
| 3 | 35 ± 22 | |
| 4 | 287 ± 215 | |
| Δ124-184 ZrgA | 1 | 0.5 ± 0.3 * |
| 2 | 22 ± 3 | |
| 3 | 155 ± 66 | |
| Δ124-183 ZrgA | 1 | 0.8 ± 0.7 * |
| 2 | 148 ± 126 | |
| 3 | 237 ± 154 | |
| Δ124-180 ZrgA | 1 | 0.03 ± 0.06 * |
| 2 | 15 ± 4 | |
| 3 | 86 ± 21 | |
| 4 | 111 ± 53 |
| Δ124-184 ZrgA Apo | Δ124-184 ZrgA Zinc | Δ124-184 ZrgA Zinc, Zn-Edge | |
|---|---|---|---|
| Data collection | |||
| Wavelength (Å) | 1.000 | 1.000 | 1.281 |
| Space group | P21 | P21 | P21 |
| Unit cell parameters a, b, c (Å) α, β, γ (°) | 81.1, 65.4, 88.7 90, 98.8, 90 | 82.0, 66.9, 88.2 90, 99.9, 90 | 82.0, 66.9, 88.4 90, 99.8, 90 |
| Resolution range (Å) | 45.84–2.00 | 46.63–2.41 | 46.65–2.70 |
| Number of reflections (measured/unique) | 236,438/61,741 | 139,225/36,368 | 99,000/26,001 |
| Rmerge | 0.06 (0.64) | 0.10 (0.69) | 0.11 (0.69) |
| I/σI | 11.1 (2.2) | 9.3 (1.9) | 7.1 (1.9) |
| Completeness (%) | 99.2 (98.9) | 99.4 (99.8) | 99.3 (99.7) |
| CC1/2 | 1.00 (0.87) | 1.00 (0.80) | 0.99 (0.83) |
| Redundancy | 3.8 (3.8) | 3.8 (3.9) | 3.8 (3.9) |
| Refinement Statistics | |||
| Resolution (Å) | 45.88–2.00 | 46.67–2.41 | |
| Rwork/Rfree | 19.2/23.6 | 21.8/26.2 | |
| Number of atoms | |||
| Protein | 6381 | 6666 | |
| Zinc | 0 | 21 | |
| Water | 362 | 96 | |
| Other | 20 | 10 | |
| R.m.s. deviations | |||
| Bond lengths (Å) | 0.016 | 0.013 | |
| Bond angles (°) | 1.847 | 1.323 | |
| Ramachandran Statistics | |||
| Allowed | 99.2% | 98.5% | |
| Outliers | 0.8% | 1.5% | |
| Average B-factor (Å2) | 36.6 | 40.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valencia, D.W.; Meléndez, A.B.; Melendrez, I.A.; Yukl, E.T. Structure and Function of the Zinc Binding Protein ZrgA from Vibrio cholerae. Int. J. Mol. Sci. 2023, 24, 548. https://doi.org/10.3390/ijms24010548
Valencia DW, Meléndez AB, Melendrez IA, Yukl ET. Structure and Function of the Zinc Binding Protein ZrgA from Vibrio cholerae. International Journal of Molecular Sciences. 2023; 24(1):548. https://doi.org/10.3390/ijms24010548
Chicago/Turabian StyleValencia, Daniel W., Ady B. Meléndez, Isaac A. Melendrez, and Erik T. Yukl. 2023. "Structure and Function of the Zinc Binding Protein ZrgA from Vibrio cholerae" International Journal of Molecular Sciences 24, no. 1: 548. https://doi.org/10.3390/ijms24010548