

Overexpression of ZmDHN15 Enhances Cold Tolerance in Yeast and Arabidopsis
 , and
, and 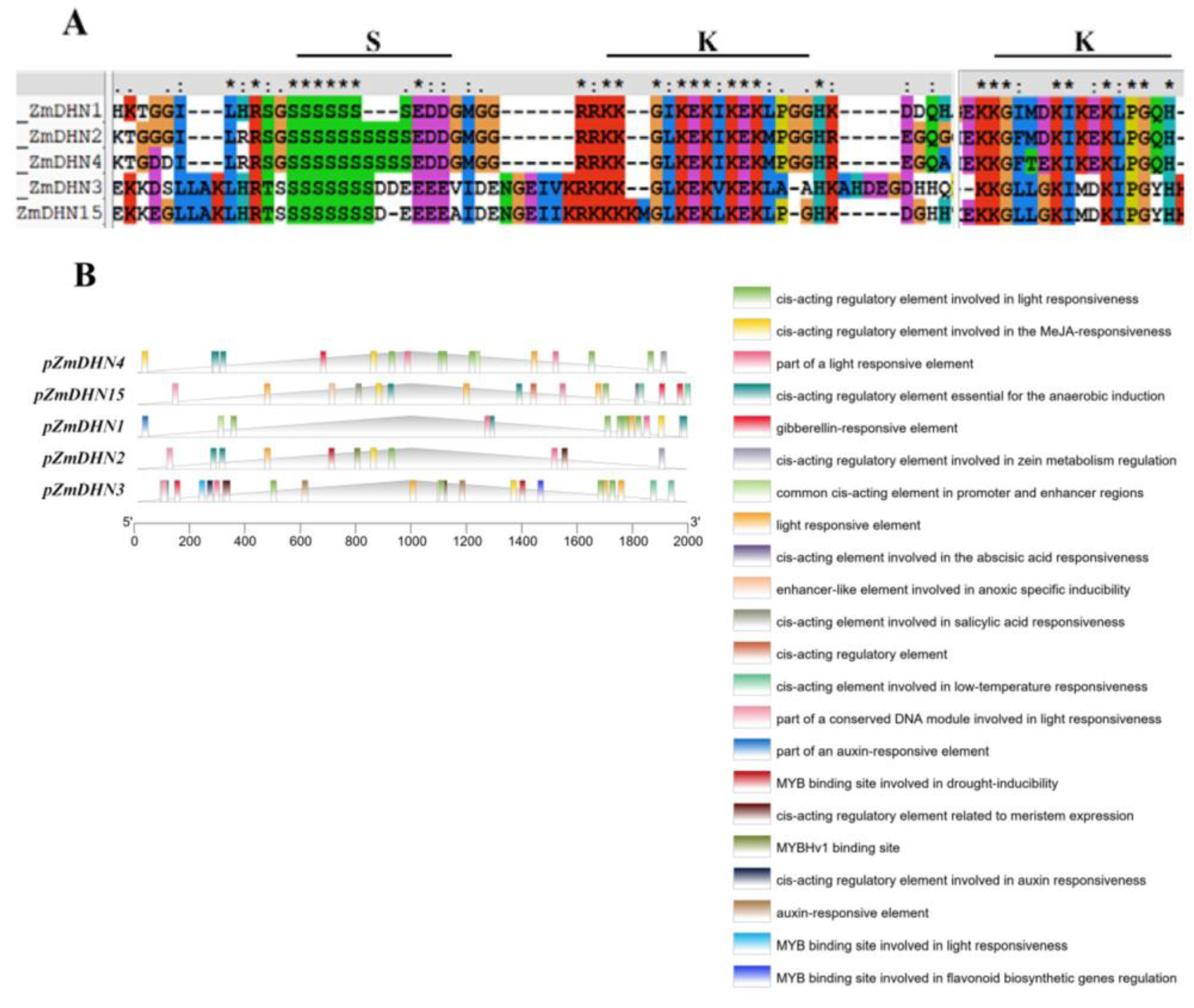{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bioinformatics Analysis of Maize Dehydrin Gene
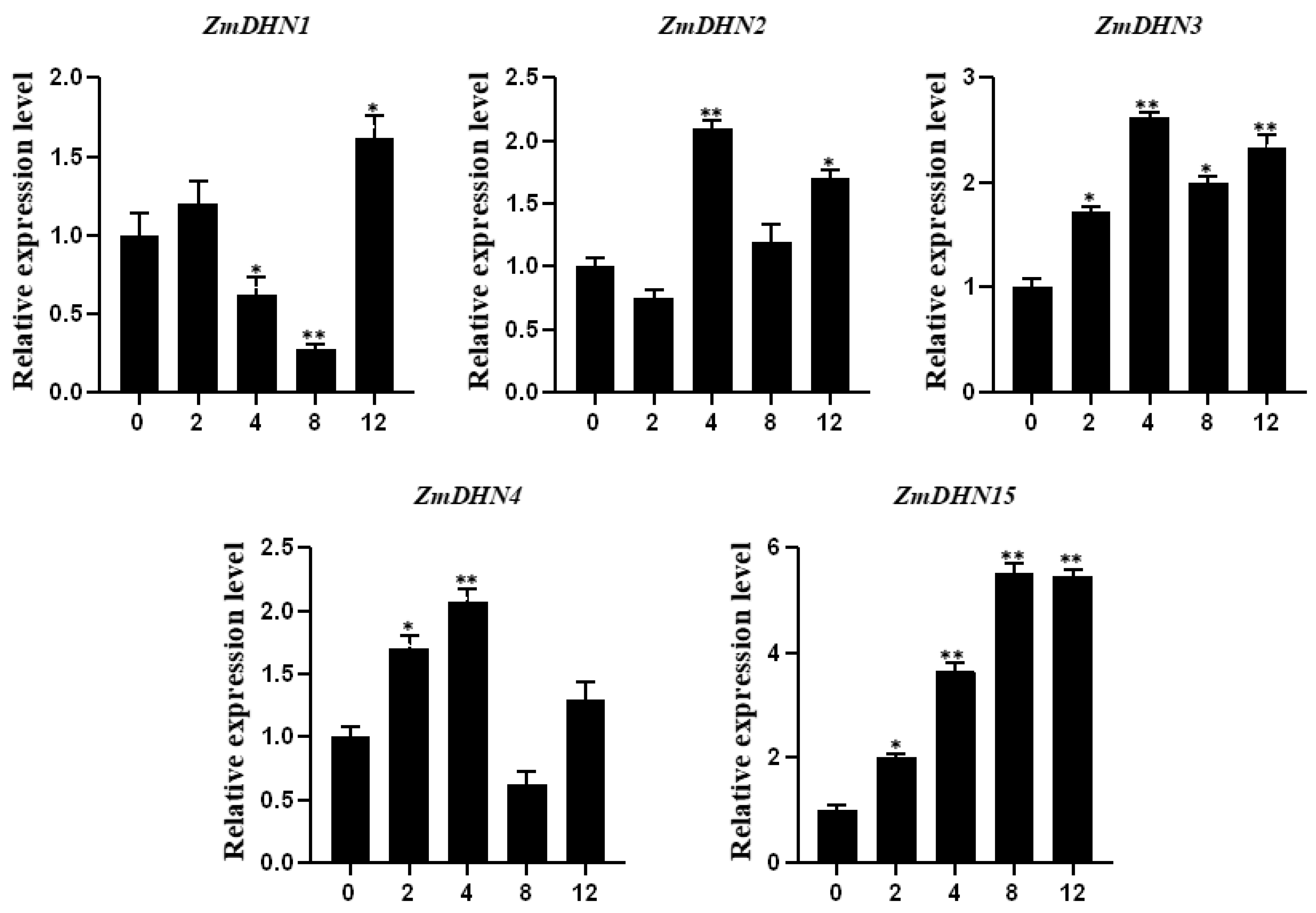
2.2. Analysis of the Expression Pattern of the Maize Dehydrin Gene under Cold Stress
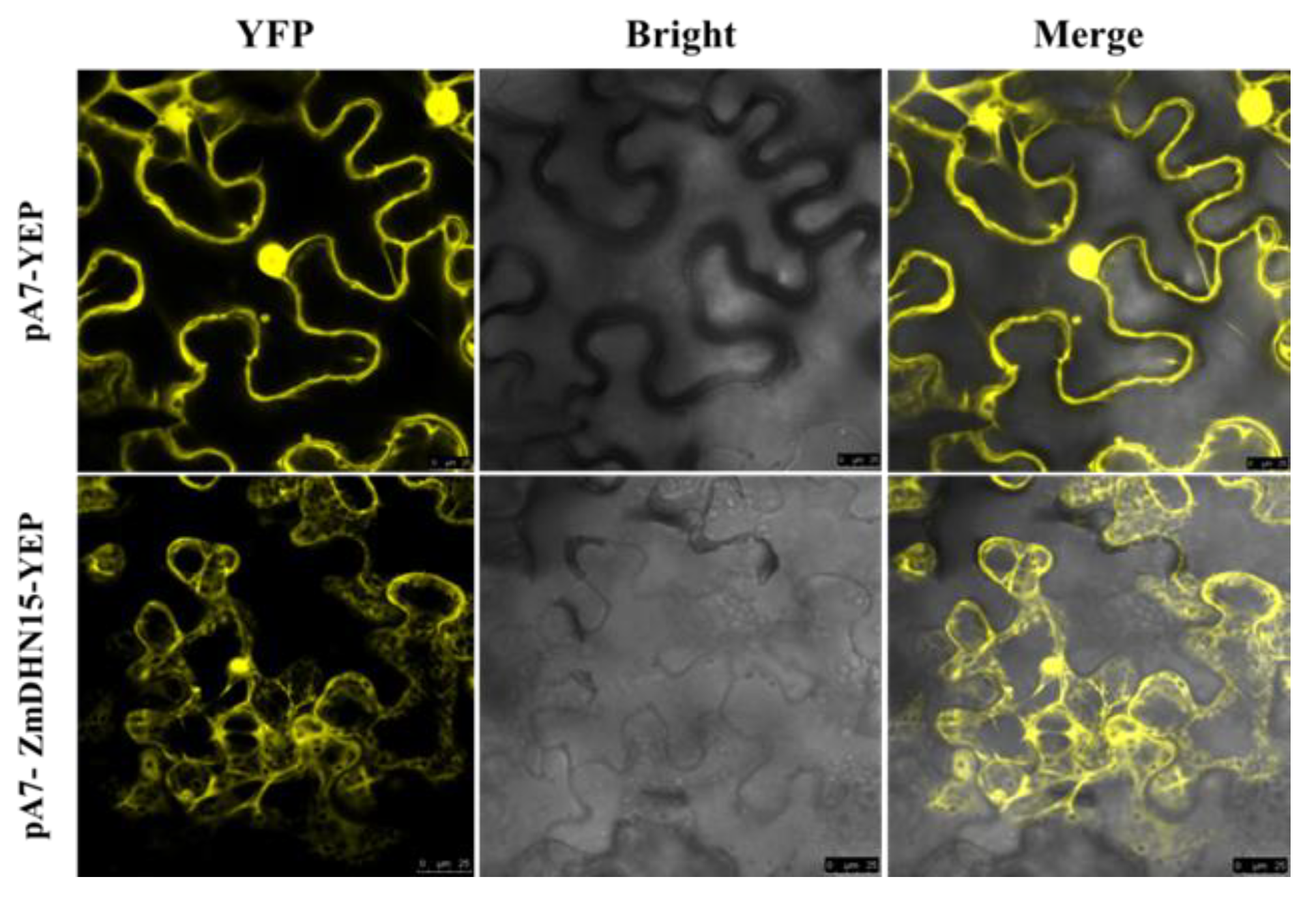
2.3. Subcellular Localization of ZmDHN15 Protein
2.4. Low-Temperature-Tolerance Assays of Yeast Transformants
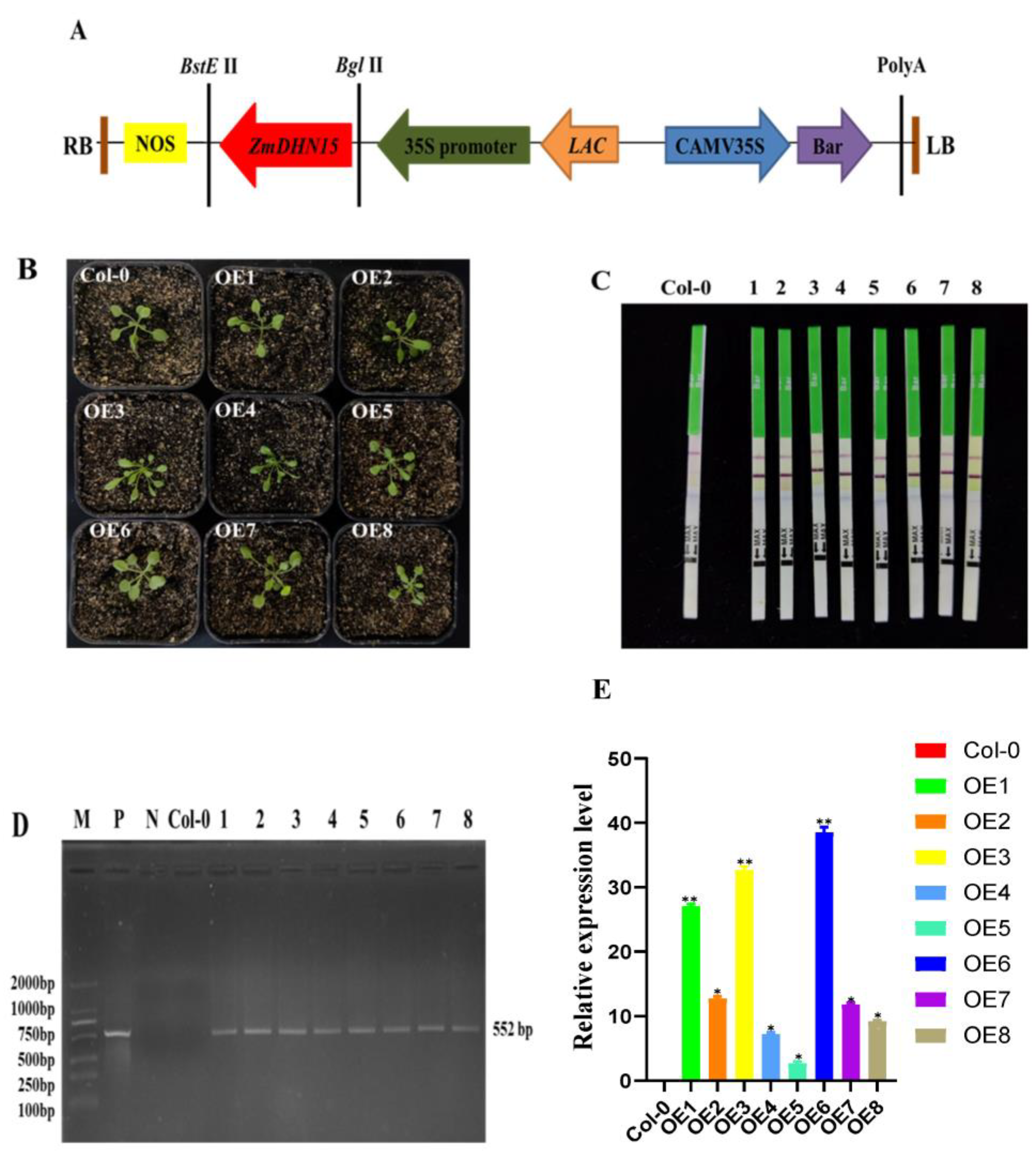
2.5. Generation of Transgenic Plants and Molecular Identification
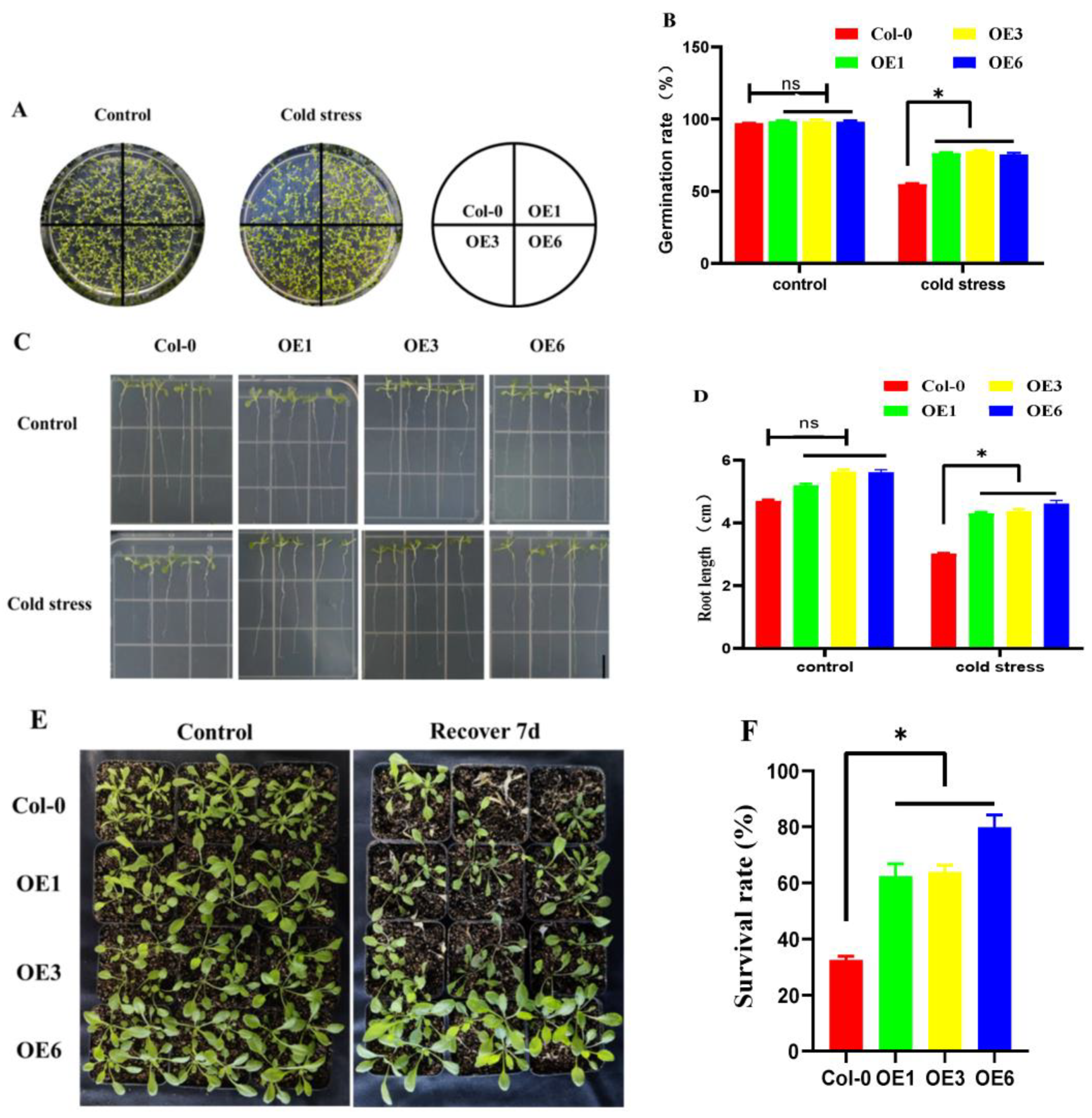
2.6. Overexpression of ZmDHN15 Enhances Cold Resistance in Transgenic Arabidopsis
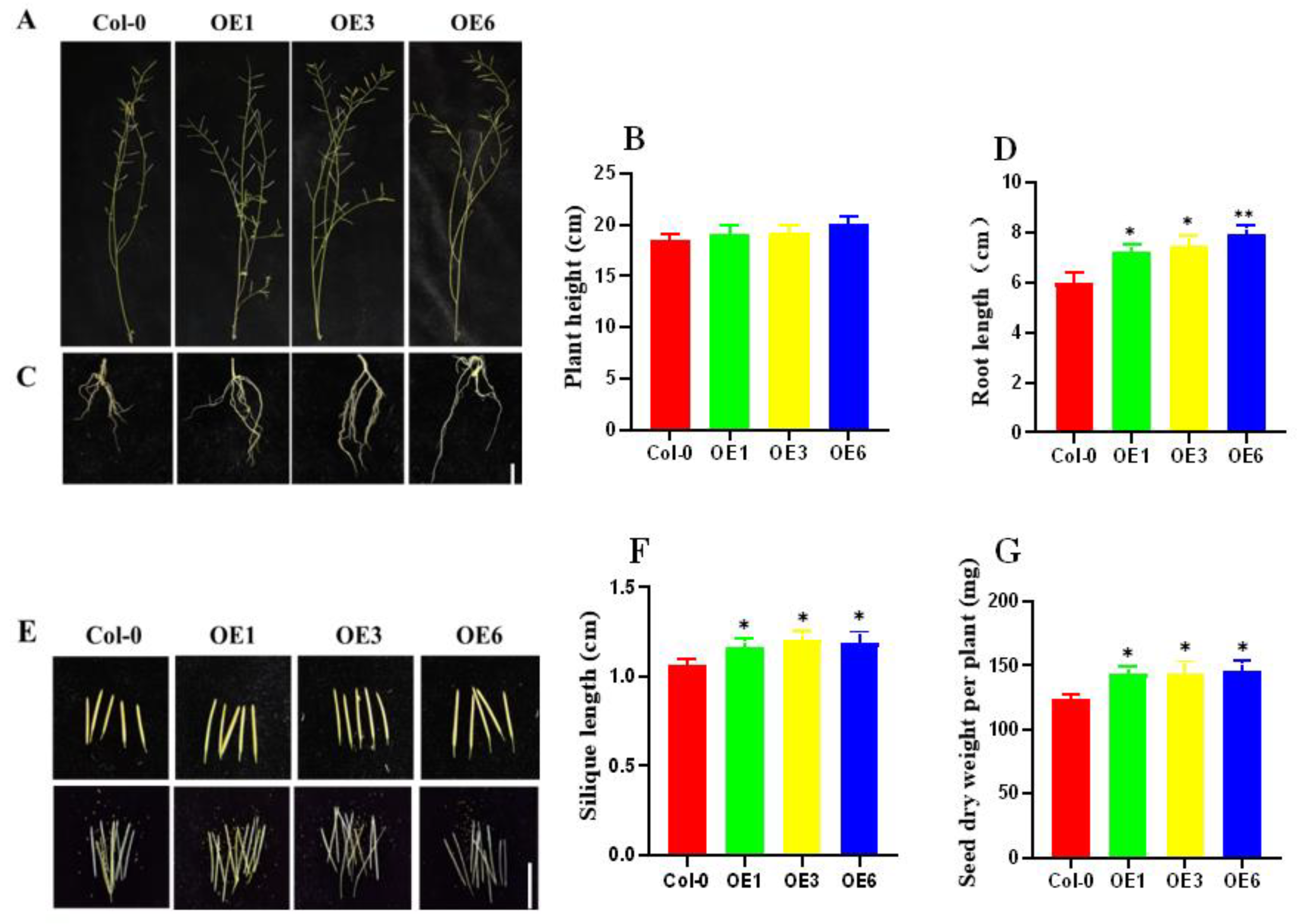
2.7. Phenotypic Characterization of Overexpressed ZmDHN15 at the Mature Stage
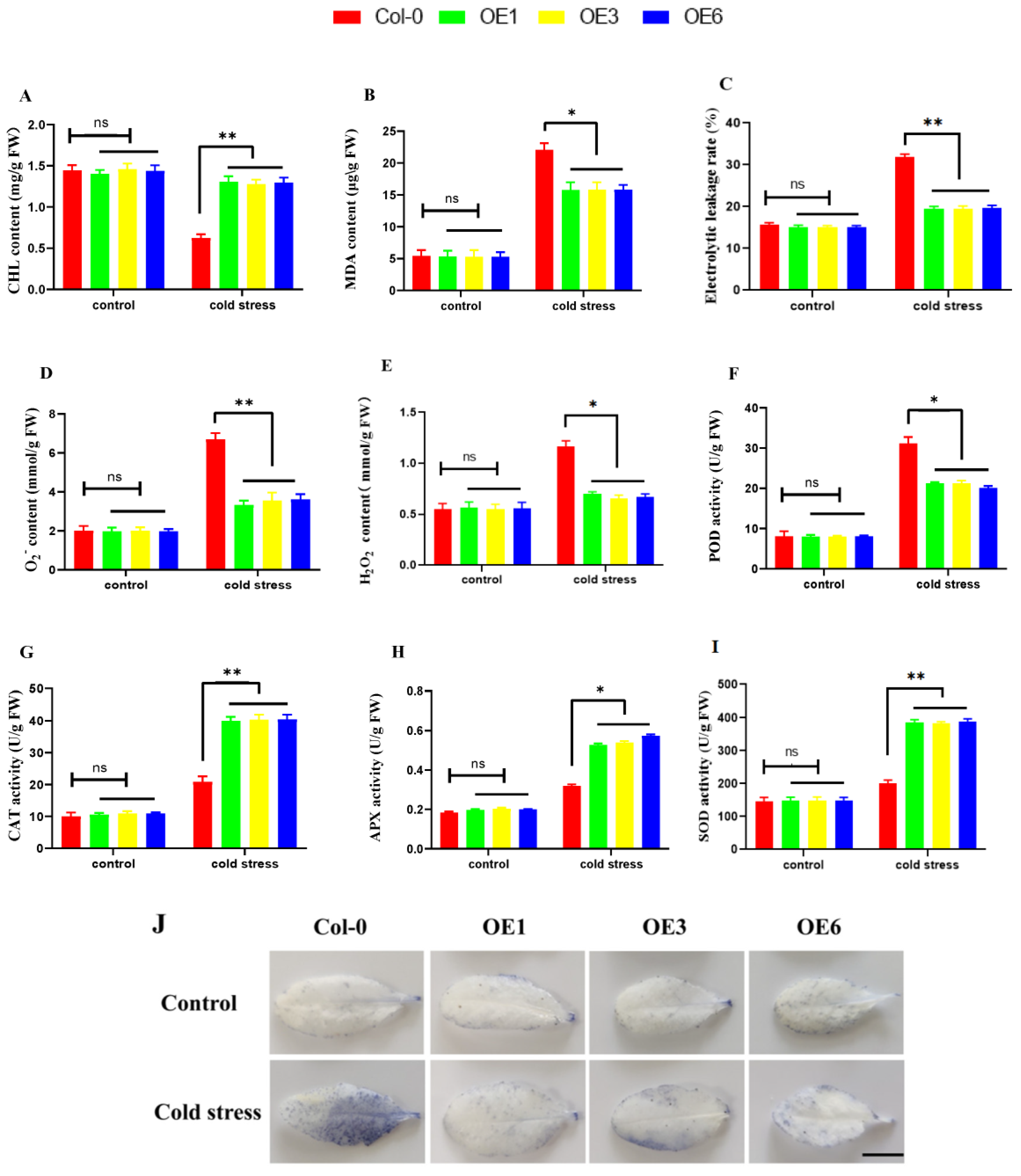
2.8. Overexpression of ZmDHN15 Reduces the ROS Accumulation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Screening of Maize Dehydrin Gene under Cold Stress
4.3. Subcellular Localization of ZmDHN15
4.4. Expression of ZmDHN15 in Saccharomyces Cerevisiae INVSc1
4.5. Low-Temperature Tolerance Assay of Yeast Transformants
4.6. Generation of Transgenic Plants and Phenotypic Analysis
4.7. Determination of Chlorophyll, Malondialdehyde, H2O2, and O2− Content and Antioxidative Enzyme Activity
4.8. NBT (Nitro-Blue Tetrazolium Chloride) Staining Assay
4.9. Analysis of Expression Patterns of ROS-Related Marker Genes and Cold-Responsive Genes in Transgenic Plants
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiao, P.; Jiang, Z.; Wei, X.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Overexpression of the homeobox-leucine zipper protein ATHB-6 improves the drought tolerance of maize (Zea mays L.). Plant Sci. 2022, 316, 111159. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Jin, S.; Chen, N.; Wang, C.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Improvement of cold tolerance in maize (Zea mays L.) using Agrobacterium-mediated transformation of ZmSAMDC gene. GM Crops Food 2022, 13, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Tan, J.; Feng, H.; Zhou, Z.; Deng, M.; Luo, H.; Deng, Z. Integrative Proteome and Phosphoproteome Profiling of Early Cold Response in Maize Seedlings. Int. J. Mol. Sci. 2022, 23, 6493. [Google Scholar] [CrossRef] [PubMed]
- Bilska-Kos, A.; Solecka, D.; Dziewulska, A.; Ochodzki, P.; Jonczyk, M.; Bilski, H.; Sowinski, P. Low temperature caused modifications in the arrangement of cell wall pectins due to changes of osmotic potential of cells of maize leaves (Zea mays L.). Protoplasma 2017, 254, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Xu, J.; Gao, Y.; Wang, C.; Guo, G.; Luo, Y.; Huang, Y.; Hu, W.; Sheteiwy, M.S.; Guan, Y.; et al. The Synergistic Priming Effect of Exogenous Salicylic Acid and H2O2 on Chilling Tolerance Enhancement during Maize (Zea mays L.) Seed Germination. Front. Plant Sci. 2017, 8, 1153. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Lin, L.; Zhang, Y.; Sui, N. ZmMYB31, a R2R3-MYB transcription factor in maize, positively regulates the expression of CBF genes and enhances resistance to chilling and oxidative stress. Mol. Biol. Rep. 2019, 46, 3937–3944. [Google Scholar] [CrossRef]
- Li, X.; Li, L.; Zuo, S.; Li, J.; Wei, S. Differentially expressed ZmASR genes associated with chilling tolerance in maize (Zea mays) varieties. Funct. Plant Biol. 2018, 45, 1173–1180. [Google Scholar] [CrossRef]
- Zhao, P.; Liu, F.; Ma, M.; Gong, J.; Wang, Q.; Jia, P.; Zheng, G.; Liu, H. Overexpression of AtLEA3-3 confers resistance to cold stress in Escherichia coli and provides enhanced osmotic stress tolerance and ABA sensitivity in Arabidopsis thaliana. Mol. Biol. 2011, 45, 785. [Google Scholar] [CrossRef]
- Xiong, H.; Yu, J.; Miao, J.; Li, J.; Zhang, H.; Xin, W.; Liu, P.; Yan, Z.; Jiang, C.; Yin, Z. Natural Variation in OsLG3 Increases Drought Tolerance in Rice by Inducing ROS Scavenging. Plant Physiol. 2018, 178, 451–467. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, X.; Liu, F.; Pan, D.; Ai, X. Hydrogen sulfide interacting with abscisic acid counteracts oxidative damages against chilling stress in cucumber seedlings. Acta Hortic. Sin. 2018, 45, 2395–2406. [Google Scholar]
- Ma; Xiaocui; Chen; Chong; Yang; Minmin; Dong; Xinchun; Lv; Wei Cold-regulated protein (SlCOR413IM1) confers chilling stress tolerance in tomato plants. Plant Physiol. Biochem. 2018, 124, 29–39. [CrossRef] [PubMed]
- Shi, H.; He, X.; Zhao, Y.; Lu, S.; Guo, Z. Constitutive expression of a group 3 LEA protein from Medicago falcata (MfLEA3) increases cold and drought tolerance in transgenic tobacco. Plant Cell Rep. 2020, 39, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, J.; Sun, L.; Yang, X.; Li, D. Group 3 LEA Protein, ZmLEA3, Is Involved in Protection from Low Temperature Stress. Front. Plant Sci. 2016, 7, 1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galau, G.A.; Dure, L., III. Developmental Biochemistry of Cottonseed Embryogenesis and Germination:Changing Messenger Ribonucleic Acid Populations As Shown by Reciprocal Heterologous Complementary Deoxyribonucleic Acid-Messenger Ribonucleic Acid Hybridization. Biochemistry 1981, 20, 4162–4168. [Google Scholar]
- Lei, Z.; Akinori, O.; Masamichi, T.; Ryozo, I. Expression of Plant Group 2 and Group 3 lea Genes in Saccharomyces cerevisiae Revealed Functional Divergence among LEA Proteins. J. Biochem. 2000, 127, 611–616. [Google Scholar]
- Ali, M.; Gul, A.; Hasan, H.; Alipour, H.; Abbasi, A.A.; Khan, F.; Abbas, S.; Fatima, T.; Taimoor, Z. LEA proteins and drought stress in wheat. Clim. Chang. Food Secur. Emphas. Wheat 2020, 12, 193–205. [Google Scholar]
- Kobayashi, F.; Ishibashi, M.; Takumi, S. Transcriptional activation of Cor/Lea genes and increase in abiotic stress tolerance through expression of a wheat DREB2 homolog in transgenic tobacco. Transgenic Res. 2008, 17, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, S.K.; Kutzer, M.; Procek, J.; Grbner, G.; Harryson, P. Tunable Membrane Binding of the Intrinsically Disordered Dehydrin Lti30, a Cold-Induced Plant Stress Protein. Plant Cell 2011, 23, 2391–2404. [Google Scholar] [CrossRef] [Green Version]
- Falavigna, V.; Malabarba, J.; Silveira, C.P.; Buffon, V.; Mariath, J.; Pasquali, G.; Margis-Pinheiro, M.; Revers, L.F. Characterization of the nucellus-specific dehydrin MdoDHN11 demonstrates its involvement in the tolerance to water deficit. Plant Cell Rep. 2019, 38, 1099–1107. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, D.; Lu, X.; Zhang, L.; Zhang, H. Characterisation of an SKn-type Dehydrin Promoter from Wheat and Its Responsiveness to Various Abiotic and Biotic Stresses. Plant Mol. Biol. Rep. 2014, 32, 664–678. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, X.; Lv, Q.; Zhu, D.; Qiu, T.; Xu, Y.; Bao, F.; He, Y.; Hu, Y. Physcomitrella Patens Dehydrins (PpDHNA and PpDHNC) Confer Salinity and Drought Tolerance to Transgenic Arabidopsis Plants. Front. Plant Sci. 2017, 8, 1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal. Behav. 2014, 6, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Du, D.; An, Y.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Overexpression of Prunus mume Dehydrin Genes in Tobacco Enhances Tolerance to Cold and Drought. Front. Plant Sci. 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Zhang, L.; Hui, L.; Li, H.; Zhang, Y.; Xu, Y.; Yu, J. The K-segments of wheat dehydrin WZY2 are essential for its protective functions under temperature stress. Front. Plant Sci. 2015, 6, 406. [Google Scholar] [CrossRef]
- Zhao, X.; Zhan, L.; Zou, X. Improvement of cold tolerance of the half-high bush Northland blueberry by transformation with the LEA gene from Tamarix androssowii. Plant Growth Regul. 2011, 63, 13–22. [Google Scholar] [CrossRef]
- Yokoyama, T.; Ohkubo, T.; Kamiya, K.; Hara, M. Cryoprotective activity of Arabidopsis KS-type dehydrin depends on the hydrophobic amino acids of two active segments. Arch. Biochem. Biophys. 2020, 691, 108510. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, F.; Wang, X.; Liu, S.; Saeed, U.H.; Hou, X.; Zhang, Y.; Luo, D.; Meng, Y.; Zhang, W.; et al. Molecular and Functional Characterization of CaNAC035, an NAC Transcription Factor From Pepper (Capsicum annuum L.). Front. Plant Sci. 2020, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, H.; Wang, A.; Zhang, L. An ERF-type transcription factor is involved in the regulation of the dehydrin wzy1-2 gene in wheat. Plant Signal. Behav. 2020, 15, 1778920. [Google Scholar] [CrossRef]
- Gao, T.; Mo, Y.; Huang, H.; Yu, J.; Wang, Y.; Wang, W. Heterologous Expression of Camellia sinensis Late Embryogenesis Abundant Protein Gene 1 (CsLEA1) Confers Cold Stress Tolerance in Escherichia coli and Yeast. Hortic. Plant J. 2021, 7, 89–96. [Google Scholar] [CrossRef]
- Shibuya, T.; Itai, R.; Maeda, M.; Kitashiba, H.; Isuzugawa, K.; Kato, K.; Kanayama, Y. Characterization of PcLEA14, a Group 5 Late Embryogenesis Abundant Protein Gene from Pear (Pyrus communis). Plants 2020, 9, 1138. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, H.; Pan, X.; Chen, N.; Hu, H.; Haq, S.U.; Khan, A.; Chen, R. CaDHN3, a Pepper (Capsicum annuum L.) Dehydrin Gene Enhances the Tolerance against Salt and Drought Stresses by Reducing ROS Accumulation. Int. J. Mol. Sci. 2021, 22, 3205. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Xia, W.; Hou, M.; Ruan, N.; Li, J.; Zhu, J. Cloning and function analysis of a Saussurea involucrata LEA4 gene. Front. Plant Sci. 2022, 13, 957133. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, D.; Song, Q.; Zhang, T.; Li, D.; Yang, X. The maize late embryogenesis abundant protein ZmDHN13 positively regulates copper tolerance in transgenic yeast and tobacco. Crop. J. 2019, 7, 403–410. [Google Scholar] [CrossRef]
- Xing, X.; Liu, Y.; Kong, X.; Liu, Y.; Li, D. Overexpression of a maize dehydrin gene, ZmDHN2b, in tobacco enhances tolerance to low temperature. Plant Growth Regul. 2011, 65, 109–118. [Google Scholar] [CrossRef]
- Ju, H.; Li, D.; Li, D.; Yang, X.; Liu, Y. Overexpression of ZmDHN11 could enhance transgenic yeast and tobacco tolerance to osmotic stress. Plant Cell Rep. 2021, 40, 1723–1733. [Google Scholar] [CrossRef]
- Hughes, S.L.; Schart, V.; Malcolmson, J.; Hogarth, K.A.; Martynowicz, D.M.; Tralman-Baker, E.; Patel, S.N.; Graether, S.P. The Importance of Size and Disorder in the Cryoprotective Effects of Dehydrins. Plant Physiol. 2013, 163, 1376–1386. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Tong, Q.; Wang, Y.; Wang, Z.; Xu, G.; Elias, G.K.; Li, S.; Liang, Z. Transcriptomic Analysis of the Grapevine LEA Gene Family in Response to Osmotic and Cold Stress Reveals a Key Role for VamDHN3. Plant Cell Physiol. 2020, 61, 775–786. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Zhang, T.; Yang, X.; Li, D. Functional characterization of KS-type dehydrin ZmDHN13 and its related conserved domains under oxidative stress. Sci. Rep. 2017, 7, 7361. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.F.; Liu, S.Y.; Ma, J.H.; Wang, X.K.; Haq, S.U.; Meng, Y.C.; Zhang, Y.M.; Chen, R.G. CaDHN4, a Salt and Cold Stress-Responsive Dehydrin Gene from Pepper Decreases Abscisic Acid Sensitivity in Arabidopsis. Int. J. Mol. Sci. 2019, 21, 26. [Google Scholar] [CrossRef] [Green Version]
- Lv, A.; Fan, N.; Xie, J.; Yuan, S.; An, Y.; Zhou, P. Expression of CdDHN4, a Novel YSK2-Type Dehydrin Gene from Bermudagrass, Responses to Drought Stress through the ABA-Dependent Signal Pathway. Front. Plant Sci. 2017, 8, 748. [Google Scholar] [CrossRef]
- Richard, S.; Morency, M.J.; Drevet, C.; Jouanin, L.; Séguin, A. Isolation and characterization of a dehydrin gene from white spruce induced upon wounding, drought and cold stresses. Plant Mol. Biol. 2000, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Terashima, S.; Fukaya, T.; Kuboi, T. Enhancement of cold tolerance and inhibition of lipid peroxidation by citrus dehydrin in transgenic tobacco. Planta 2003, 217, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Shah Jahan, M.; Wang, Y.; Shu, S.; Zhong, M.; Chen, Z.; Wu, J.; Sun, J.; Guo, S. Exogenous salicylic acid increases the heat tolerance in Tomato (Solanum lycopersicum L.) by enhancing photosynthesis efficiency and improving antioxidant defense system through scavenging of reactive oxygen species. Sci. Hortic.-Amst. 2019, 247, 421–429. [Google Scholar] [CrossRef]
- Yang, L.; Fountain, J.C.; Wang, H.; Ni, X.; Ji, P.; Lee, R.D.; Kemerait, R.C.; Scully, B.T.; Guo, B. Stress Sensitivity Is Associated with Differential Accumulation of Reactive Oxygen and Nitrogen Species in Maize Genotypes with Contrasting Levels of Drought Tolerance. Int. J. Mol. Sci. 2015, 16, 24791–24819. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, Y.; Qian, Y.; Chan, Z. Low Temperature-Induced 30 (LTI30) positively regulates drought stress resistance in Arabidopsis: Effect on abscisic acid sensitivity and hydrogen peroxide accumulation. Front. Plant Sci. 2015, 6, 893. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Cao, L.; Zhang, X.; Zhang, W.; Yang, T.; Zhang, J.; Che, D. An R2R3-MYB Transcription Factor RmMYB108 Responds to Chilling Stress of Rosa multiflora and Conferred Cold Tolerance of Arabidopsis. Front. Plant Sci. 2021, 12, 696919. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rasheed, R.; Hussain, I.; Iqbal, M.; Haider, M.Z.; Parveen, S.; Sajid, M.A. Hydrogen peroxide modulates antioxidant system and nutrient relation in maize (Zea mays L.) under water-deficit conditions. Arch. Agron. Soil Sci. 2014, 61, 507–523. [Google Scholar] [CrossRef]
- Novillo, F.; Medina, J.; Salinas, J. Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon. Proc. Natl. Acad. Sci. USA 2007, 104, 21002–21007. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dang, P.; Liu, L.; He, C. Cold acclimation by the CBF–COR pathway in a changing climate: Lessons from Arabidopsis thaliana. Plant Cell Rep. 2019, 38, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.N.; Tossounian, M.; Kovacs, D.S.; Thu, T.T.; Stijlemans, B.; Vertommen, D.; Pauwels, J.; Gevaert, K.; Angenon, G.; Messens, J.; et al. Dehydrin ERD14 activates glutathione transferase Phi9 in Arabidopsis thaliana under osmotic stress. Biochim. Et Biophys. Acta (BBA)—Gen. Subj. 2020, 1864, 129506. [Google Scholar] [CrossRef]
- Lee, S.C.; Kim, S.H.; Kim, S.R. Drought inducible OsDhn1 promoter is activated by OsDREB1A and OsDREB1D. J. Plant Biol. 2013, 56, 115–121. [Google Scholar] [CrossRef]
- Kamisugi, Y.; Cuming, A.C. The Evolution of the Abscisic Acid-response in Land Plants: Comparative Analysis of Group 1 LEA Gene Expression in Moss and Cereals. Plant Mol. Biol. 2005, 59, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Klima, M.; Vitamvas, P.; Zelenkova, S.; Vyvadilova, M.; Prasil, I.T. Dehydrin and proline content in Brassica napus and B. carinata under cold stress at two irradiances. Biol. Plant. 2012, 56, 157–161. [Google Scholar] [CrossRef]
- Xiong, J.; Zhang, W.; Zheng, D.; Xiong, H.; Feng, X.; Zhang, X.; Wang, Q.; Wu, F.; Xu, J.; Lu, Y. ZmLBD5 Increases Drought Sensitivity by Suppressing ROS Accumulation in Arabidopsis. Plants 2022, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Kjg, A.; Gm, B.; Sgk, C.; Ah, D.; Fs, E. Short-term cold stress affects physiological and biochemical traits of pistachio rootstocks. S. Afr. J. Bot. 2021, 141, 90–98. [Google Scholar]
- Yu, Y.; Li, Y.; Jia, F.; Zhao, M.; Li, W.; Sun, Q.; Li, N.; Li, W.; Meng, Z. ZmFKBP20-1 improves the drought and salt tolerance of transformed Arabidopsis. J. Plant Biol. 2017, 60, 558–570. [Google Scholar] [CrossRef]
- Sohag, A.; Tahjib-Ul-Arif, M.; Brestic, M.; Afrin, S.; Hossain, M.A. Exogenous salicylic acid and hydrogen peroxide attenuate drought stress in rice. Plant Soil Environ. 2020, 66, 7–13. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, N.; Fan, X.; Wang, C.; Jiao, P.; Jiang, Z.; Ma, Y.; Guan, S.; Liu, S. Overexpression of ZmDHN15 Enhances Cold Tolerance in Yeast and Arabidopsis. Int. J. Mol. Sci. 2023, 24, 480. https://doi.org/10.3390/ijms24010480
Chen N, Fan X, Wang C, Jiao P, Jiang Z, Ma Y, Guan S, Liu S. Overexpression of ZmDHN15 Enhances Cold Tolerance in Yeast and Arabidopsis. International Journal of Molecular Sciences. 2023; 24(1):480. https://doi.org/10.3390/ijms24010480
Chicago/Turabian StyleChen, Nannan, Xuhong Fan, Chunlai Wang, Peng Jiao, Zhenzhong Jiang, Yiyong Ma, Shuyan Guan, and Siyan Liu. 2023. "Overexpression of ZmDHN15 Enhances Cold Tolerance in Yeast and Arabidopsis" International Journal of Molecular Sciences 24, no. 1: 480. https://doi.org/10.3390/ijms24010480