
The Update Immune-Regulatory Role of Pro- and Anti-Inflammatory Cytokines in Recurrent Pregnancy Losses

Abstract
:1. Introduction
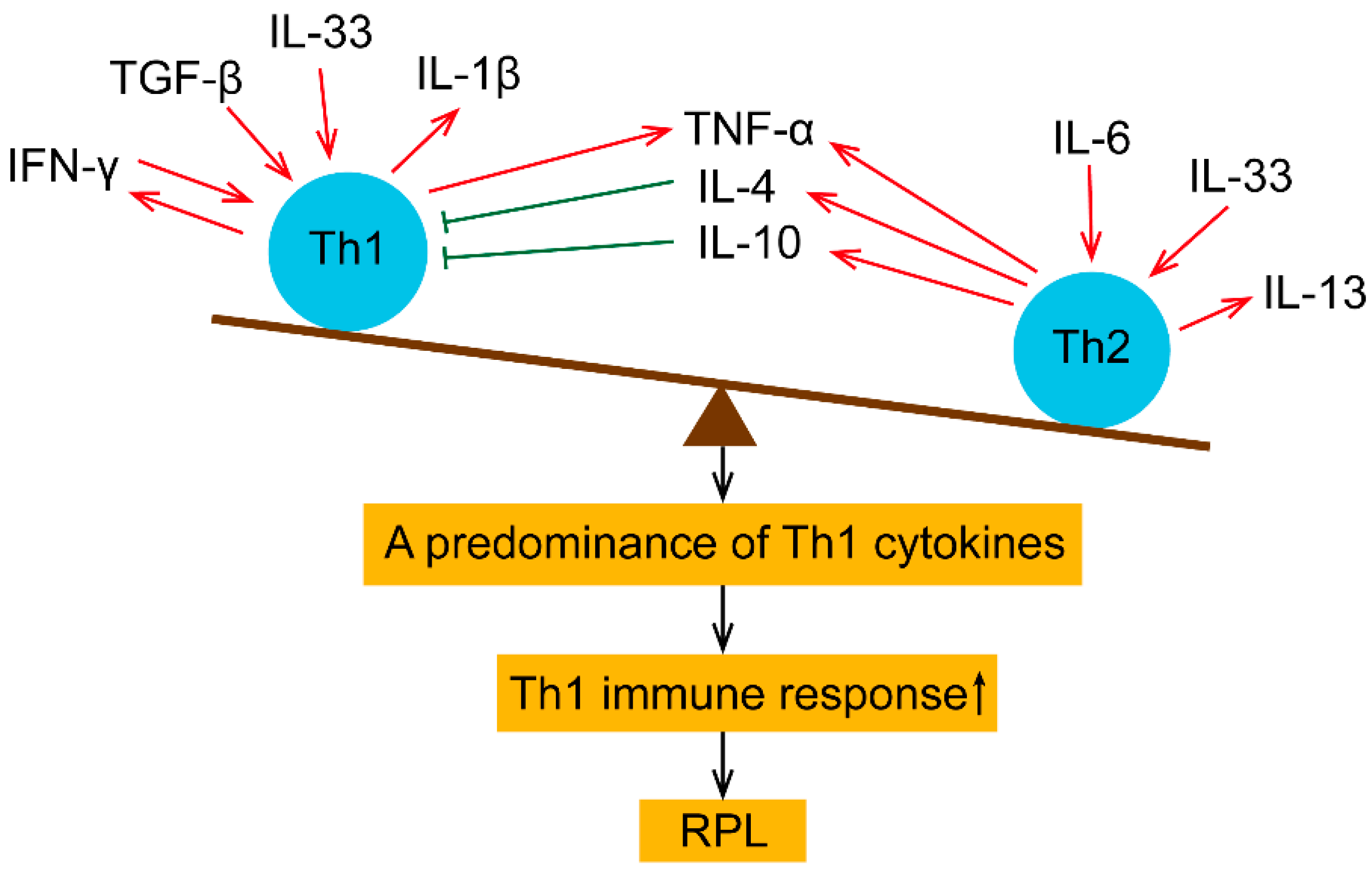
2. Pro- and Anti-Inflammatory Cytokines in Maternal Peripheral Blood
2.1. TNF-α
2.2. IFN-γ
2.3. Transforming Growth Factor (TGF)-β
2.4. IL-1β
2.5. IL-33
2.6. IL-4
2.7. IL-6
2.8. IL-10
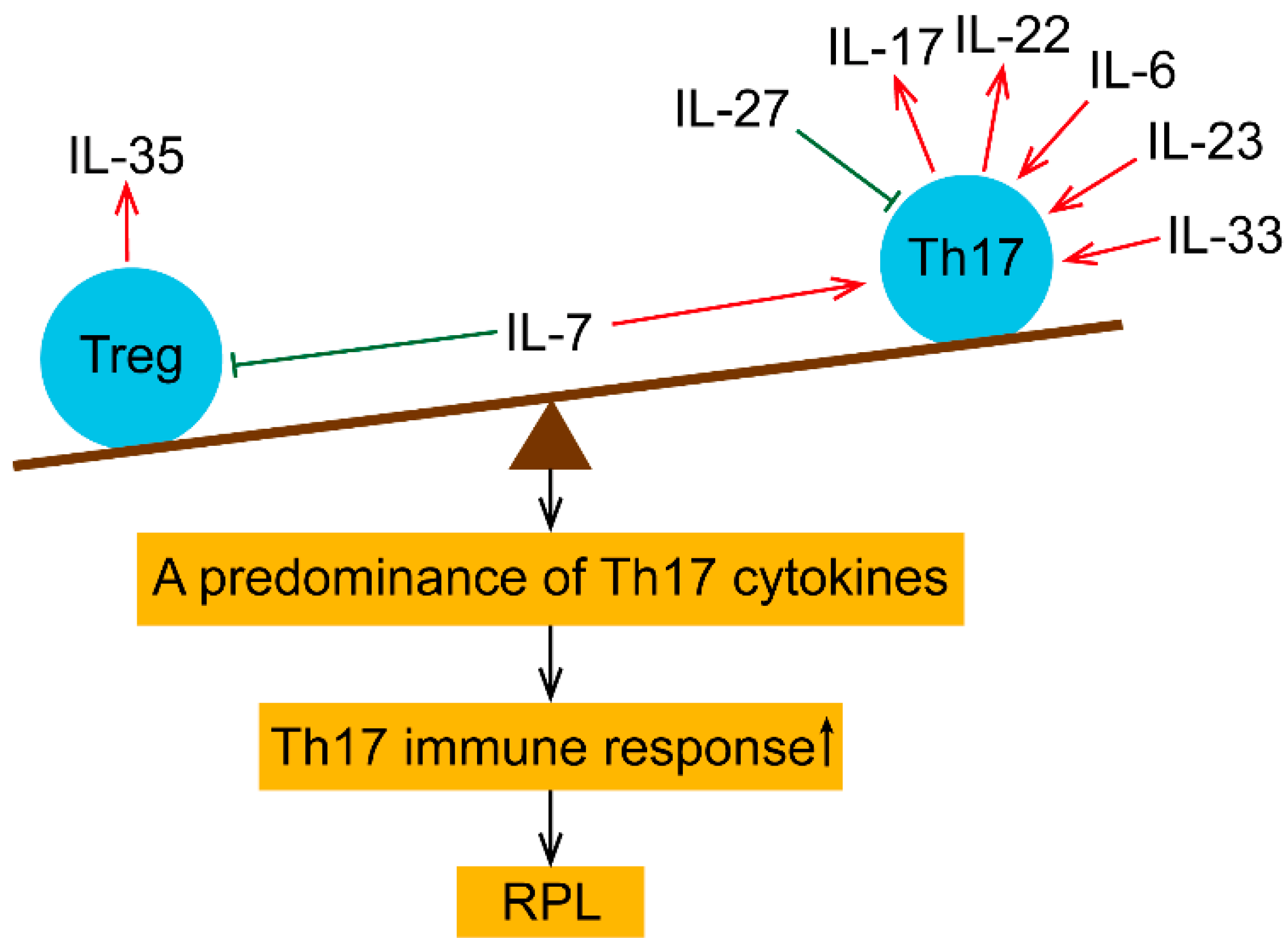
3. Treg and Th17-Related Cytokines in Maternal Peripheral Blood
3.1. IL-6
3.2. IL-7
3.3. IL-17
3.4. IL-22
3.5. IL-23
3.6. IL-27
3.7. IL-35
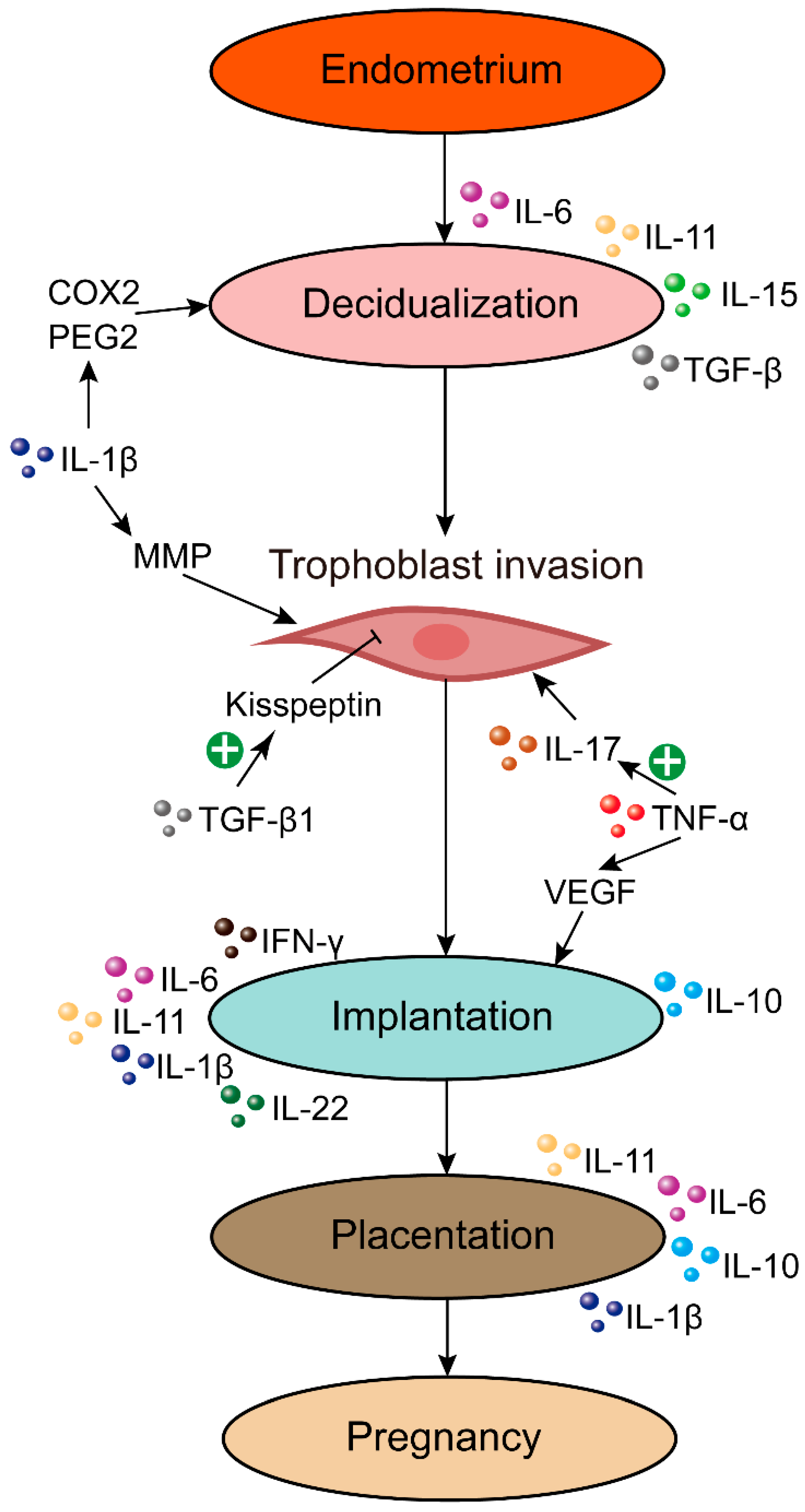
4. Endometrial Cytokine Imbalance in RPL
5. Cytokines Produced by NK Cells
5.1. Cytokine in the Micro Milieu of NK Cells
5.2. uNK Cells and Angiogenic Cytokines
6. Cytokine Gene Polymorphisms in RPL
6.1. TNF-α Gene Polymorphisms in RPL
6.2. IFN-γ Gene Polymorphisms in RPL
6.3. IL-10 Gene Polymorphisms in RPL
6.4. Other Cytokines Gene Polymorphisms in RPL
7. Hormonal Regulation of Cytokines and RPL
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- El Hachem, H.; Crepaux, V.; May-Panloup, P.; Descamps, P.; Legendre, G.; Bouet, P.E. Recurrent pregnancy loss: Current perspectives. Int. J. Women’s Health 2017, 9, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Youssef, A.; Vermeulen, N.; Lashley, E.; Goddijn, M.; van der Hoorn, M.L.P. Comparison and appraisal of (inter)national recurrent pregnancy loss guidelines. Reprod. Biomed. Online 2019, 39, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Bagkou Dimakou, D.; Lissauer, D.; Tamblyn, J.; Coomarasamy, A.; Richter, A. Understanding human immunity in idiopathic recurrent pregnancy loss. Eur. J. Obstet. Gynecol. Reprod. Biol. 2022, 270, 17–29. [Google Scholar] [CrossRef]
- Dambaeva, S.V.; Lee, D.H.; Sung, N.; Chen, C.Y.; Bao, S.; Gilman-Sachs, A.; Kwak-Kim, J.; Beaman, K.D. Recurrent Pregnancy Loss in Women with Killer Cell Immunoglobulin-Like Receptor KIR2DS1 is Associated with an Increased HLA-C2 Allelic Frequency. Am. J. Reprod. Immunol. 2016, 75, 94–103. [Google Scholar] [CrossRef]
- Ewington, L.J.; Tewary, S.; Brosens, J.J. New insights into the mechanisms underlying recurrent pregnancy loss. J. Obstet. Gynaecol. Res. 2019, 45, 258–265. [Google Scholar] [CrossRef]
- Nair, R.R.; Verma, P.; Singh, K. Immune-endocrine crosstalk during pregnancy. Gen. Comp. Endocrinol. 2017, 242, 18–23. [Google Scholar] [CrossRef]
- Saini, V.; Arora, S.; Yadav, A.; Bhattacharjee, J. Cytokines in recurrent pregnancy loss. Clin. Chim. Acta 2011, 412, 702–708. [Google Scholar] [CrossRef]
- Motedayyen, H.; Rezaei, A.; Zarnani, A.H.; Tajik, N. Human amniotic epithelial cells inhibit activation and pro-inflammatory cytokines production of naive CD4+ T cells from women with unexplained recurrent spontaneous abortion. Reprod. Biol. 2018, 18, 182–188. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Chen, X.; Xu, Z.Z.; Xu, L.; Mao, T.; Zhang, H. Changes and clinical significance of peripheral blood helper T lymphocyte and natural killer (NK) cells in unexplained recurrent spontaneous abortion (URSA) patients after abortion and successful pregnancy. Clin. Exp. Obstet. Gynecol. 2015, 42, 62–66. [Google Scholar] [CrossRef]
- Fu, B.; Li, X.; Sun, R.; Tong, X.; Ling, B.; Tian, Z.; Wei, H. Natural killer cells promote immune tolerance by regulating inflammatory TH17 cells at the human maternal-fetal interface. Proc. Natl. Acad. Sci. USA 2013, 110, E231–E240. [Google Scholar] [CrossRef]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Ernerudh, J. Decidual cytokines and pregnancy complications: Focus on spontaneous miscarriage. J. Reprod. Immunol. 2015, 108, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Chen, H.; Liu, M.; Yuan, Y.; Wang, Z.; Chen, Y.; Wei, J.; Su, F.; Zhang, J. Treg/Th17 Cell Imbalance and IL-6 Profile in Patients With Unexplained Recurrent Spontaneous Abortion. Reprod. Sci. 2017, 24, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, J.Y.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. Th17 and regulatory T cells in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2012, 67, 311–318. [Google Scholar] [CrossRef]
- Keller, C.C.; Eikmans, M.; van der Hoorn, M.P.; Lashley, L. Recurrent miscarriages and the association with regulatory T cells; A systematic review. J. Reprod. Immunol. 2020, 139, 103105. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Vacca, P.; Moretta, L.; Moretta, A.; Mingari, M.C. Origin, phenotype and function of human natural killer cells in pregnancy. Trends Immunol. 2011, 32, 517–523. [Google Scholar] [CrossRef]
- Chazara, O.; Xiong, S.; Moffett, A. Maternal KIR and fetal HLA-C: A fine balance. J. Leukoc. Biol. 2011, 90, 703–716. [Google Scholar] [CrossRef]
- Saito, S.; Nakashima, A.; Myojo-Higuma, S.; Shiozaki, A. The balance between cytotoxic NK cells and regulatory NK cells in human pregnancy. J. Reprod. Immunol. 2008, 77, 14–22. [Google Scholar] [CrossRef]
- Fu, Y.Y.; Ren, C.E.; Qiao, P.Y.; Meng, Y.H. Uterine natural killer cells and recurrent spontaneous abortion. Am. J. Reprod. Immunol. 2021, 86, e13433. [Google Scholar] [CrossRef]
- Maldonado, R.A.; von Andrian, U.H. How tolerogenic dendritic cells induce regulatory T cells. Adv. Immunol. 2010, 108, 111–165. [Google Scholar] [CrossRef] [Green Version]
- Salamone, G.; Fraccaroli, L.; Gori, S.; Grasso, E.; Paparini, D.; Geffner, J.; Pérez Leirós, C.; Ramhorst, R. Trophoblast cells induce a tolerogenic profile in dendritic cells. Hum. Reprod. 2012, 27, 2598–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilczyński, J.R. Cancer and pregnancy share similar mechanisms of immunological escape. Chemotherapy 2006, 52, 107–110. [Google Scholar] [CrossRef]
- Bansal, A.S. Joining the immunological dots in recurrent miscarriage. Am. J. Reprod. Immunol. 2010, 64, 307–315. [Google Scholar] [CrossRef]
- Chen, Q.; Carroll, H.P.; Gadina, M. The newest interleukins: Recent additions to the ever-growing cytokine family. Vitam. Horm. 2006, 74, 207–228. [Google Scholar] [CrossRef]
- Ali, S.; Majid, S.; Ali, M.N.; Taing, S.; Rehman, M.U.; Arafah, A. Cytokine imbalance at materno-embryonic interface as a potential immune mechanism for recurrent pregnancy loss. Int. Immunopharmacol. 2021, 90, 107118. [Google Scholar] [CrossRef]
- Vignali, D. How many mechanisms do regulatory T cells need? Eur. J. Immunol. 2008, 38, 908–911. [Google Scholar] [CrossRef]
- Murphy, S.P.; Tayade, C.; Ashkar, A.A.; Hatta, K.; Zhang, J.; Croy, B.A. Interferon gamma in successful pregnancies. Biol. Reprod. 2009, 80, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Lash, G.E.; Schiessl, B.; Kirkley, M.; Innes, B.A.; Cooper, A.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Expression of angiogenic growth factors by uterine natural killer cells during early pregnancy. J. Leukoc. Biol. 2006, 80, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Kwak-Kim, J.Y.; Chung-Bang, H.S.; Ng, S.C.; Ntrivalas, E.I.; Mangubat, C.P.; Beaman, K.D.; Beer, A.E.; Gilman-Sachs, A. Increased T helper 1 cytokine responses by circulating T cells are present in women with recurrent pregnancy losses and in infertile women with multiple implantation failures after IVF. Hum. Reprod. 2003, 18, 767–773. [Google Scholar] [CrossRef]
- Löb, S.; Amann, N.; Kuhn, C.; Schmoeckel, E.; Wöckel, A.; Zati Zehni, A.; Kaltofen, T.; Keckstein, S.; Mumm, J.N.; Meister, S.; et al. Interleukin-1 beta is significantly upregulated in the decidua of spontaneous and recurrent miscarriage placentas. J. Reprod. Immunol. 2021, 144, 103283. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, J.; Xu, H.L.; Xu, B.; Tong, X.H.; Kwak-Kim, J.; Liu, Y.S. IL-7/IL-7R signaling pathway might play a role in recurrent pregnancy losses by increasing inflammatory Th17 cells and decreasing Treg cells. Am. J. Reprod. Immunol. 2016, 76, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.A.; Polgar, K.; Anderson, D.J. T-helper 1-type immunity to trophoblast in women with recurrent spontaneous abortion. JAMA 1995, 273, 1933–1936. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yin, S.; Wang, M. Significance of the ratio interferon-γ/interleukin-4 in early diagnosis and immune mechanism of unexplained recurrent spontaneous abortion. Int. J. Gynaecol. Obstet. 2021, 154, 39–43. [Google Scholar] [CrossRef]
- Bates, M.D.; Quenby, S.; Takakuwa, K.; Johnson, P.M.; Vince, G.S. Aberrant cytokine production by peripheral blood mononuclear cells in recurrent pregnancy loss? Hum. Reprod. 2002, 17, 2439–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghupathy, R.; Makhseed, M.; Azizieh, F.; Hassan, N.; Al-Azemi, M.; Al-Shamali, E. Maternal Th1- and Th2-type reactivity to placental antigens in normal human pregnancy and unexplained recurrent spontaneous abortions. Cell. Immunol. 1999, 196, 122–130. [Google Scholar] [CrossRef]
- Raghupathy, R.; Makhseed, M.; Azizieh, F.; Omu, A.; Gupta, M.; Farhat, R. Cytokine production by maternal lymphocytes during normal human pregnancy and in unexplained recurrent spontaneous abortion. Hum. Reprod. 2000, 15, 713–718. [Google Scholar] [CrossRef] [Green Version]
- Makhseed, M.; Raghupathy, R.; Azizieh, F.; Al-Azemi, M.M.; Hassan, N.A.; Bandar, A. Mitogen-induced cytokine responses of maternal peripheral blood lymphocytes indicate a differential Th-type bias in normal pregnancy and pregnancy failure. Am. J. Reprod. Immunol. 1999, 42, 273–281. [Google Scholar] [CrossRef]
- Marzi, M.; Vigano, A.; Trabattoni, D.; Villa, M.L.; Salvaggio, A.; Clerici, E.; Clerici, M. Characterization of type 1 and type 2 cytokine production profile in physiologic and pathologic human pregnancy. Clin. Exp. Immunol. 1996, 106, 127–133. [Google Scholar] [CrossRef]
- Tranchot-Diallo, J.; Gras, G.; Parnet-Mathieu, F.; Benveniste, O.; Marcé, D.; Roques, P.; Milliez, J.; Chaouat, G.; Dormont, D. Modulations of cytokine expression in pregnant women. Am. J. Reprod. Immunol. 1997, 37, 215–226. [Google Scholar] [CrossRef]
- Fallon, P.G.; Jolin, H.E.; Smith, P.; Emson, C.L.; Townsend, M.J.; Fallon, R.; Smith, P.; McKenzie, A.N. IL-4 induces characteristic Th2 responses even in the combined absence of IL-5, IL-9, and IL-13. Immunity 2002, 17, 7–17. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Stelzl, P.; Joseph, D.N.; Aldo, P.B.; Maxwell, A.J.; Dekel, N.; Liao, A.; Whirledge, S.; Mor, G. TNF-α Regulated Endometrial Stroma Secretome Promotes Trophoblast Invasion. Front. Immunol. 2021, 12, 737401. [Google Scholar] [CrossRef]
- Chung, I.B.; Yelian, F.D.; Zaher, F.M.; Gonik, B.; Evans, M.I.; Diamond, M.P.; Svinarich, D.M. Expression and regulation of vascular endothelial growth factor in a first trimester trophoblast cell line. Placenta 2000, 21, 320–324. [Google Scholar] [CrossRef]
- Lio, D.; Candore, G.; Colombo, A.; Colonna Romano, G.; Gervasi, F.; Marino, V.; Scola, L.; Caruso, C. A genetically determined high setting of TNF-alpha influences immunologic parameters of HLA-B8,DR3 positive subjects: Implications for autoimmunity. Hum. Immunol. 2001, 62, 705–713. [Google Scholar] [CrossRef]
- Ng, S.C.; Gilman-Sachs, A.; Thaker, P.; Beaman, K.D.; Beer, A.E.; Kwak-Kim, J. Expression of intracellular Th1 and Th2 cytokines in women with recurrent spontaneous abortion, implantation failures after IVF/ET or normal pregnancy. Am. J. Reprod. Immunol. 2002, 48, 77–86. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Chrousos, G.P.; Wilder, R.L. Neuroendocrine regulation of IL-12 and TNF-alpha/IL-10 balance. Clinical implications. Ann. N. Y. Acad. Sci. 2000, 917, 94–105. [Google Scholar] [CrossRef]
- Dominguez, F.; Gadea, B.; Mercader, A.; Esteban, F.J.; Pellicer, A.; Simón, C. Embryologic outcome and secretome profile of implanted blastocysts obtained after coculture in human endometrial epithelial cells versus the sequential system. Fertil. Steril. 2010, 93, 774–782.e1. [Google Scholar] [CrossRef]
- Saifi, B.; Rezaee, S.A.; Tajik, N.; Ahmadpour, M.E.; Ashrafi, M.; Vakili, R.; SoleimaniAsl, S.; Aflatoonian, R.; Mehdizadeh, M. Th17 cells and related cytokines in unexplained recurrent spontaneous miscarriage at the implantation window. Reprod. Biomed. Online 2014, 29, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Salama, K.M.; Alloush, M.K.; Al Hussini, R.M. Are the cytokines TNF alpha and IL 1Beta early predictors of embryo implantation? Cross sectional study. J. Reprod. Immunol. 2020, 137, 102618. [Google Scholar] [CrossRef]
- Guo, F.; Si, C.; Zhou, M.; Wang, J.; Zhang, D.; Leung, P.C.K.; Xu, B.; Zhang, A. Decreased PECAM1-mediated TGF-β1 expression in the mid-secretory endometrium in women with recurrent implantation failure. Hum. Reprod. 2018, 33, 832–843. [Google Scholar] [CrossRef]
- Bahrami-Asl, Z.; Hajipour, H.; Rastgar Rezaei, Y.; Novinbahador, T.; Latifi, Z.; Nejabati, H.R.; Farzadi, L.; Fattahi, A.; Nouri, M.; Dominguez, F. Cytokines in embryonic secretome as potential markers for embryo selection. Am. J. Reprod. Immunol. 2021, 85, e13385. [Google Scholar] [CrossRef]
- Hirota, Y.; Osuga, Y.; Hasegawa, A.; Kodama, A.; Tajima, T.; Hamasaki, K.; Koga, K.; Yoshino, O.; Hirata, T.; Harada, M.; et al. Interleukin (IL)-1beta stimulates migration and survival of first-trimester villous cytotrophoblast cells through endometrial epithelial cell-derived IL-8. Endocrinology 2009, 150, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Y.; Chen, H.F.; Chen, S.U.; Chao, K.H.; Yang, Y.S.; Ho, H.N. Increase in the production of interleukin-10 early after implantation is related to the success of pregnancy. Am. J. Reprod. Immunol. 2001, 46, 386–392. [Google Scholar] [CrossRef]
- Dimitriadis, E.; Robb, L.; Liu, Y.X.; Enders, A.C.; Martin, H.; Stoikos, C.; Wallace, E.; Salamonsen, L.A. IL-11 and IL-11Ralpha immunolocalisation at primate implantation sites supports a role for IL-11 in placentation and fetal development. Reprod. Biol. Endocrinol. 2003, 1, 34. [Google Scholar] [CrossRef]
- Logiodice, F.; Lombardelli, L.; Kullolli, O.; Haller, H.; Maggi, E.; Rukavina, D.; Piccinni, M.P. Decidual Interleukin-22-Producing CD4+ T Cells (Th17/Th0/IL-22+ and Th17/Th2/IL-22+, Th2/IL-22+, Th0/IL-22+), Which Also Produce IL-4, Are Involved in the Success of Pregnancy. Int. J. Mol. Sci. 2019, 20, 428. [Google Scholar] [CrossRef] [Green Version]
- Salker, M.S.; Nautiyal, J.; Steel, J.H.; Webster, Z.; Sućurović, S.; Nicou, M.; Singh, Y.; Lucas, E.S.; Murakami, K.; Chan, Y.W.; et al. Disordered IL-33/ST2 activation in decidualizing stromal cells prolongs uterine receptivity in women with recurrent pregnancy loss. PLoS ONE 2012, 7, e52252. [Google Scholar] [CrossRef] [Green Version]
- Makhseed, M.; Raghupathy, R.; Azizieh, F.; Farhat, R.; Hassan, N.; Bandar, A. Circulating cytokines and CD30 in normal human pregnancy and recurrent spontaneous abortions. Hum. Reprod. 2000, 15, 2011–2017. [Google Scholar] [CrossRef] [Green Version]
- Arslan, E.; Colakoğlu, M.; Celik, C.; Gezginç, K.; Acar, A.; Capar, M.; Aköz, M.; Akyürek, C. Serum TNF-alpha, IL-6, lupus anticoagulant and anticardiolipin antibody in women with and without a past history of recurrent miscarriage. Arch. Gynecol. Obstet. 2004, 270, 227–229. [Google Scholar] [CrossRef]
- Piosik, Z.M.; Goegebeur, Y.; Klitkou, L.; Steffensen, R.; Christiansen, O.B. Plasma TNF-α levels are higher in early pregnancy in patients with secondary compared with primary recurrent miscarriage. Am. J. Reprod. Immunol. 2013, 70, 347–358. [Google Scholar] [CrossRef]
- Garcia-Lloret, M.I.; Winkler-Lowen, B.; Guilbert, L.J. Monocytes adhering by LFA-1 to placental syncytiotrophoblasts induce local apoptosis via release of TNF-alpha. A model for hematogenous initiation of placental inflammations. J. Leukoc. Biol. 2000, 68, 903–908. [Google Scholar] [CrossRef]
- Fortunato, S.J.; Menon, R.; Lombardi, S.J. Support for an infection-induced apoptotic pathway in human fetal membranes. Am. J. Obstet. Gynecol. 2001, 184, 1392–1397; discussion 1397–1398. [Google Scholar] [CrossRef]
- Berman, J.; Girardi, G.; Salmon, J.E. TNF-alpha is a critical effector and a target for therapy in antiphospholipid antibody-induced pregnancy loss. J. Immunol. 2005, 174, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, P.; Jana, S.K.; Pasricha, P.; Ghosh, S.; Chakravarty, B.; Chaudhury, K. Proinflammatory cytokines induced altered expression of cyclooxygenase-2 gene results in unreceptive endometrium in women with idiopathic recurrent spontaneous miscarriage. Fertil. Steril. 2013, 99, 179–187.e2. [Google Scholar] [CrossRef] [PubMed]
- Hossein, H.; Mahroo, M.; Abbas, A.; Firouzeh, A.; Nadia, H. Cytokine production by peripheral blood mononuclear cells in recurrent miscarriage. Cytokine 2004, 28, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Daher, S.; de Arruda Geraldes Denardi, K.; Blotta, M.H.; Mamoni, R.L.; Reck, A.P.; Camano, L.; Mattar, R. Cytokines in recurrent pregnancy loss. J. Reprod. Immunol. 2004, 62, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Zidan, H.E.; Abdul-Maksoud, R.S.; Mowafy, H.E.; Elsayed, W.S.H. The association of IL-33 and Foxp3 gene polymorphisms with recurrent pregnancy loss in Egyptian women. Cytokine 2018, 108, 115–119. [Google Scholar] [CrossRef]
- Cai, J.; Li, M.; Huang, Q.; Fu, X.; Wu, H. Differences in Cytokine Expression and STAT3 Activation between Healthy Controls and Patients of Unexplained Recurrent Spontaneous Abortion (URSA) during Early Pregnancy. PLoS ONE 2016, 11, e0163252. [Google Scholar] [CrossRef] [Green Version]
- Roomandeh, N.; Saremi, A.; Arasteh, J.; Pak, F.; Mirmohammadkhani, M.; Kokhaei, P.; Zare, A. Comparing Serum Levels of Th17 and Treg Cytokines in Women with Unexplained Recurrent Spontaneous Abortion and Fertile Women. Iran. J. Immunol. 2018, 15, 59–67. [Google Scholar]
- Wang, W.J.; Hao, C.F.; Yi, L.; Yin, G.J.; Bao, S.H.; Qiu, L.H.; Lin, Q.D. Increased prevalence of T helper 17 (Th17) cells in peripheral blood and decidua in unexplained recurrent spontaneous abortion patients. J. Reprod. Immunol. 2010, 84, 164–170. [Google Scholar] [CrossRef]
- Ozkan, Z.S.; Deveci, D.; Simsek, M.; Ilhan, F.; Risvanli, A.; Sapmaz, E. What is the impact of SOCS3, IL-35 and IL17 in immune pathogenesis of recurrent pregnancy loss? J. Matern. Fetal Neonatal Med. 2015, 28, 324–328. [Google Scholar] [CrossRef]
- Yue, C.Y.; Zhang, B.; Ying, C.M. Elevated Serum Level of IL-35 Associated with the Maintenance of Maternal-Fetal Immune Tolerance in Normal Pregnancy. PLoS ONE 2015, 10, e0128219. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, G.R.; Collins, J.E.; Currie, J.K.; Guyre, P.M.; Wira, C.R.; Fanger, M.W. IFN-gamma is produced by polymorphonuclear neutrophils in human uterine endometrium and by cultured peripheral blood polymorphonuclear neutrophils. J. Immunol. 1998, 160, 5145–5153. [Google Scholar] [PubMed]
- Nasu, K.; Matsui, N.; Narahara, H.; Tanaka, Y.; Miyakawa, I. Effects of interferon-gamma on cytokine production by endometrial stromal cells. Hum. Reprod. 1998, 13, 2598–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calleja-Agius, J.; Jauniaux, E.; Pizzey, A.R.; Muttukrishna, S. Investigation of systemic inflammatory response in first trimester pregnancy failure. Hum. Reprod. 2012, 27, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Palfi, M.; Jablonowska, B.; Matthiesen, L.; Ernerudh, J. Circulating interferon-gamma- and interleukin-4-secreting cells in recurrent spontaneous abortions. Am. J. Reprod. Immunol. 1999, 41, 257–263. [Google Scholar] [CrossRef]
- Robertson, S.A.; Seamark, R.F.; Guilbert, L.J.; Wegmann, T.G. The role of cytokines in gestation. Crit. Rev. Immunol. 1994, 14, 239–292. [Google Scholar] [CrossRef]
- Fontana, V.; Choren, V.; Vauthay, L.; Calvo, J.C.; Calvo, L.; Cameo, M. Exogenous interferon-gamma alters murine inner cell mass and trophoblast development. Effect on the expression of ErbB1, ErbB4 and heparan sulfate proteoglycan (perlecan). Reproduction 2004, 128, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, T.; Suzuki, T.; Aizawa, K.; Sawaki, D.; Munemasa, Y.; Ishida, J.; Nagai, R. Regulation of transforming growth factor-beta-dependent cyclooxygenase-2 expression in fibroblasts. J. Biol. Chem. 2009, 284, 35861–35871. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.L.; Huston, G.; Tonkonogy, S.; Weinberg, A. Transforming growth factor-beta and IL-4 cause helper T cell precursors to develop into distinct effector helper cells that differ in lymphokine secretion pattern and cell surface phenotype. J. Immunol. 1991, 147, 2991–3000. [Google Scholar]
- Freis, A.; Schlegel, J.; Daniel, V.; Jauckus, J.; Strowitzki, T.; Germeyer, A. Cytokines in relation to hCG are significantly altered in asymptomatic women with miscarriage—A pilot study. Reprod. Biol. Endocrinol. 2018, 16, 93. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, M.S.; Aoki, K.; Aoyama, T.; Katano, K.; Iinuma, Y.; Ozaki, Y.; Suzumori, K. Elevation of transforming growth factor-beta1 is associated with recurrent miscarriage. J. Clin. Immunol. 2000, 20, 453–457. [Google Scholar] [CrossRef]
- Fang, L.; Yan, Y.; Gao, Y.; Wu, Z.; Wang, Z.; Yang, S.; Cheng, J.C.; Sun, Y.P. TGF-β1 inhibits human trophoblast cell invasion by upregulating kisspeptin expression through ERK1/2 but not SMAD signaling pathway. Reprod. Biol. Endocrinol. 2022, 20, 22. [Google Scholar] [CrossRef] [PubMed]
- Hefler, L.A.; Tempfer, C.B.; Unfried, G.; Schneeberger, C.; Lessl, K.; Nagele, F.; Huber, J.C. A polymorphism of the interleukin-1beta gene and idiopathic recurrent miscarriage. Fertil. Steril. 2001, 76, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Nilkaeo, A.; Bhuvanath, S. Interleukin-1 modulation of human placental trophoblast proliferation. Mediat. Inflamm. 2006, 2006, 79359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantos, K.; Grigoriadis, S.; Maziotis, E.; Pistola, K.; Xystra, P.; Pantou, A.; Kokkali, G.; Pappas, A.; Lambropoulou, M.; Sfakianoudis, K.; et al. The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature. Int. J. Mol. Sci. 2022, 23, 2198. [Google Scholar] [CrossRef] [PubMed]
- Furcron, A.E.; Romero, R.; Mial, T.N.; Balancio, A.; Panaitescu, B.; Hassan, S.S.; Sahi, A.; Nord, C.; Gomez-Lopez, N. Human Chorionic Gonadotropin Has Anti-Inflammatory Effects at the Maternal-Fetal Interface and Prevents Endotoxin-Induced Preterm Birth, but Causes Dystocia and Fetal Compromise in Mice. Biol. Reprod. 2016, 94, 136. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Kong, L.; Qing, P.; Cheng, K.; Chen, H.; Zhang, S.; Hu, X.; Hu, Z.; Yu, F.; Qin, L. Interleukin-1β as clinically predictive risk marker for recurrent pregnancy loss in women positive for antinuclear antibody. Int. J. Clin. Pract. 2021, 75, e14887. [Google Scholar] [CrossRef]
- Spandorfer, S.D.; Neuer, A.; Liu, H.C.; Rosenwaks, Z.; Witkin, S.S. Involvement of interleukin-1 and the interleukin-1 receptor antagonist in in vitro embryo development among women undergoing in vitro fertilization-embryo transfer. J. Assist. Reprod. Genet. 2003, 20, 502–505. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.P. Arterial thrombosis--insidious, unpredictable and deadly. Nat. Med. 2011, 17, 1423–1436. [Google Scholar] [CrossRef]
- Haraldsen, G.; Balogh, J.; Pollheimer, J.; Sponheim, J.; Küchler, A.M. Interleukin-33—Cytokine of dual function or novel alarmin? Trends Immunol. 2009, 30, 227–233. [Google Scholar] [CrossRef]
- Smithgall, M.D.; Comeau, M.R.; Yoon, B.R.; Kaufman, D.; Armitage, R.; Smith, D.E. IL-33 amplifies both Th1- and Th2-type responses through its activity on human basophils, allergen-reactive Th2 cells, iNKT and NK cells. Int. Immunol. 2008, 20, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Jiang, H.R.; Kewin, P.; Li, Y.; Mu, R.; Fraser, A.R.; Pitman, N.; Kurowska-Stolarska, M.; McKenzie, A.N.; McInnes, I.B.; et al. IL-33 exacerbates antigen-induced arthritis by activating mast cells. Proc. Natl. Acad. Sci. USA 2008, 105, 10913–10918. [Google Scholar] [CrossRef] [PubMed]
- Nakae, S.; Morita, H.; Ohno, T.; Arae, K.; Matsumoto, K.; Saito, H. Role of interleukin-33 in innate-type immune cells in allergy. Allergol. Int. 2013, 62, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgeois, E.; Van, L.P.; Samson, M.; Diem, S.; Barra, A.; Roga, S.; Gombert, J.M.; Schneider, E.; Dy, M.; Gourdy, P.; et al. The pro-Th2 cytokine IL-33 directly interacts with invariant NKT and NK cells to induce IFN-gamma production. Eur. J. Immunol. 2009, 39, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Gajardo Carrasco, T.; Morales, R.A.; Pérez, F.; Terraza, C.; Yáñez, L.; Campos-Mora, M.; Pino-Lagos, K. Alarmin’ Immunologists: IL-33 as a Putative Target for Modulating T Cell-Dependent Responses. Front. Immunol. 2015, 6, 232. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.; Tong, Y.; Xie, L.; Ma, T.; Yang, J. Genetic variant in IL-33 is associated with idiopathic recurrent miscarriage in Chinese Han population. Sci. Rep. 2016, 6, 23806. [Google Scholar] [CrossRef] [Green Version]
- Kaitu’u-Lino, T.J.; Tuohey, L.; Tong, S. Maternal serum interleukin-33 and soluble ST2 across early pregnancy, and their association with miscarriage. J. Reprod. Immunol. 2012, 95, 46–49. [Google Scholar] [CrossRef]
- Hu, W.T.; Li, M.Q.; Liu, W.; Jin, L.P.; Li, D.J.; Zhu, X.Y. IL-33 enhances proliferation and invasiveness of decidual stromal cells by up-regulation of CCL2/CCR2 via NF-κB and ERK1/2 signaling. Mol. Hum. Reprod. 2014, 20, 358–372. [Google Scholar] [CrossRef] [Green Version]
- Prins, J.R.; Gomez-Lopez, N.; Robertson, S.A. Interleukin-6 in pregnancy and gestational disorders. J. Reprod. Immunol. 2012, 95, 1–14. [Google Scholar] [CrossRef]
- Arntzen, K.J.; Lien, E.; Austgulen, R. Maternal serum levels of interleukin-6 and clinical characteristics of normal delivery at term. Acta Obstet. Gynecol. Scand. 1997, 76, 55–60. [Google Scholar] [CrossRef]
- Oláh, K.S.; Vince, G.S.; Neilson, J.P.; Deniz, G.; Johnson, P.M. Interleukin-6, interferon-gamma, interleukin-8, and granulocyte-macrophage colony stimulating factor levels in human amniotic fluid at term. J. Reprod. Immunol. 1996, 32, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Steinborn, A.; Günes, H.; Röddiger, S.; Halberstadt, E. Elevated placental cytokine release, a process associated with preterm labor in the absence of intrauterine infection. Obstet. Gynecol. 1996, 88, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Interleukin-6 Family Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef] [Green Version]
- Sabat, R.; Grütz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.J.; Hao, C.F.; Lin, Q.D. Dysregulation of macrophage activation by decidual regulatory T cells in unexplained recurrent miscarriage patients. J. Reprod. Immunol. 2011, 92, 97–102. [Google Scholar] [CrossRef]
- Chaouat, G.; Assal Meliani, A.; Martal, J.; Raghupathy, R.; Elliott, J.F.; Mosmann, T.; Wegmann, T.G. IL-10 prevents naturally occurring fetal loss in the CBA x DBA/2 mating combination, and local defect in IL-10 production in this abortion-prone combination is corrected by in vivo injection of IFN-tau. J. Immunol. 1995, 154, 4261–4268. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S.A.; Care, A.S.; Skinner, R.J. Interleukin 10 regulates inflammatory cytokine synthesis to protect against lipopolysaccharide-induced abortion and fetal growth restriction in mice. Biol. Reprod. 2007, 76, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C.; Roberts, J.; Wilson, R.; MacLean, M.A.; Shilito, J.; Walker, J.J. Evidence of a T(H) 1 type response associated with recurrent miscarriage. Fertil. Steril. 2000, 73, 1206–1208. [Google Scholar] [CrossRef]
- Hasnain, S.Z.; Tauro, S.; Das, I.; Tong, H.; Chen, A.C.; Jeffery, P.L.; McDonald, V.; Florin, T.H.; McGuckin, M.A. IL-10 promotes production of intestinal mucus by suppressing protein misfolding and endoplasmic reticulum stress in goblet cells. Gastroenterology 2013, 144, 357–368.e9. [Google Scholar] [CrossRef]
- Shkoda, A.; Ruiz, P.A.; Daniel, H.; Kim, S.C.; Rogler, G.; Sartor, R.B.; Haller, D. Interleukin-10 blocked endoplasmic reticulum stress in intestinal epithelial cells: Impact on chronic inflammation. Gastroenterology 2007, 132, 190–207. [Google Scholar] [CrossRef]
- Yang, H.; Qiu, L.; Chen, G.; Ye, Z.; Lü, C.; Lin, Q. Proportional change of CD4+CD25+ regulatory T cells in decidua and peripheral blood in unexplained recurrent spontaneous abortion patients. Fertil. Steril. 2008, 89, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.S.; Wu, L.; Tong, X.H.; Wu, L.M.; He, G.P.; Zhou, G.X.; Luo, L.H.; Luan, H.B. Study on the relationship between Th17 cells and unexplained recurrent spontaneous abortion. Am. J. Reprod. Immunol. 2011, 65, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, J.Y.; Hur, S.E.; Kim, C.J.; Na, B.J.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. An imbalance in interleukin-17-producing T and Foxp3⁺ regulatory T cells in women with idiopathic recurrent pregnancy loss. Hum. Reprod. 2011, 26, 2964–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, A.; Ito, M.; Shima, T.; Bac, N.D.; Hidaka, T.; Saito, S. Accumulation of IL-17-positive cells in decidua of inevitable abortion cases. Am. J. Reprod. Immunol. 2010, 64, 4–11. [Google Scholar] [CrossRef]
- Piccinni, M.P. T cell tolerance towards the fetal allograft. J. Reprod. Immunol. 2010, 85, 71–75. [Google Scholar] [CrossRef]
- Arruvito, L.; Billordo, A.; Capucchio, M.; Prada, M.E.; Fainboim, L. IL-6 trans-signaling and the frequency of CD4+FOXP3+ cells in women with reproductive failure. J. Reprod. Immunol. 2009, 82, 158–165. [Google Scholar] [CrossRef]
- Pasare, C.; Medzhitov, R. Toll pathway-dependent blockade of CD4+CD25+ T cell-mediated suppression by dendritic cells. Science 2003, 299, 1033–1036. [Google Scholar] [CrossRef]
- Corthay, A. How do regulatory T cells work? Scand. J. Immunol. 2009, 70, 326–336. [Google Scholar] [CrossRef]
- Vilsmaier, T.; Amann, N.; Löb, S.; Schmoeckel, E.; Kuhn, C.; Zati Zehni, A.; Meister, S.; Beyer, S.; Kolben, T.M.; Becker, J.; et al. The decidual expression of Interleukin-7 is upregulated in early pregnancy loss. Am. J. Reprod. Immunol. 2021, 86, e13437. [Google Scholar] [CrossRef]
- Kwak, J.Y.; Beer, A.E.; Kim, S.H.; Mantouvalos, H.P. Immunopathology of the implantation site utilizing monoclonal antibodies to natural killer cells in women with recurrent pregnancy losses. Am. J. Reprod. Immunol. 1999, 41, 91–98. [Google Scholar] [CrossRef]
- Haas, J.; Korporal, M.; Schwarz, A.; Balint, B.; Wildemann, B. The interleukin-7 receptor α chain contributes to altered homeostasis of regulatory T cells in multiple sclerosis. Eur. J. Immunol. 2011, 41, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Hartgring, S.A.; Bijlsma, J.W.; Lafeber, F.P.; van Roon, J.A. Interleukin-7 induced immunopathology in arthritis. Ann. Rheum. Dis. 2006, 65 (Suppl. 3), iii69–iii74. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, W.; Xie, H.; Xu, X.; Wu, J.; Jiang, Z.; Zhang, M.; Zhou, L.; Zheng, S. A pathogenic role of IL- 17 at the early stage of corneal allograft rejection. Transpl. Immunol. 2009, 21, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Paez-Cortez, J.; Schmitt-Knosalla, I.; D’Addio, F.; Mfarrej, B.; Donnarumma, M.; Habicht, A.; Clarkson, M.R.; Iacomini, J.; Glimcher, L.H.; et al. A novel role of CD4 Th17 cells in mediating cardiac allograft rejection and vasculopathy. J. Exp. Med. 2008, 205, 3133–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.J.; Liu, F.J.; Qu, H.M.; Hao, C.F.; Qu, Q.L.; Xiong, W.; Bao, H.C.; Wang, X.R. Regulation of the expression of Th17 cells and regulatory T cells by IL-27 in patients with unexplained early recurrent miscarriage. J. Reprod. Immunol. 2013, 99, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.X.; Jin, L.P.; Xu, B.; Liang, S.S.; Li, D.J. Decidual stromal cells recruit Th17 cells into decidua to promote proliferation and invasion of human trophoblast cells by secreting IL-17. Cell. Mol. Immunol. 2014, 11, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, V.L.; Ellwanger, J.H.; Matte, M.C.C.; Savaris, R.F.; Vianna, P.; Chies, J.A.B. IL-17 blood levels increase in healthy pregnancy but not in spontaneous abortion. Mol. Biol. Rep. 2018, 45, 1565–1568. [Google Scholar] [CrossRef]
- Xu, W.M.; Xiao, Z.N.; Wang, X.B.; Huang, Y. IL-17 Induces Fetal Loss in a CBA/J×BALB/c Mouse Model, and an Anti-IL-17 Antibody Prevents Fetal Loss in a CBA/J×DBA/2 Mouse Model. Am. J. Reprod. Immunol. 2016, 75, 51–58. [Google Scholar] [CrossRef]
- Lindemans, C.A.; Calafiore, M.; Mertelsmann, A.M.; O’Connor, M.H.; Dudakov, J.A.; Jenq, R.R.; Velardi, E.; Young, L.F.; Smith, O.M.; Lawrence, G.; et al. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial regeneration. Nature 2015, 528, 560–564. [Google Scholar] [CrossRef] [Green Version]
- McGee, H.M.; Schmidt, B.A.; Booth, C.J.; Yancopoulos, G.D.; Valenzuela, D.M.; Murphy, A.J.; Stevens, S.; Flavell, R.A.; Horsley, V. IL-22 promotes fibroblast-mediated wound repair in the skin. J. Investig. Dermatol. 2013, 133, 1321–1329. [Google Scholar] [CrossRef] [Green Version]
- Uz, Y.H.; Murk, W.; Yetkin, C.E.; Kayisli, U.A.; Arici, A. Expression and role of interleukin-23 in human endometrium throughout the menstrual cycle and early pregnancy. J. Reprod. Immunol. 2010, 87, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Coulomb-L’Herminé, A.; Larousserie, F.; Pflanz, S.; Bardel, E.; Kastelein, R.A.; Devergne, O. Expression of interleukin-27 by human trophoblast cells. Placenta 2007, 28, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Mas, A.E.; Petitbarat, M.; Dubanchet, S.; Fay, S.; Ledée, N.; Chaouat, G. Immune regulation at the interface during early steps of murine implantation: Involvement of two new cytokines of the IL-12 family (IL-23 and IL-27) and of TWEAK. Am. J. Reprod. Immunol. 2008, 59, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Chaouat, G.; Petitbarat, M.; Bulla, R.; Dubanchet, S.; Valdivia, K.; Ledée, N.; Steffen, T.; Jensenius, J.C.; Tedesco, F. Early regulators in abortion and implications for a preeclampsia model. J. Reprod. Immunol. 2009, 82, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef]
- Mao, H.; Gao, W.; Ma, C.; Sun, J.; Liu, J.; Shao, Q.; Song, B.; Qu, X. Human placental trophoblasts express the immunosuppressive cytokine IL-35. Hum. Immunol. 2013, 74, 872–877. [Google Scholar] [CrossRef]
- Qian, J.; Zhang, N.; Lin, J.; Wang, C.; Pan, X.; Chen, L.; Li, D.; Wang, L. Distinct pattern of Th17/Treg cells in pregnant women with a history of unexplained recurrent spontaneous abortion. Biosci. Trends 2018, 12, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Li, J.; Huang, S.Y.; Sun, X. Characterization of the subsets of human NKT-like cells and the expression of Th1/Th2 cytokines in patients with unexplained recurrent spontaneous abortion. J. Reprod. Immunol. 2015, 110, 81–88. [Google Scholar] [CrossRef]
- Jasper, M.J.; Tremellen, K.P.; Robertson, S.A. Reduced expression of IL-6 and IL-1alpha mRNAs in secretory phase endometrium of women with recurrent miscarriage. J. Reprod. Immunol. 2007, 73, 74–84. [Google Scholar] [CrossRef]
- Linjawi, S.; Li, T.C.; Tuckerman, E.M.; Blakemore, A.I.; Laird, S.M. Expression of interleukin-11 receptor alpha and interleukin-11 protein in the endometrium of normal fertile women and women with recurrent miscarriage. J. Reprod. Immunol. 2004, 64, 145–155. [Google Scholar] [CrossRef]
- Löb, S.; Ochmann, B.; Ma, Z.; Vilsmaier, T.; Kuhn, C.; Schmoeckel, E.; Herbert, S.L.; Kolben, T.; Wöckel, A.; Mahner, S.; et al. The role of Interleukin-18 in recurrent early pregnancy loss. J. Reprod. Immunol. 2021, 148, 103432. [Google Scholar] [CrossRef] [PubMed]
- Tabibzadeh, S. Ubiquitous expression of TNF-alpha/cachectin immunoreactivity in human endometrium. Am. J. Reprod. Immunol. 1991, 26, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.S.; Chen, H.L.; Hu, X.L.; Tabibzadeh, S. Tumor necrosis factor-alpha messenger ribonucleic acid and protein in human endometrium. Biol. Reprod. 1992, 47, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippeaux, M.M.; Piguet, P.F. Expression of tumor necrosis factor-alpha and its mRNA in the endometrial mucosa during the menstrual cycle. Am. J. Pathol. 1993, 143, 480–486. [Google Scholar]
- Arck, P.C.; Troutt, A.B.; Clark, D.A. Soluble receptors neutralizing TNF-alpha and IL-1 block stress-triggered murine abortion. Am. J. Reprod. Immunol. 1997, 37, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Otun, H.A.; Lash, G.E.; Innes, B.A.; Bulmer, J.N.; Naruse, K.; Hannon, T.; Searle, R.F.; Robson, S.C. Effect of tumour necrosis factor-α in combination with interferon-γ on first trimester extravillous trophoblast invasion. J. Reprod. Immunol. 2011, 88, 1–11. [Google Scholar] [CrossRef]
- Lim, K.J.; Odukoya, O.A.; Ajjan, R.A.; Li, T.C.; Weetman, A.P.; Cooke, I.D. The role of T-helper cytokines in human reproduction. Fertil. Steril. 2000, 73, 136–142. [Google Scholar] [CrossRef]
- Von Wolff, M.; Thaler, C.J.; Strowitzki, T.; Broome, J.; Stolz, W.; Tabibzadeh, S. Regulated expression of cytokines in human endometrium throughout the menstrual cycle: Dysregulation in habitual abortion. Mol. Hum. Reprod. 2000, 6, 627–634. [Google Scholar] [CrossRef]
- Lin, H.; Mosmann, T.R.; Guilbert, L.; Tuntipopipat, S.; Wegmann, T.G. Synthesis of T helper 2-type cytokines at the maternal-fetal interface. J. Immunol. 1993, 151, 4562–4573. [Google Scholar]
- Bennett, W.A.; Lagoo-Deenadayalan, S.; Whitworth, N.S.; Brackin, M.N.; Hale, E.; Cowan, B.D. Expression and production of interleukin-10 by human trophoblast: Relationship to pregnancy immunotolerance. Early Pregnancy 1997, 3, 190–198. [Google Scholar]
- Zhang, A.; Fu, J.; Ning, B.; Li, D.; Sun, N.; Wei, W.; Wei, J.; Ju, X. Tolerogenic dendritic cells generated with IL-10/TGFβ1 relieve immune thrombocytopenia in mice. Thromb. Res. 2013, 132, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Costa, S.D.; Zenclussen, A.C. Endocrine factors modulating immune responses in pregnancy. Front. Immunol. 2014, 5, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Qiu, T.; Wang, Y.; Chen, Y.; Cheng, W. Expression of C-type lectin receptors and Toll-like receptors in decidua of patients with unexplained recurrent spontaneous abortion. Reprod. Fertil. Dev. 2017, 29, 1613–1624. [Google Scholar] [CrossRef]
- Bao, S.H.; Wang, X.P.; De Lin, Q.; Wang, W.J.; Yin, G.J.; Qiu, L.H. Decidual CD4+CD25+CD127dim/− regulatory T cells in patients with unexplained recurrent spontaneous miscarriage. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 155, 94–98. [Google Scholar] [CrossRef]
- Piccinni, M.P.; Beloni, L.; Livi, C.; Maggi, E.; Scarselli, G.; Romagnani, S. Defective production of both leukemia inhibitory factor and type 2 T-helper cytokines by decidual T cells in unexplained recurrent abortions. Nat. Med. 1998, 4, 1020–1024. [Google Scholar] [CrossRef] [PubMed]
- Thaxton, J.E.; Sharma, S. Interleukin-10: A multi-faceted agent of pregnancy. Am. J. Reprod. Immunol. 2010, 63, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2019, 20, 5332. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Dong, P.; Jia, N.; Wen, X.; Luo, L.; Wang, S.; Li, J. The expression of intracellular cytokines of decidual natural killer cells in unexplained recurrent pregnancy loss. J. Matern. Fetal Neonatal Med. 2022, 35, 3209–3215. [Google Scholar] [CrossRef]
- Liu, J.; Hao, S.; Chen, X.; Zhao, H.; Du, L.; Ren, H.; Wang, C.; Mao, H. Human placental trophoblast cells contribute to maternal-fetal tolerance through expressing IL-35 and mediating iT(R)35 conversion. Nat. Commun. 2019, 10, 4601. [Google Scholar] [CrossRef] [Green Version]
- Marwood, M.; Visser, K.; Salamonsen, L.A.; Dimitriadis, E. Interleukin-11 and leukemia inhibitory factor regulate the adhesion of endometrial epithelial cells: Implications in fertility regulation. Endocrinology 2009, 150, 2915–2923. [Google Scholar] [CrossRef] [Green Version]
- Paiva, P.; Salamonsen, L.A.; Manuelpillai, U.; Walker, C.; Tapia, A.; Wallace, E.M.; Dimitriadis, E. Interleukin-11 promotes migration, but not proliferation, of human trophoblast cells, implying a role in placentation. Endocrinology 2007, 148, 5566–5572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpovich, N.; Klemmt, P.; Hwang, J.H.; McVeigh, J.E.; Heath, J.K.; Barlow, D.H.; Mardon, H.J. The production of interleukin-11 and decidualization are compromised in endometrial stromal cells derived from patients with infertility. J. Clin. Endocrinol. Metab. 2005, 90, 1607–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, L.; Li, R.; Hartley, L.; Nandurkar, H.H.; Koentgen, F.; Begley, C.G. Infertility in female mice lacking the receptor for interleukin 11 is due to a defective uterine response to implantation. Nat. Med. 1998, 4, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Bilinski, P.; Roopenian, D.; Gossler, A. Maternal IL-11Ralpha function is required for normal decidua and fetoplacental development in mice. Genes Dev. 1998, 12, 2234–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maconochie, N.; Doyle, P.; Prior, S.; Simmons, R. Risk factors for first trimester miscarriage--results from a UK-population-based case-control study. BJOG 2007, 114, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, L.; Kou, Z.; Huo, M.; An, J.; Zhang, X. GnRH agonist treatment regulates IL-6 and IL-11 expression in endometrial stromal cells for patients with HRT regiment in frozen embryo transfer cycles. Reprod. Biol. 2022, 22, 100608. [Google Scholar] [CrossRef]
- Giannubilo, S.R.; Landi, B.; Pozzi, V.; Sartini, D.; Cecati, M.; Stortoni, P.; Corradetti, A.; Saccucci, F.; Tranquilli, A.L.; Emanuelli, M. The involvement of inflammatory cytokines in the pathogenesis of recurrent miscarriage. Cytokine 2012, 58, 50–56. [Google Scholar] [CrossRef]
- Haider, S.; Lackner, A.I.; Dietrich, B.; Kunihs, V.; Haslinger, P.; Meinhardt, G.; Maxian, T.; Saleh, L.; Fiala, C.; Pollheimer, J.; et al. Transforming growth factor-β signaling governs the differentiation program of extravillous trophoblasts in the developing human placenta. Proc. Natl. Acad. Sci. USA 2022, 119, e2120667119. [Google Scholar] [CrossRef]
- Clark, D.A.; Chaouat, G.; Arck, P.C.; Mittruecker, H.W.; Levy, G.A. Cytokine-dependent abortion in CBA x DBA/2 mice is mediated by the procoagulant fgl2 prothrombinase [correction of prothombinase]. J. Immunol. 1998, 160, 545–549. [Google Scholar]
- Knackstedt, M.; Ding, J.W.; Arck, P.C.; Hertwig, K.; Coulam, C.B.; August, C.; Lea, R.; Dudenhausen, J.W.; Gorczynski, R.M.; Levy, G.A.; et al. Activation of the novel prothrombinase, fg12, as a basis for the pregnancy complications spontaneous abortion and pre-eclampsia. Am. J. Reprod. Immunol. 2001, 46, 196–210. [Google Scholar] [CrossRef]
- Drake, B.L.; Head, J.R. Murine trophoblast can be killed by lymphokine-activated killer cells. J. Immunol. 1989, 143, 9–14. [Google Scholar] [PubMed]
- Hoshino, T.; Winkler-Pickett, R.T.; Mason, A.T.; Ortaldo, J.R.; Young, H.A. IL-13 production by NK cells: IL-13-producing NK and T cells are present in vivo in the absence of IFN-gamma. J. Immunol. 1999, 162, 51–59. [Google Scholar] [PubMed]
- Loza, M.J.; Perussia, B. Final steps of natural killer cell maturation: A model for type 1-type 2 differentiation? Nat. Immunol. 2001, 2, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Peritt, D.; Robertson, S.; Gri, G.; Showe, L.; Aste-Amezaga, M.; Trinchieri, G. Differentiation of human NK cells into NK1 and NK2 subsets. J. Immunol. 1998, 161, 5821–5824. [Google Scholar] [PubMed]
- Warren, H.S.; Kinnear, B.F.; Phillips, J.H.; Lanier, L.L. Production of IL-5 by human NK cells and regulation of IL-5 secretion by IL-4, IL-10, and IL-12. J. Immunol. 1995, 154, 5144–5152. [Google Scholar]
- Higuma-Myojo, S.; Sasaki, Y.; Miyazaki, S.; Sakai, M.; Siozaki, A.; Miwa, N.; Saito, S. Cytokine profile of natural killer cells in early human pregnancy. Am. J. Reprod. Immunol. 2005, 54, 21–29. [Google Scholar] [CrossRef]
- Veenstra van Nieuwenhoven, A.L.; Bouman, A.; Moes, H.; Heineman, M.J.; de Leij, L.F.; Santema, J.; Faas, M.M. Cytokine production in natural killer cells and lymphocytes in pregnant women compared with women in the follicular phase of the ovarian cycle. Fertil. Steril. 2002, 77, 1032–1037. [Google Scholar] [CrossRef]
- Borzychowski, A.M.; Croy, B.A.; Chan, W.L.; Redman, C.W.; Sargent, I.L. Changes in systemic type 1 and type 2 immunity in normal pregnancy and pre-eclampsia may be mediated by natural killer cells. Eur. J. Immunol. 2005, 35, 3054–3063. [Google Scholar] [CrossRef]
- Shakhar, K.; Ben-Eliyahu, S.; Loewenthal, R.; Rosenne, E.; Carp, H. Differences in number and activity of peripheral natural killer cells in primary versus secondary recurrent miscarriage. Fertil. Steril. 2003, 80, 368–375. [Google Scholar] [CrossRef]
- Aoki, K.; Kajiura, S.; Matsumoto, Y.; Ogasawara, M.; Okada, S.; Yagami, Y.; Gleicher, N. Preconceptional natural-killer-cell activity as a predictor of miscarriage. Lancet 1995, 345, 1340–1342. [Google Scholar] [CrossRef]
- Yamada, H.; Morikawa, M.; Kato, E.H.; Shimada, S.; Kobashi, G.; Minakami, H. Pre-conceptional natural killer cell activity and percentage as predictors of biochemical pregnancy and spontaneous abortion with normal chromosome karyotype. Am. J. Reprod. Immunol. 2003, 50, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Souza, S.S.; Ferriani, R.A.; Santos, C.M.; Voltarelli, J.C. Immunological evaluation of patients with recurrent abortion. J. Reprod. Immunol. 2002, 56, 111–121. [Google Scholar] [CrossRef] [PubMed]
- King, K.; Smith, S.; Chapman, M.; Sacks, G. Detailed analysis of peripheral blood natural killer (NK) cells in women with recurrent miscarriage. Hum. Reprod. 2010, 25, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, S.; Zarnani, A.H.; Asgarian-Omran, H.; Vahedian-Dargahi, Z.; Eshraghian, M.R.; Akbarzadeh-Pasha, Z.; Arefi, S.; Jeddi-Tehrani, M.; Shokri, F. Comparative analysis of NK cell subsets in menstrual and peripheral blood of patients with unexplained recurrent spontaneous abortion and fertile subjects. J. Reprod. Immunol. 2014, 103, 9–17. [Google Scholar] [CrossRef]
- Karami, N.; Boroujerdnia, M.G.; Nikbakht, R.; Khodadadi, A. Enhancement of peripheral blood CD56(dim) cell and NK cell cytotoxicity in women with recurrent spontaneous abortion or in vitro fertilization failure. J. Reprod. Immunol. 2012, 95, 87–92. [Google Scholar] [CrossRef]
- El-Azzamy, H.; Dambaeva, S.V.; Katukurundage, D.; Salazar Garcia, M.D.; Skariah, A.; Hussein, Y.; Germain, A.; Fernandez, E.; Gilman-Sachs, A.; Beaman, K.D.; et al. Dysregulated uterine natural killer cells and vascular remodeling in women with recurrent pregnancy losses. Am. J. Reprod. Immunol. 2018, 80, e13024. [Google Scholar] [CrossRef]
- Dong, P.; Wen, X.; Liu, J.; Yan, C.Y.; Yuan, J.; Luo, L.R.; Hu, Q.F.; Li, J. Simultaneous detection of decidual Th1/Th2 and NK1/NK2 immunophenotyping in unknown recurrent miscarriage using 8-color flow cytometry with FSC/Vt extended strategy. Biosci. Rep. 2017, 37, BSR20170150. [Google Scholar] [CrossRef]
- Fukui, A.; Funamizu, A.; Fukuhara, R.; Shibahara, H. Expression of natural cytotoxicity receptors and cytokine production on endometrial natural killer cells in women with recurrent pregnancy loss or implantation failure, and the expression of natural cytotoxicity receptors on peripheral blood natural killer cells in pregnant women with a history of recurrent pregnancy loss. J. Obstet. Gynaecol. Res. 2017, 43, 1678–1686. [Google Scholar] [CrossRef] [Green Version]
- Fukui, A.; Kwak-Kim, J.; Ntrivalas, E.; Gilman-Sachs, A.; Lee, S.K.; Beaman, K. Intracellular cytokine expression of peripheral blood natural killer cell subsets in women with recurrent spontaneous abortions and implantation failures. Fertil. Steril. 2008, 89, 157–165. [Google Scholar] [CrossRef]
- Zhu, L.; Aly, M.; Wang, H.; Karakizlis, H.; Weimer, R.; Morath, C.; Kuon, R.J.; Toth, B.; Ekpoom, N.; Opelz, G.; et al. Increased natural killer cell subsets with inhibitory cytokines and inhibitory surface receptors in patients with recurrent miscarriage and decreased or normal subsets in kidney transplant recipients late post-transplant. Clin. Exp. Immunol. 2018, 193, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Kamoi, M.; Fukui, A.; Kwak-Kim, J.; Fuchinoue, K.; Funamizu, A.; Chiba, H.; Yokota, M.; Fukuhara, R.; Mizunuma, H. NK22 Cells in the Uterine Mid-Secretory Endometrium and Peripheral Blood of Women with Recurrent Pregnancy Loss and Unexplained Infertility. Am. J. Reprod. Immunol. 2015, 73, 557–567. [Google Scholar] [CrossRef] [PubMed]
- O’Hern Perfetto, C.; Fan, X.; Dahl, S.; Krieg, S.; Westphal, L.M.; Bunker Lathi, R.; Nayak, N.R. Expression of interleukin-22 in decidua of patients with early pregnancy and unexplained recurrent pregnancy loss. J. Assist. Reprod. Genet. 2015, 32, 977–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zourbas, S.; Dubanchet, S.; Martal, J.; Chaouat, G. Localization of pro-inflammatory (IL-12, IL-15) and anti-inflammatory (IL-11, IL-13) cytokines at the foetomaternal interface during murine pregnancy. Clin. Exp. Immunol. 2001, 126, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Hiby, S.E.; Loke, Y.W.; King, A. Human decidual natural killer cells express the receptor for and respond to the cytokine interleukin 15. Biol. Reprod. 2000, 62, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Ashkar, A.A.; Black, G.P.; Wei, Q.; He, H.; Liang, L.; Head, J.R.; Croy, B.A. Assessment of requirements for IL-15 and IFN regulatory factors in uterine NK cell differentiation and function during pregnancy. J. Immunol. 2003, 171, 2937–2944. [Google Scholar] [CrossRef] [Green Version]
- Wilkens, J.; Male, V.; Ghazal, P.; Forster, T.; Gibson, D.A.; Williams, A.R.; Brito-Mutunayagam, S.L.; Craigon, M.; Lourenco, P.; Cameron, I.T.; et al. Uterine NK cells regulate endometrial bleeding in women and are suppressed by the progesterone receptor modulator asoprisnil. J. Immunol. 2013, 191, 2226–2235. [Google Scholar] [CrossRef] [Green Version]
- Barber, E.M.; Pollard, J.W. The uterine NK cell population requires IL-15 but these cells are not required for pregnancy nor the resolution of a Listeria monocytogenes infection. J. Immunol. 2003, 171, 37–46. [Google Scholar] [CrossRef]
- Murata, H.; Tanaka, S.; Okada, H. Immune Tolerance of the Human Decidua. J. Clin. Med. 2021, 10, 351. [Google Scholar] [CrossRef]
- Chegini, N.; Ma, C.; Roberts, M.; Williams, R.S.; Ripps, B.A. Differential expression of interleukins (IL) IL-13 and IL-15 throughout the menstrual cycle in endometrium of normal fertile women and women with recurrent spontaneous abortion. J. Reprod. Immunol. 2002, 56, 93–110. [Google Scholar] [CrossRef]
- Toth, B.; Haufe, T.; Scholz, C.; Kuhn, C.; Friese, K.; Karamouti, M.; Makrigiannakis, A.; Jeschke, U. Placental interleukin-15 expression in recurrent miscarriage. Am. J. Reprod. Immunol. 2010, 64, 402–410. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Takeda, K.; Tanaka, T.; Ohkusu, K.; Kashiwamura, S.; Okamura, H.; Akira, S.; Nakanishi, K. IL-12 up-regulates IL-18 receptor expression on T cells, Th1 cells, and B cells: Synergism with IL-18 for IFN-gamma production. J. Immunol. 1998, 161, 3400–3407. [Google Scholar] [PubMed]
- Hyodo, Y.; Matsui, K.; Hayashi, N.; Tsutsui, H.; Kashiwamura, S.; Yamauchi, H.; Hiroishi, K.; Takeda, K.; Tagawa, Y.; Iwakura, Y.; et al. IL-18 up-regulates perforin-mediated NK activity without increasing perforin messenger RNA expression by binding to constitutively expressed IL-18 receptor. J. Immunol. 1999, 162, 1662–1668. [Google Scholar] [PubMed]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 regulates both Th1 and Th2 responses. Annu. Rev. Immunol. 2001, 19, 423–474. [Google Scholar] [CrossRef] [PubMed]
- Comba, C.; Bastu, E.; Dural, O.; Yasa, C.; Keskin, G.; Ozsurmeli, M.; Buyru, F.; Serdaroglu, H. Role of inflammatory mediators in patients with recurrent pregnancy loss. Fertil. Steril. 2015, 104, 1467–1474.e1461. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, L.; Wang, C.C.; Huang, J.; Li, T.C. Hypoxia inducible factor and microvessels in peri-implantation endometrium of women with recurrent miscarriage. Fertil. Steril. 2016, 105, 1496–1502.e1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Liu, Y.; Cheung, W.C.; Zhao, Y.; Huang, J.; Chung, J.P.W.; Wang, C.C.; Li, T.C. Increased expression of angiogenic cytokines in CD56+ uterine natural killer cells from women with recurrent miscarriage. Cytokine 2018, 110, 272–276. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, X.; He, G.; Yang, C. Association between TNF, IL1B, IL6, IL10 and IFNG polymorphisms and recurrent miscarriage: A case control study. Reprod. Biol. Endocrinol. 2017, 15, 83. [Google Scholar] [CrossRef]
- Bombell, S.; McGuire, W. Cytokine polymorphisms in women with recurrent pregnancy loss: Meta-analysis. Aust. N. Z. J. Obstet. Gynaecol. 2008, 48, 147–154. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, T.; Wang, Z. Association of tumor necrosis factor-α gene promoter polymorphisms (-308G/A, -238G/A) with recurrent spontaneous abortion: A meta-analysis. Hum. Immunol. 2012, 73, 574–579. [Google Scholar] [CrossRef]
- Li, H.H.; Xu, X.H.; Tong, J.; Zhang, K.Y.; Zhang, C.; Chen, Z.J. Association of TNF-α genetic polymorphisms with recurrent pregnancy loss risk: A systematic review and meta-analysis. Reprod. Biol. Endocrinol. 2016, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Xie, X.; Jia, Y.; Li, S. Maternal genetic polymorphisms and unexplained recurrent miscarriage: A systematic review and meta-analysis. Clin. Genet. 2017, 91, 265–284. [Google Scholar] [CrossRef]
- Zammiti, W.; Mtiraoui, N.; Finan, R.R.; Almawi, W.Y.; Mahjoub, T. Tumor necrosis factor alpha and lymphotoxin alpha haplotypes in idiopathic recurrent pregnancy loss. Fertil. Steril. 2009, 91, 1903–1908. [Google Scholar] [CrossRef]
- Finan, R.R.; Al-Irhayim, Z.; Mustafa, F.E.; Al-Zaman, I.; Mohammed, F.A.; Al-Khateeb, G.M.; Madan, S.; Issa, A.A.; Almawi, W.Y. Tumor necrosis factor-alpha polymorphisms in women with idiopathic recurrent miscarriage. J. Reprod. Immunol. 2010, 84, 186–192. [Google Scholar] [CrossRef]
- Liu, C.; Wang, J.; Zhou, S.; Wang, B.; Ma, X. Association between -238 but not -308 polymorphism of Tumor necrosis factor alpha (TNF-alpha)v and unexplained recurrent spontaneous abortion (URSA) in Chinese population. Reprod. Biol. Endocrinol. 2010, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Prakash, S.; Parveen, F.; Agrawal, S. Association of CTLA-4 and TNF-α polymorphism with recurrent miscarriage among North Indian women. Cytokine 2012, 60, 456–462. [Google Scholar] [CrossRef]
- Alkhuriji, A.F.; Alhimaidi, A.R.; Babay, Z.A.; Wary, A.S. The relationship between cytokine gene polymorphism and unexplained recurrent spontaneous abortion in Saudi females. Saudi Med. J. 2013, 34, 484–489. [Google Scholar]
- Lee, B.E.; Jeon, Y.J.; Shin, J.E.; Kim, J.H.; Choi, D.H.; Jung, Y.W.; Shim, S.H.; Lee, W.S.; Kim, N.K. Tumor necrosis factor-α gene polymorphisms in Korean patients with recurrent spontaneous abortion. Reprod. Sci. 2013, 20, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Babbage, S.J.; Arkwright, P.D.; Vince, G.S.; Perrey, C.; Pravica, V.; Quenby, S.; Bates, M.; Hutchinson, I.V. Cytokine promoter gene polymorphisms and idiopathic recurrent pregnancy loss. J. Reprod. Immunol. 2001, 51, 21–27. [Google Scholar] [CrossRef]
- Daher, S.; Shulzhenko, N.; Morgun, A.; Mattar, R.; Rampim, G.F.; Camano, L.; DeLima, M.G. Associations between cytokine gene polymorphisms and recurrent pregnancy loss. J. Reprod. Immunol. 2003, 58, 69–77. [Google Scholar] [CrossRef]
- Pietrowski, D.; Bettendorf, H.; Keck, C.; Bürkle, B.; Unfried, G.; Riener, E.K.; Hefler, L.A.; Tempfer, C. Lack of association of TNFalpha gene polymorphisms and recurrent pregnancy loss in Caucasian women. J. Reprod. Immunol. 2004, 61, 51–58. [Google Scholar] [CrossRef]
- Prigoshin, N.; Tambutti, M.; Larriba, J.; Gogorza, S.; Testa, R. Cytokine gene polymorphisms in recurrent pregnancy loss of unknown cause. Am. J. Reprod. Immunol. 2004, 52, 36–41. [Google Scholar] [CrossRef]
- Kamali-Sarvestani, E.; Zolghadri, J.; Gharesi-Fard, B.; Sarvari, J. Cytokine gene polymorphisms and susceptibility to recurrent pregnancy loss in Iranian women. J. Reprod. Immunol. 2005, 65, 171–178. [Google Scholar] [CrossRef]
- Pravica, V.; Perrey, C.; Stevens, A.; Lee, J.H.; Hutchinson, I.V. A single nucleotide polymorphism in the first intron of the human IFN-gamma gene: Absolute correlation with a polymorphic CA microsatellite marker of high IFN-gamma production. Hum. Immunol. 2000, 61, 863–866. [Google Scholar] [CrossRef]
- Lin, M.T.; Storer, B.; Martin, P.J.; Tseng, L.H.; Gooley, T.; Chen, P.J.; Hansen, J.A. Relation of an interleukin-10 promoter polymorphism to graft-versus-host disease and survival after hematopoietic-cell transplantation. N. Engl. J. Med. 2003, 349, 2201–2210. [Google Scholar] [CrossRef] [Green Version]
- Temple, S.E.; Lim, E.; Cheong, K.Y.; Almeida, C.A.; Price, P.; Ardlie, K.G.; Waterer, G.W. Alleles carried at positions -819 and -592 of the IL10 promoter affect transcription following stimulation of peripheral blood cells with Streptococcus pneumoniae. Immunogenetics 2003, 55, 629–632. [Google Scholar] [CrossRef]
- Gu, C.; Gong, H.; Zhang, Z.; Yang, Z.; Ma, Y. Association of interleukin-10 gene promoter polymorphisms with recurrent pregnancy loss: A meta-analysis. J. Assist. Reprod. Genet. 2016, 33, 907–917. [Google Scholar] [CrossRef] [Green Version]
- Medica, I.; Ostojic, S.; Pereza, N.; Kastrin, A.; Peterlin, B. Association between genetic polymorphisms in cytokine genes and recurrent miscarriage--a meta-analysis. Reprod. Biomed. Online 2009, 19, 406–414. [Google Scholar] [CrossRef]
- Parveen, F.; Shukla, A.; Agarwal, S. Cytokine gene polymorphisms in northern Indian women with recurrent miscarriages. Fertil. Steril. 2013, 99, 433–440. [Google Scholar] [CrossRef]
- Peng, Z.; Lv, X.; Sun, Y.; Dai, S. Association of Interleukin-10-1082A/G Polymorphism with Idiopathic Recurrent Miscarriage: A Systematic Review and Meta-Analysis. Am. J. Reprod. Immunol. 2016, 75, 162–171. [Google Scholar] [CrossRef]
- Karhukorpi, J.; Laitinen, T.; Karttunen, R.; Tiilikainen, A.S. The functionally important IL-10 promoter polymorphism (-1082G-->A) is not a major genetic regulator in recurrent spontaneous abortions. Mol. Hum. Reprod. 2001, 7, 201–203. [Google Scholar] [CrossRef] [Green Version]
- Zammiti, W.; Mtiraoui, N.; Cochery-Nouvellon, E.; Mahjoub, T.; Almawi, W.Y.; Gris, J.C. Association of -592C/A, -819C/T and -1082A/G interleukin-10 promoter polymorphisms with idiopathic recurrent spontaneous abortion. Mol. Hum. Reprod. 2006, 12, 771–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.O.; Lee, W.S.; Lee, B.E.; Jeon, Y.J.; Kim, Y.R.; Jung, S.H.; Chang, S.W.; Kim, N.K. Interleukin-1beta -511T>C genetic variant contributes to recurrent pregnancy loss risk and peripheral natural killer cell proportion. Fertil. Steril. 2014, 102, 206–212.e205. [Google Scholar] [CrossRef]
- Saijo, Y.; Sata, F.; Yamada, H.; Konodo, T.; Kato, E.H.; Kataoka, S.; Shimada, S.; Morikawa, M.; Minakami, H.; Kishi, R. Interleukin-4 gene polymorphism is not involved in the risk of recurrent pregnancy loss. Am. J. Reprod. Immunol. 2004, 52, 143–146. [Google Scholar] [CrossRef]
- Gremlich, S.; Fratta, S.; Rebellato, E.; Uras, R.; Reymondin, D.; Damnon, F.; Germond, M.; Gerber, S. Interleukin-1 receptor antagonist gene (IL-1RN) polymorphism is a predictive factor of clinical pregnancy after IVF. Hum. Reprod. 2008, 23, 1200–1206. [Google Scholar] [CrossRef] [Green Version]
- Naeimi, S.; Ghiam, A.F.; Mojtahedi, Z.; Dehaghani, A.S.; Amani, D.; Ghaderi, A. Interleukin-18 gene promoter polymorphisms and recurrent spontaneous abortion. Eur. J. Obstet. Gynecol. Reprod. Biol. 2006, 128, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Ostojić, S.; Volk, M.; Medica, I.; Kapović, M.; Meden-Vrtovec, H.; Peterlin, B. Polymorphisms in the interleukin-12/18 genes and recurrent spontaneous abortion. Am. J. Reprod. Immunol. 2007, 58, 403–408. [Google Scholar] [CrossRef]
- Yue, J.; Tong, Y.; Zhou, J.; Liu, Q.; Yang, J. Genetic variant in interleukin-18 is associated with idiopathic recurrent miscarriage in Chinese Han population. Int. J. Mol. Sci. 2015, 16, 4180–4189. [Google Scholar] [CrossRef] [Green Version]
- Al-Khateeb, G.M.; Sater, M.S.; Finan, R.R.; Mustafa, F.E.; Al-Busaidi, A.S.; Al-Sulaiti, M.A.; Almawi, W.Y. Analysis of interleukin-18 promoter polymorphisms and changes in interleukin-18 serum levels underscores the involvement of interleukin-18 in recurrent spontaneous miscarriage. Fertil. Steril. 2011, 96, 921–926. [Google Scholar] [CrossRef]
- Messaoudi, S.; Dandana, M.; Magdoud, K.; Meddeb, S.; Ben Slama, N.; Hizem, S.; Mahjoub, T. Interleukin-18 promoter polymorphisms and risk of idiopathic recurrent pregnancy loss in a Tunisian population. J. Reprod. Immunol. 2012, 93, 109–113. [Google Scholar] [CrossRef]
- Unfried, G.; Böcskör, S.; Endler, G.; Nagele, F.; Huber, J.C.; Tempfer, C.B. A polymorphism of the interleukin-6 gene promoter and idiopathic recurrent miscarriage. Hum. Reprod. 2003, 18, 267–270. [Google Scholar] [CrossRef]
- Saijo, Y.; Sata, F.; Yamada, H.; Kondo, T.; Kato, E.H.; Kishi, R. Single nucleotide polymorphisms in the promoter region of the interleukin-6 gene and the risk of recurrent pregnancy loss in Japanese women. Fertil. Steril. 2004, 81, 374–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirturk, F.; Ates, O.; Gunal, O.; Bozkurt, N.; Aysal, T.; Nacar, M.C. IL-6 gene promoter polymorphisms: Genetic susceptibility to recurrent pregnancy loss. Bratisl. Lek. Listy 2014, 115, 479–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Xu, L.J.; Wang, J.; Xian, M.M.; Liu, M. Association of IL-1β and IL-6 gene polymorphisms with recurrent spontaneous abortion in a Chinese Han population. Int. J. Immunogenet. 2012, 39, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.G.; Simpson, N.A.; Walker, R.G.; Economidou, O.; Shillito, J.; Gooi, H.C.; Duffy, S.R.; Walker, J.J. The carriage of pro-inflammatory cytokine gene polymorphisms in recurrent pregnancy loss. Am. J. Reprod. Immunol. 2001, 45, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Hefler, L.A.; Tempfer, C.B.; Bashford, M.T.; Unfried, G.; Zeillinger, R.; Schneeberger, C.; Koelbl, H.; Nagele, F.; Huber, J.C. Polymorphisms of the angiotensinogen gene, the endothelial nitric oxide synthase gene, and the interleukin-1beta gene promoter in women with idiopathic recurrent miscarriage. Mol. Hum. Reprod. 2002, 8, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linjawi, S.; Li, T.C.; Laird, S.; Blakemore, A. Interleukin-1 receptor antagonist and interleukin-1 beta polymorphisms in women with recurrent miscarriage. Fertil. Steril. 2005, 83, 1549–1552. [Google Scholar] [CrossRef] [PubMed]
- Unfried, G.; Tempfer, C.; Schneeberger, C.; Widmar, B.; Nagele, F.; Huber, J.C. Interleukin 1 receptor antagonist polymorphism in women with idiopathic recurrent miscarriage. Fertil. Steril. 2001, 75, 683–687. [Google Scholar] [CrossRef]
- Wang, Z.C.; Yunis, E.J.; De los Santos, M.J.; Xiao, L.; Anderson, D.J.; Hill, J.A. T helper 1-type immunity to trophoblast antigens in women with a history of recurrent pregnancy loss is associated with polymorphism of the IL1B promoter region. Genes Immun. 2002, 3, 38–42. [Google Scholar] [CrossRef]
- Karhukorpi, J.; Laitinen, T.; Kivelä, H.; Tiilikainen, A.; Hurme, M. IL-1 receptor antagonist gene polymorphism in recurrent spontaneous abortion. J. Reprod. Immunol. 2003, 58, 61–67. [Google Scholar] [CrossRef]
- Daher, S.; Mattar, R.; Gueuvoghlanian-Silva, B.Y.; Torloni, M.R. Genetic polymorphisms and recurrent spontaneous abortions: An overview of current knowledge. Am. J. Reprod. Immunol. 2012, 67, 341–347. [Google Scholar] [CrossRef]
- Shivers, K.Y.; Amador, N.; Abrams, L.; Hunter, D.; Jenab, S.; Quiñones-Jenab, V. Estrogen alters baseline and inflammatory-induced cytokine levels independent from hypothalamic-pituitary-adrenal axis activity. Cytokine 2015, 72, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arck, P.; Hansen, P.J.; Mulac Jericevic, B.; Piccinni, M.P.; Szekeres-Bartho, J. Progesterone during pregnancy: Endocrine-immune cross talk in mammalian species and the role of stress. Am. J. Reprod. Immunol. 2007, 58, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Piccinni, M.P.; Scaletti, C.; Vultaggio, A.; Maggi, E.; Romagnani, S. Defective production of LIF, M-CSF and Th2-type cytokines by T cells at fetomaternal interface is associated with pregnancy loss. J. Reprod. Immunol. 2001, 52, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Raghupathy, R.; Al-Azemi, M. Modulation of Cytokine Production by the Dydrogesterone Metabolite Dihydrodydrogesterone. Am. J. Reprod. Immunol. 2015, 74, 419–426. [Google Scholar] [CrossRef]
- Lissauer, D.; Eldershaw, S.A.; Inman, C.F.; Coomarasamy, A.; Moss, P.A.; Kilby, M.D. Progesterone promotes maternal-fetal tolerance by reducing human maternal T-cell polyfunctionality and inducing a specific cytokine profile. Eur. J. Immunol. 2015, 45, 2858–2872. [Google Scholar] [CrossRef] [Green Version]
- AbdulHussain, G.; Azizieh, F.; Makhseed, M.; Raghupathy, R. Effects of Progesterone, Dydrogesterone and Estrogen on the Production of Th1/Th2/Th17 Cytokines by Lymphocytes from Women with Recurrent Spontaneous Miscarriage. J. Reprod. Immunol. 2020, 140, 103132. [Google Scholar] [CrossRef]
- Devall, A.J.; Papadopoulou, A.; Podesek, M.; Haas, D.M.; Price, M.J.; Coomarasamy, A.; Gallos, I.D. Progestogens for preventing miscarriage: A network meta-analysis. Cochrane Database Syst. Rev 2021, 4, Cd013792. [Google Scholar] [CrossRef]
- Coomarasamy, A.; Truchanowicz, E.G.; Rai, R. Does first trimester progesterone prophylaxis increase the live birth rate in women with unexplained recurrent miscarriages? BMJ 2011, 342, d1914. [Google Scholar] [CrossRef]
- Verma, P.; Verma, R.; Nair, R.R.; Budhwar, S.; Khanna, A.; Agrawal, N.R.; Sinha, R.; Birendra, R.; Rajender, S.; Singh, K. Altered crosstalk of estradiol and progesterone with Myeloid-derived suppressor cells and Th1/Th2 cytokines in early miscarriage is associated with early breakdown of maternal-fetal tolerance. Am. J. Reprod. Immunol. 2019, 81, e13081. [Google Scholar] [CrossRef]
- Matalka, K.Z.; Ali, D.A. Stress-induced versus preovulatory and pregnancy hormonal levels in modulating cytokine production following whole blood stimulation. Neuroimmunomodulation 2005, 12, 366–374. [Google Scholar] [CrossRef]
- Wang, C.; Dehghani, B.; Li, Y.; Kaler, L.J.; Vandenbark, A.A.; Offner, H. Oestrogen modulates experimental autoimmune encephalomyelitis and interleukin-17 production via programmed death 1. Immunology 2009, 126, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, M.D.; Kutteh, W.H.; Purkiss, S.; Librach, C.; Schultz, P.; Houlihan, E.; Liao, C. Intravenous immunoglobulin and idiopathic secondary recurrent miscarriage: A multicentered randomized placebo-controlled trial. Hum. Reprod. 2010, 25, 2203–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleussner, E.; Kamin, G.; Seliger, G.; Rogenhofer, N.; Ebner, S.; Toth, B.; Schenk, M.; Henes, M.; Bohlmann, M.K.; Fischer, T.; et al. Low-molecular-weight heparin for women with unexplained recurrent pregnancy loss: A multicenter trial with a minimization randomization scheme. Ann. Intern. Med. 2015, 162, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Zafardoust, S.; Akhondi, M.M.; Sadeghi, M.R.; Mohammadzadeh, A.; Karimi, A.; Jouhari, S.; Ansaripour, S. Efficacy of Intrauterine Injection of Granulocyte Colony Stimulating Factor (G-CSF) on Treatment of Unexplained Recurrent Miscarriage: A Pilot RCT Study. J. Reprod. Infertil. 2017, 18, 379–385. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Anti-Implantation | |
|---|---|
| IL-6 | [47] |
| IL-17 | [48] |
| IL-23 | [48] |
| Pro-Implantation | |
| TNF-α | [49] |
| IFN-γ | [28] |
| TGF-β1 | [50] |
| IL-1 | [51] |
| IL-1β | [52] |
| IL-10 | [53] |
| IL-11 | [54] |
| IL-22 | [55] |
| IL-33 | [56] |
| Cytokines | Expression | Number of the Experimental Group | Number of the Control Group | Reference |
|---|---|---|---|---|
| TNF-α | Increased | 23 | 24 | [37] |
| IFN-γ | Increased | 23 | 24 | [37] |
| TGF-β | Increased | 36 | 30 | [63] |
| No difference | 32 | 32 | [64] | |
| No difference | 29 | 27 | [65] | |
| IL-33 | Decreased | 142 | 123 | [66] |
| IL-4 | Increased | 19 | 15 | [35] |
| No difference | 32 | 32 | [64] | |
| IL-6 | Decreased | 23 | 24 | [37] |
| IL-10 | Decreased | 23 | 28 | [57] |
| Decreased | 19 | 16 | [36] | |
| Decreased | 23 | 24 | [37] | |
| Decreased | 32 | 32 | [64] | |
| Decreased | 30 | 30 | [67] | |
| IL-17 | Increased | 20 | 20 | [48] |
| IL-22 | Increased | 46 | 28 | [68] |
| IL-23 | Increased | 30 | 30 | [67] |
| Increased | 15 | 15 | [69] | |
| IL-35 | Decreased | 60 | 40 | [70] |
| Decreased | 40 | 120 | [71] |
| Cytokines | Expression | Number of the Experimental Group | Number of the Control Group | Reference |
|---|---|---|---|---|
| TNF-α | Increased | 35 | 40 | [137] |
| IFN-γ | Increased | 10 | 10 | [138] |
| TGF-β | Decreased | 35 | 40 | [137] |
| IL-1β | Increased | 15 | 15 | [31] |
| IL-4 | Decreased | 10 | 10 | [138] |
| IL-6 | Increased | 35 | 40 | [137] |
| Decreased | 9 | 12 | [139] | |
| IL-10 | Decreased | 10 | 10 | [138] |
| IL-11 | Decreased | 16 | 9 | [140] |
| IL-17 | Increased | 15 | 15 | [69] |
| IL-18 | Increased | 15 | 9 | [141] |
| IL-23 | Increased | 15 | 15 | [69] |
| Gene Polymorphism | Authors | Race | Number of Study and Control Groups | Genotype | Test Method | Conclusions | |
|---|---|---|---|---|---|---|---|
| Study Group | Control Group | ||||||
| −238G/A | Zammiti, W. et al., 2009 [212] | Tunisian | 372 vs. 274 | 264/88/20 a | 215/52/7 a | PCR-RFLP | The polymorphism of the −238G/A gene was associated with the occurrence of RPL. |
| Finan, R. et al., 2010 [213] | Bahraini Arabs | 204 vs. 248 | 148/52/4 a | 200/48/0 a | PCR-RFLP | −238G/A variants were independent risk factors for RPL. | |
| Liu, C. et al., 2010 [214] | Chinese | 132 vs. 152 | 128/4/0 a | 135/17/0 a | PCR | A statistical difference was exhibited in −238G/A polymorphism. | |
| Gupta, R. et al., 2012 [215] | Indian | 300 vs. 500 | 121/63/16 a | 154/113/33 a | PCR-RFLP | RPL women tended to carry the G allele. | |
| Alkhuriji, A. et al., 2013 [216] | Saudis | 65 vs. 65 | 57/8/0 a | 55/7/3 a | PCR | NS | |
| Lee, B. et al., 2013 [217] | Korean | 357 vs. 236 | 330/26/1 a | 228/8/0 a | PCR-RFLP | TNF-α −238G > A variants elevated the incidence of RPL. | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 732/41/2 a | 745/57/3 a | PCR-RFLP | NS | |
| −308G/A | Babbage, S. et al., 2001 [218] | Caucasian | 43 vs. 73 | 30/13 b | 56/17 b | PCR | NS |
| Daher, S. et al., 2003 [219] | Brazilian | 48 vs. 108 | 36/12 b | 89/19 b | PCR | NS | |
| Pietrowski, D. et al., 2004 [220] | Caucasian | 168 vs. 212 | 133/33/2 a | 167/41/4 a | PCR | NS | |
| Prigoshin, N. et al., 2004 [221] | Argentinean | 41 vs. 54 | 35/6 b | 49/5 b | PCR-SSP | NS | |
| Kamali-Sarvestani, E. et al., 2005 [222] | Iranian | 139 vs. 143 | 117/14 b | 122/21 b | PCR | NS | |
| Zammiti, W. et al., 2009 [212] | Tunisian | 372 vs. 274 | 319/39/14 a | 222/47/5 a | PCR-RFLP | NS | |
| Finan, W. et al., 2010 [213] | Bahraini Arabs | 204 vs. 248 | 164/32/8 a | 212/32/4 a | PCR-RFLP | NS | |
| Liu, C. et al., 2010 [214] | Chinese | 132 vs. 152 | 110/22/0 a | 138/13/1 a | PCR | NS | |
| Gupta, R. et al., 2012 [215] | Indian | 300 vs. 500 | 229/62/9 a | 425/70/5 a | PCR-RFLP | The A allele was more likely to be present in RPL women. | |
| Alkhuriji, A. et al., 2013 [216] | Saudis | 65 vs. 65 | 33/24/8 a | 47/14/4 a | PCR | The −308G > A gene polymorphisms were associated with RPL. | |
| Lee, B. et al., 2013 [217] | Korean | 357 vs. 236 | 319/36/2 a | 213/21/2 a | PCR- RFLP | NS | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 683/86/6 a | 726/76/3 a | PCR- RFLP | NS | |
| −1031T/C | Finan, R. et al., 2010 [213] | Bahraini Arabs | 204 vs. 248 | 152/36/16 a | 219/29/0 a | PCR-RFLP | The frequency of the −1031C allele in RPL was significantly increased. |
| Lee, B. et al., 2013 [217] | Korean | 357 vs. 236 | 230/115/12 a | 191/45/0 a | PCR-RFLP | TNF-α − 1031T > C variants augmented the chance of experiencing RPL. | |
| −376G/A | Finan, R. et al., 2010 [213] | Bahraini Arabs | 204 vs. 248 | 174/20/10 a | 226/22/0 a | PCR-RFLP | Patients with RPL had a higher incidence of carrying the −376A allele. |
| Gene Polymorphism | Authors | Race | Number of Study and Control Groups | Genotype | Test Method | Conclusions | |
|---|---|---|---|---|---|---|---|
| Study Group | Control Group | ||||||
| −1082A/G | Babbage, S. et al., 2001 [218] | Caucasian | 43 vs. 73 | 8/23/12 a | 20/41/12 a | PCR | NS |
| Karhukorpi, J. et al., 2001 [230] | Finnish | 38 vs. 131 | 13/16/9 a | 44/64/23 a | PCR | NS | |
| Daher, S. et al., 2003 [219] | Brazilian | 43 vs. 104 | 13/19/11 a | 45/43/16 a | PCR | NS | |
| Kamali-Sarvestani, E. et al., 2005 [222] | Iranian | 139 vs. 143 | 62/41/24 a | 62/47/21 a | PCR-RFLP | NS | |
| Zammiti, W. et al., 2006 [231] | Tunisian | 350 vs. 200 | 87/185/72 a | 54/107/39 a | PCR-ASA | NS | |
| Parveen, F. et al., 2013 [228] | Indian | 200 vs. 300 | 86/99/15 a | 180/108/12 a | PCR | The A allele has a significant protective effect. | |
| Kim, J. et al., 2014 [232] | Korean | 385 vs. 232 | 333/50/2 a | 198/34/0 a | PCR-RFLP | NS | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 683/88/4 a | 685/113/7 a | PCR-RFLP | NS | |
| −819C/T | Kamali-Sarvestani, E. et al., 2005 [222] | Iranian | 139 vs. 143 | 77/49/13 a | 61/56/15 a | PCR-RFLP | NS |
| Zammiti, W. et al., 2006 [231] | Tunisian | 350 vs. 200 | 182/120/48 a | 124/57/19 a | PCR-ASA | The −819C/T polymorphisms were genetically associated with RPL. | |
| Parveen, F. et al., 2013 [228] | Indian | 200 vs. 300 | 59/111/30 a | 122/142/36 a | PCR | RPL females tended to own the CT genotype. | |
| −592C/A | Kamali-Sarvestani, E. et al., 2005 [222] | Iranian | 139 vs. 143 | 83/35/14 a | 61/56/15 a | PCR-RFLP | −592C/A polymorphism variants were one of the genetic causes of RPL. |
| Zammiti, W. et al., 2006 [231] | Tunisian | 350 vs. 200 | 206/93/51 a | 134/41/25 a | PCR-ASA | NS | |
| Parveen, F. et al., 2013 [228] | Indian | 200 vs. 300 | 91/79/30 a | 148/116/36 a | PCR | NS | |
| Gene Polymorphism | Authors | Race | Number of Study and Control Groups | Genotype | Test Method | Conclusions | |
|---|---|---|---|---|---|---|---|
| Study Group | Control Group | ||||||
| −607C/A | Naeimi, S. et al., 2006 [235] | Iranian | 102 vs. 103 | 37/23/42 a | 32/17/54 a | PCR | NS |
| Ostojic, S. et al., 2007 [236] | Slovenian | 125 vs. 136 | 43/68/14 a | 41/79/16 a | PCR | NS | |
| Yue, J. et al., 2015 [237] | Chinese | 484 vs. 468 | 87/216/181 a | 79/211/178 a | PCR | NS | |
| −137G/C | Naeimi, S. et al., 2006 [235] | Iranian | 102 vs. 103 | 57/40/5 a | 56/39/8 a | PCR | NS |
| Ostojic, S. et al., 2007 [236] | Slovenian | 125 vs. 136 | 59/54/12 a | 62/63/11 a | PCR | NS | |
| Al-Khateeb, G. et al., 2011 [238] | Bahraini | 282 vs. 283 | 146/98/38 a | 152/113/24 a | PCR | NS | |
| Messaoudi, S. et al., 2012 [239] | Tunisian | 235 vs. 235 | 122/82/31 a | 126/92/19 a | PCR | NS | |
| Yue, J. et al., 2015 [237] | Chinese | 484 vs. 468 | 338/108/38 a | 357/102/9 a | PCR | −137G/C variants had statistical relationship with RPL in additive and recessive genetic models. | |
| −656C/A | Al-Khateeb, G. et al., 2011 [238] | Bahraini | 282 vs. 283 | 80/144/58 a | 140/119/30 a | PCR | −656C/A variants were related to RPL. |
| Messaoudi, S. et al., 2012 [239] | Tunisian | 235 vs. 235 | 66/120/49 a | 114/97/24 a | PCR | The genotype frequency of −656C/A was significantly correlated with the occurrence of RPL. | |
| −119A/C | Al-Khateeb, G. et al., 2011 [238] | Bahraini | 282 vs. 283 | 157/99/26 a | 155/109/25 a | PCR | NS |
| Messaoudi, S. et al., 2012 [239] | Tunisian | 235 vs. 235 | 132/82/21 a | 127/89/19 a | PCR | NS | |
| −105G/A | Al-Khateeb, G. et al., 2011 [238] | Bahraini | 282 vs. 283 | 98/94/90 a | 146/110/33 a | PCR | −105G/A variant was prominently associated with RPL. |
| Messaoudi, S. et al., 2012 [239] | Tunisia | 235 vs. 235 | 82/78/75 a | 120/89/26 a | PCR | The genotype frequency of −105G/A was significantly different between the two groups. | |
| Yue, J. et al., 2015 [237] | Chinese | 484 vs. 468 | 349/129/6 a | 332/128/8 a | PCR | NS | |
| Gene Polymorphism | Authors | Race | Number of Study and Control Groups | Genotype | Test Method | Conclusions | |
|---|---|---|---|---|---|---|---|
| Study Group | Control Group | ||||||
| −174G/C | Unfried, G. et al., 2003 [240] | White Middle- European Caucasian women | 161 vs. 124 | 66/72/23 a | 43/58/23 a | PCR | NS |
| Daher, S. et al., 2003 [219] | Brazilian | 44 vs. 108 | 39/5 b | 99/9 b | PCR | NS | |
| Prigoshin, N et al., 2004 [221] | Argentinean | 38 vs. 54 | 35/3 b | 49/5 b | PCR-SSP | NS | |
| Saijo, Y. et al., 2004 [241] | Japanese | 76 vs. 93 | 76/0 c | 93/0 c | PCR | NS | |
| Demirturk, F. et al., 2014 [242] | Turkish | 113 vs. 113 | 72/36/5 a | 100/11/2 a | PCR-RFLP | −174G/C polymorphisms had a relationship with an elevated incidence of RPL. | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 484/248/43 a | 463/291/51 a | PCR- RFLP | NS | |
| −634C/G | Saijo, Y. et al., 2004 [241] | Japanese | 76 vs. 93 | 58/18 c | 56/37 c | PCR | Women with the G allele were more likely to develop RPL than those with the wild-type allele C. |
| Ma, X. et al., 2012 [243] | Chinese | 162 vs. 156 | 116/46/0 a | 93/52/11 a | PCR-RFLP | The distributions of the GG genotype and G allele were significantly decreased in RPL. | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 554/197/24 a | 478/277/50 a | PCR- RFLP | Women with CG and GG genotypes were less likely to develop RPL. | |
| −572G/C | Demirturk, F. et al., 2014 [242] | Turkish | 113 vs. 113 | 81/28/4 a | 88/21/4 a | PCR-RFLP | NS |
| −597G/A | Demirturk, F. et al., 2014 [242] | Turkish | 113 vs. 113 | 96/16/1 a | 87/26/0 a | PCR-RFLP | NS |
| −1363G/T | Demirturk, F. et al., 2014 [242] | Turkish | 113 vs. 113 | 95/18/0 a | 94/19/0 a | PCR-RFLP | NS |
| −2954G/C | Demirturk, F. et al., 2014 [242] | Turkish | 113 vs. 113 | 107/6/0 a | 112/1/0 a | PCR-RFLP | −2954G/C polymorphism variants were related to an elevated risk of RPL. |
| Gene Polymorphism | Authors | Race | Number of Study and Control Groups | Genotype | Test Method | Conclusions | |
|---|---|---|---|---|---|---|---|
| Study Group | Control Group | ||||||
| IL-1β + 3954C→T on exon 5 | Hefler, L. et al., 2001 [83] | Caucasian | 131 vs. 68 | 79/46/6 a | 47/16/5 a | PCR | NS |
| Reid, J. et al., 2001 [244] | British | 17 vs. 43 | 11/6/0 b | 26/14/3 b | PCR | NS | |
| Ma, X. et al., 2012 [243] | Chinese | 162 vs. 156 | 124/38/0 c | 130/26/0 c | PCR-RFLP | NS | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 602/168/5 c | 632/166/7 c | PCR-RFLP | NS | |
| IL-1β − 511C/T | Hefler, L. et al., 2002 [245] | Caucasian | 130 vs. 67 | 29/90/11 c | 20/38/9 c | PCR | NS |
| Linjawi, S. et al., 2005 [246] | British | 206 vs. 224 | 69/117/20 c | 85/110/29 c | PCR | NS | |
| Ma, X. et al., 2012 [243] | Chinese | 162 vs. 156 | 38/84/40 c | 46/84/26 c | PCR-RFLP | NS | |
| Kim, J. et al., 2014 [232] | Korean | 385 vs. 232 | 96/190/99 c | 39/120/73 c | PCR–RFLP | −511C > T polymorphism was relevant to RPL. | |
| Ma, J. et al., 2017 [207] | Chinese | 775 vs. 805 | 178/384/213 c | 156/392/257 c | PCR-RFLP | NS | |
| IL-1 RA | Unfried, G. et al., 2001 [247] | Austrian | 105 vs. 91 | 0.9/0.34/0.5 d | 0.97/0.11/0.5 d | PCR | Allele 2 was a genetic risk factor for RPL. |
| IL-1 RN | Wang, Z. et al., 2002 [248] | Caucasian | 118 vs. 60 | 88/26/2/2 e | 39/20/1/0 e | PCR-RFLP | NS |
| Karhukorpi, J. et al., 2003 [249] | Finnish | 37 vs. 800 | 12/19/2/4 f | 374/343/66/17 f | PCR | The frequency of the IL-1RN*3 allele was significantly increased in RPL patients. | |
| Linjawi, S. et al., 2005 [246] | British | 206 vs. 259 | 12/79/115 g | 17/92/150 g | PCR | NS | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Tian, Y.; Zheng, L.; Luu, T.; Kwak-Kim, J. The Update Immune-Regulatory Role of Pro- and Anti-Inflammatory Cytokines in Recurrent Pregnancy Losses. Int. J. Mol. Sci. 2023, 24, 132. https://doi.org/10.3390/ijms24010132
Yang X, Tian Y, Zheng L, Luu T, Kwak-Kim J. The Update Immune-Regulatory Role of Pro- and Anti-Inflammatory Cytokines in Recurrent Pregnancy Losses. International Journal of Molecular Sciences. 2023; 24(1):132. https://doi.org/10.3390/ijms24010132
Chicago/Turabian StyleYang, Xiuhua, Yingying Tian, Linlin Zheng, Thanh Luu, and Joanne Kwak-Kim. 2023. "The Update Immune-Regulatory Role of Pro- and Anti-Inflammatory Cytokines in Recurrent Pregnancy Losses" International Journal of Molecular Sciences 24, no. 1: 132. https://doi.org/10.3390/ijms24010132