
Genome-Wide Identification of Cassava Glyoxalase I Genes and the Potential Function of MeGLYⅠ-13 in Iron Toxicity Tolerance

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
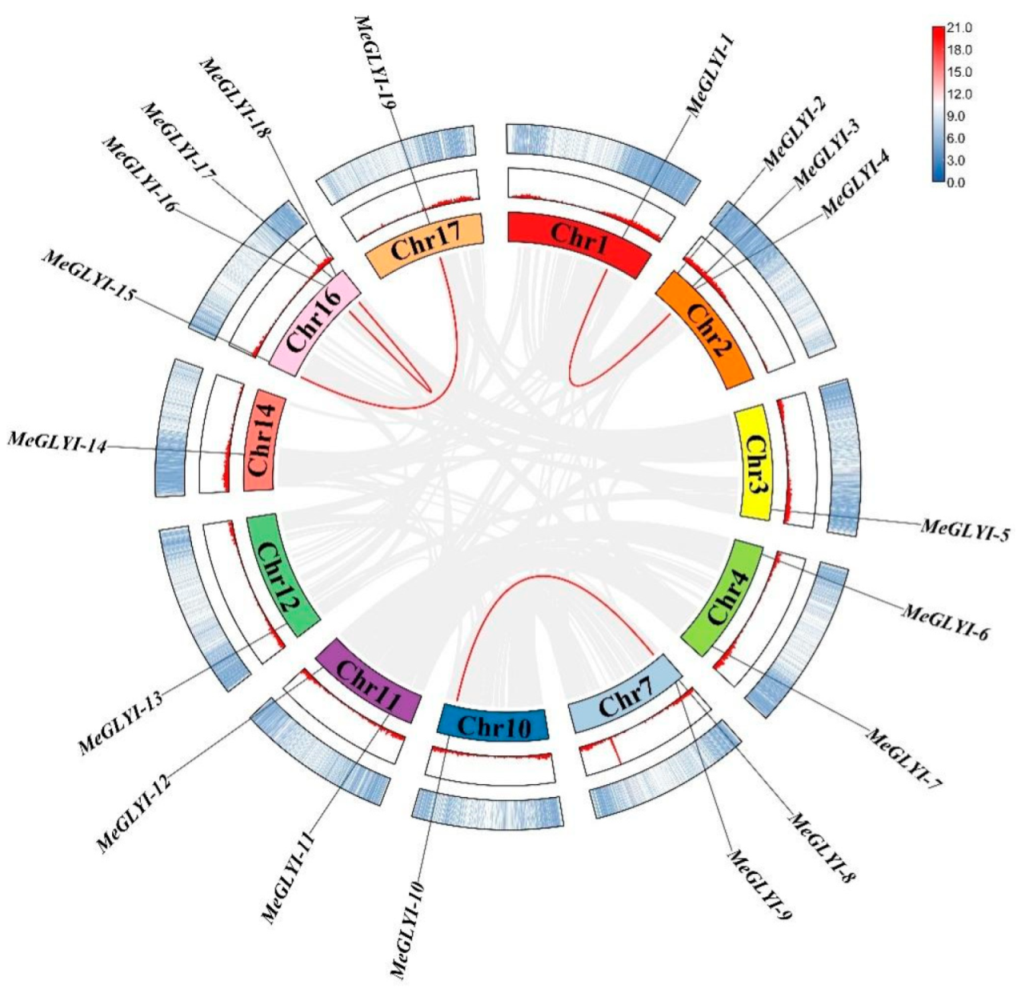
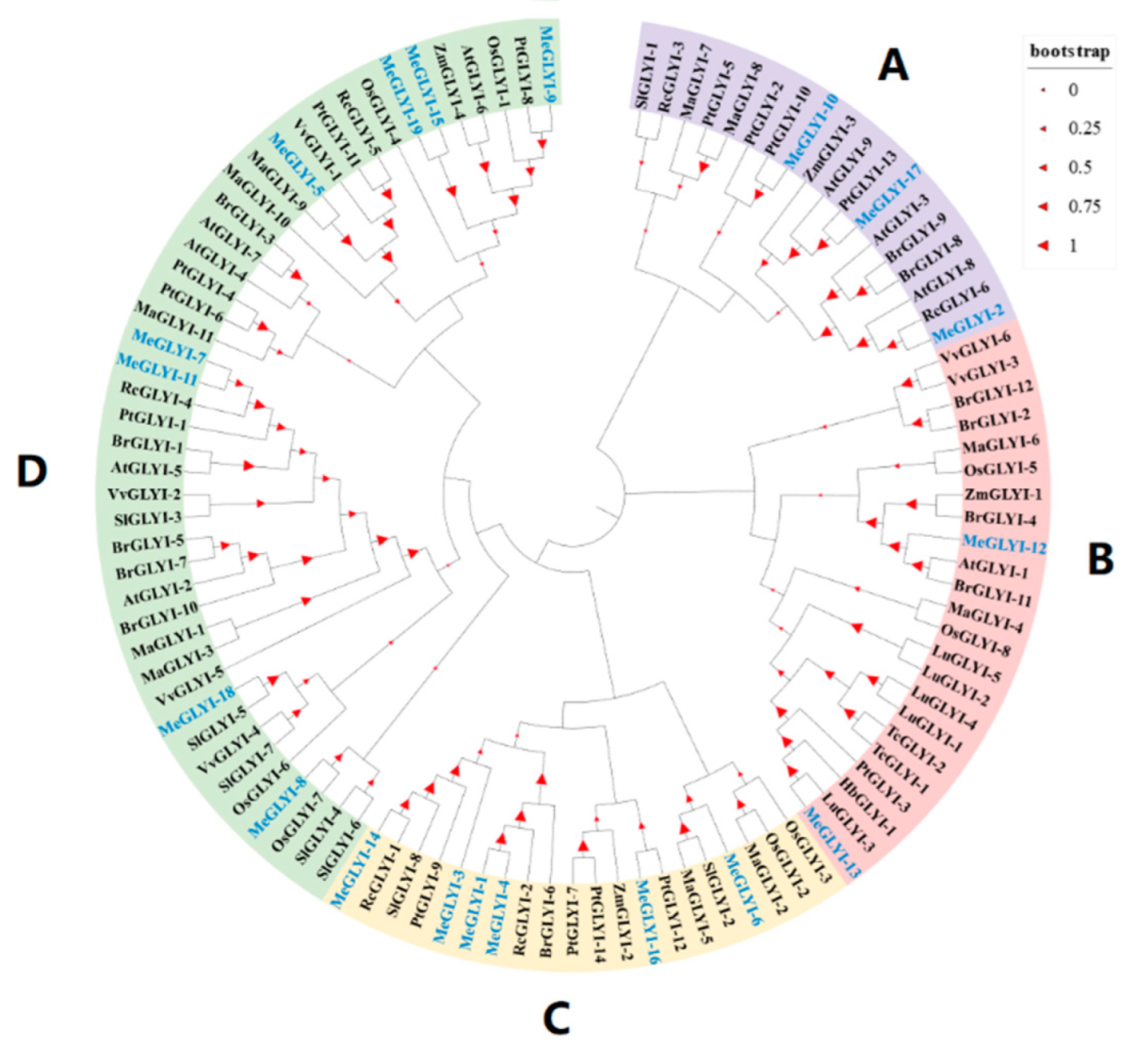
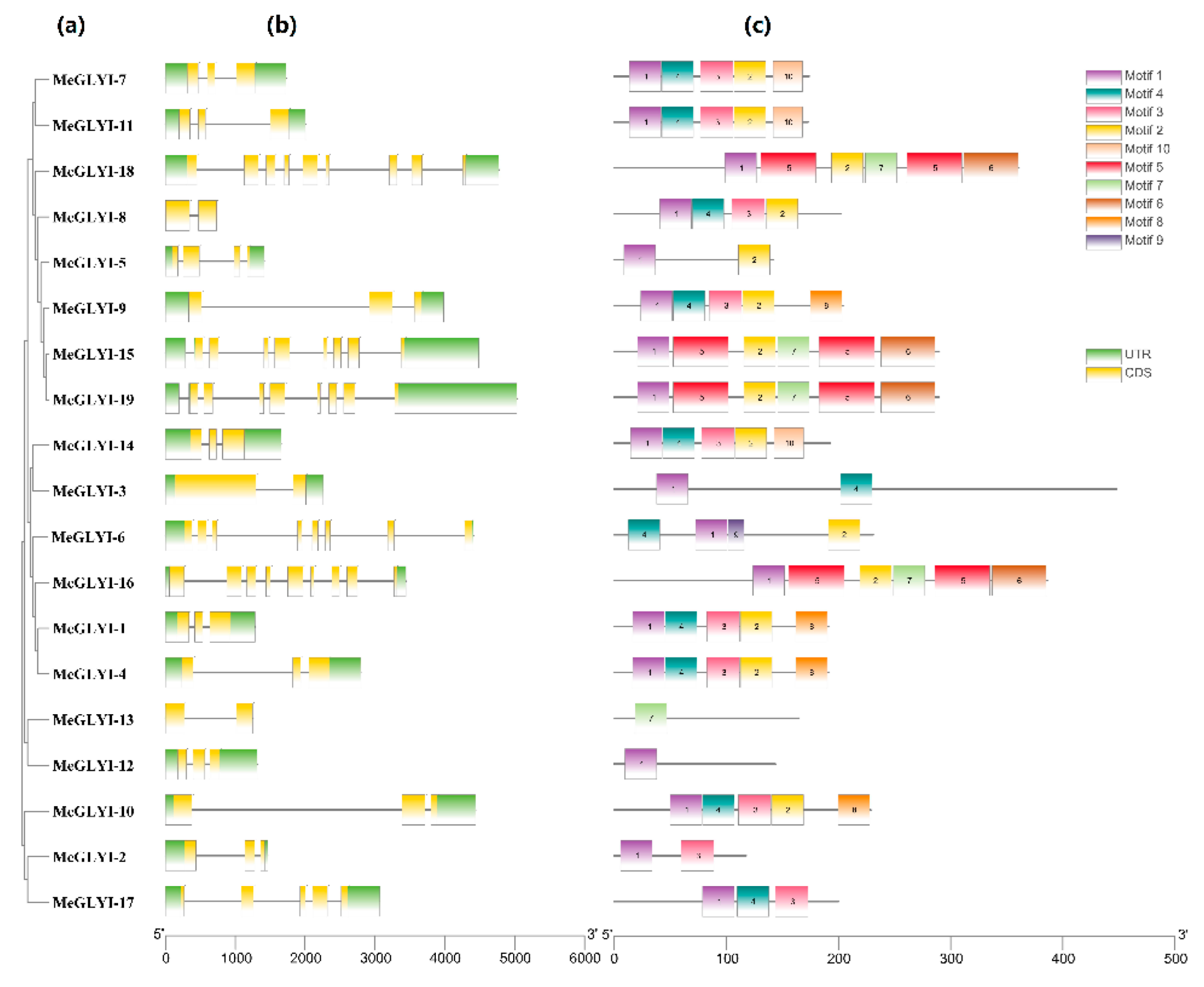
2.1. Identification of 19 GLYIs in M. esculenta
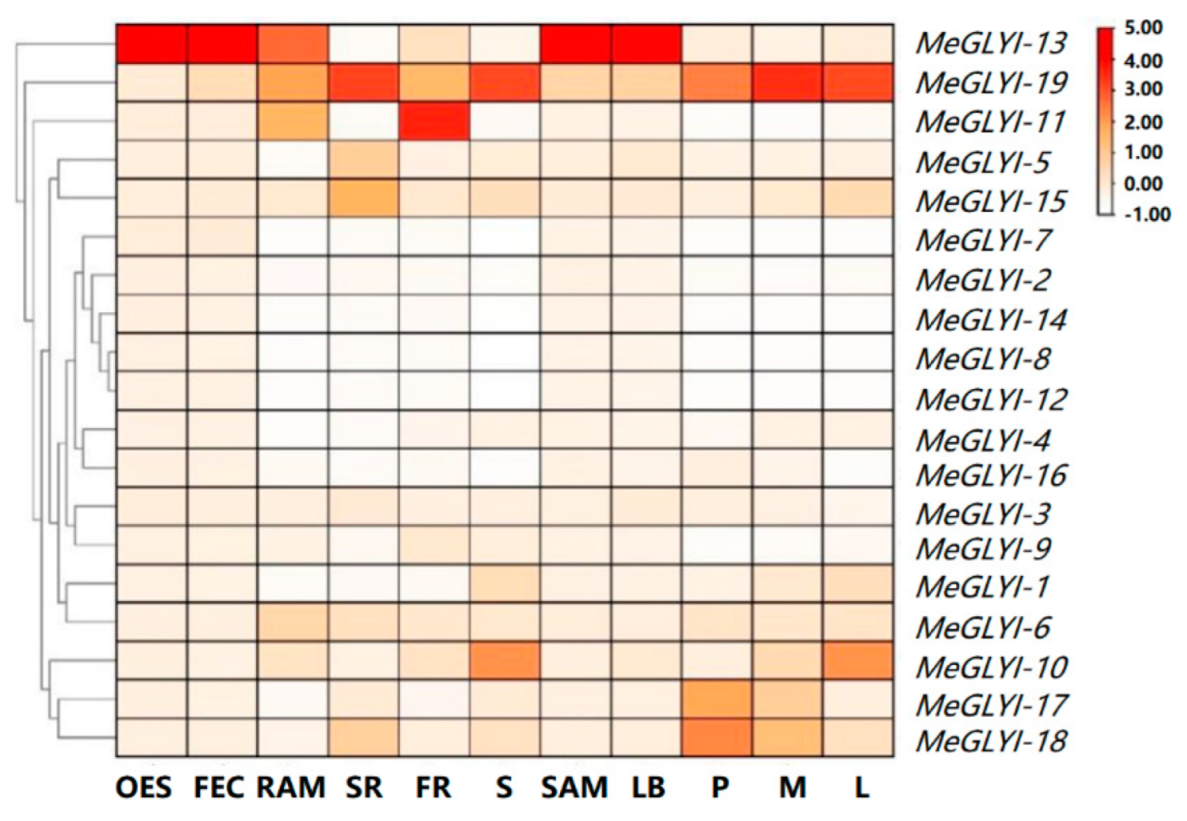
2.2. Difference in the Expression of the MeGLYI Genes in Different Tissues/Organs of Cassava
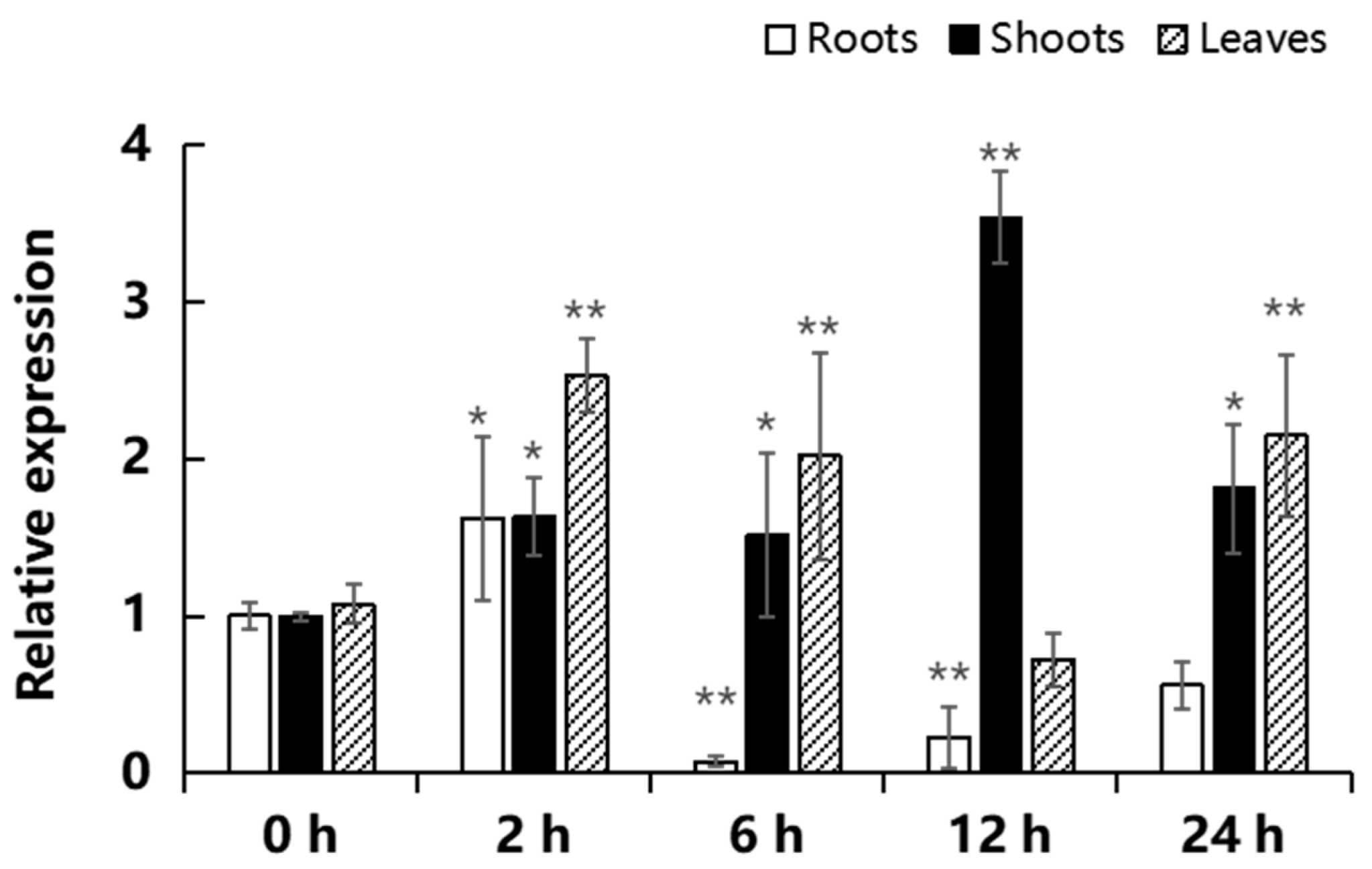
2.3. Expression of MeGLYI-13 in Response to Iron Stress
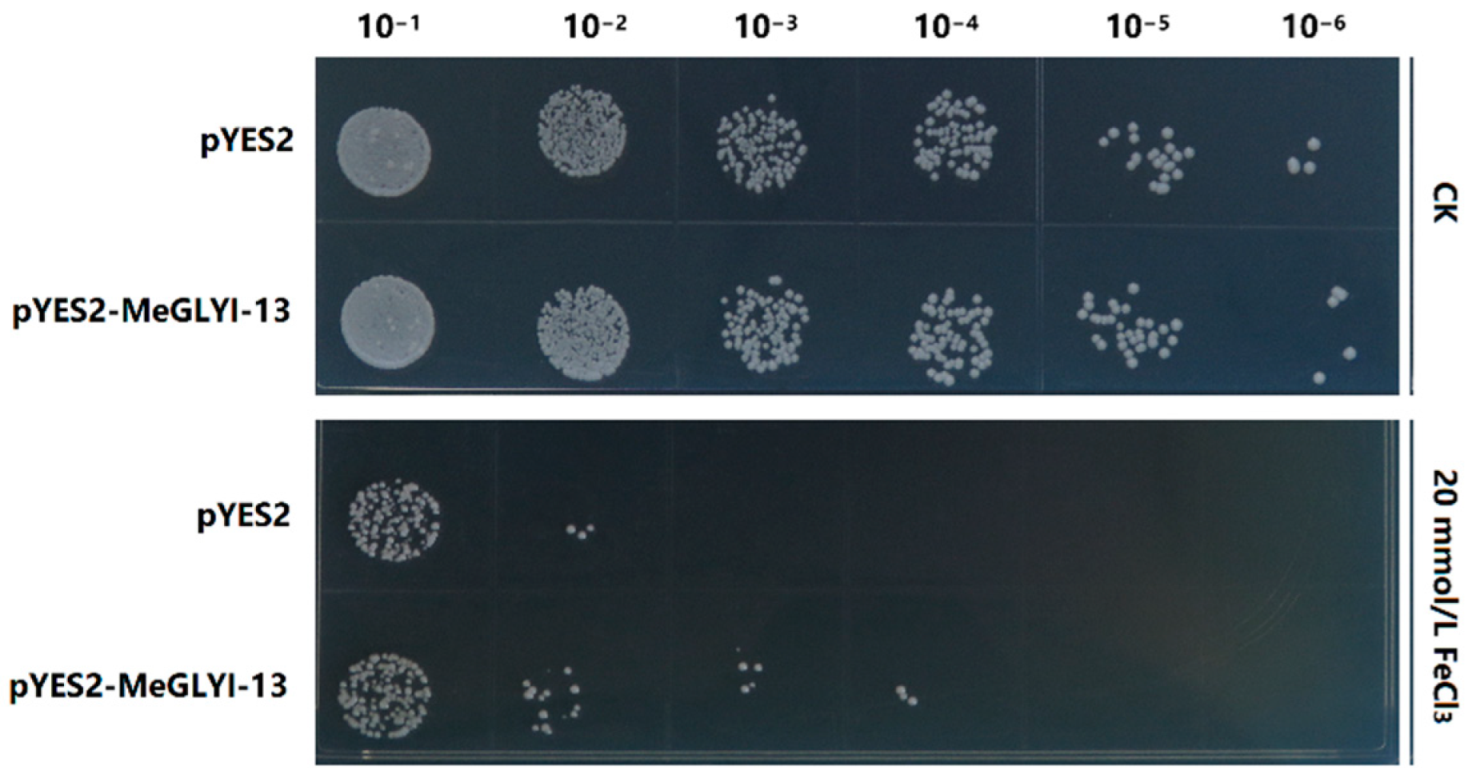
2.4. Overexpression of MeGLYI-13 in Yeast Enhanced the Tolerance to Iron Stress
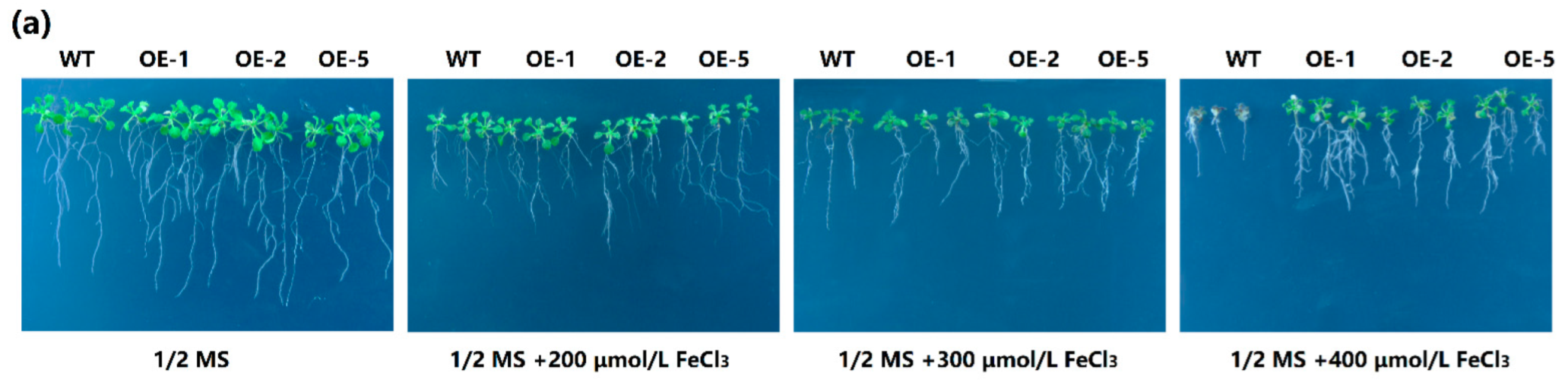
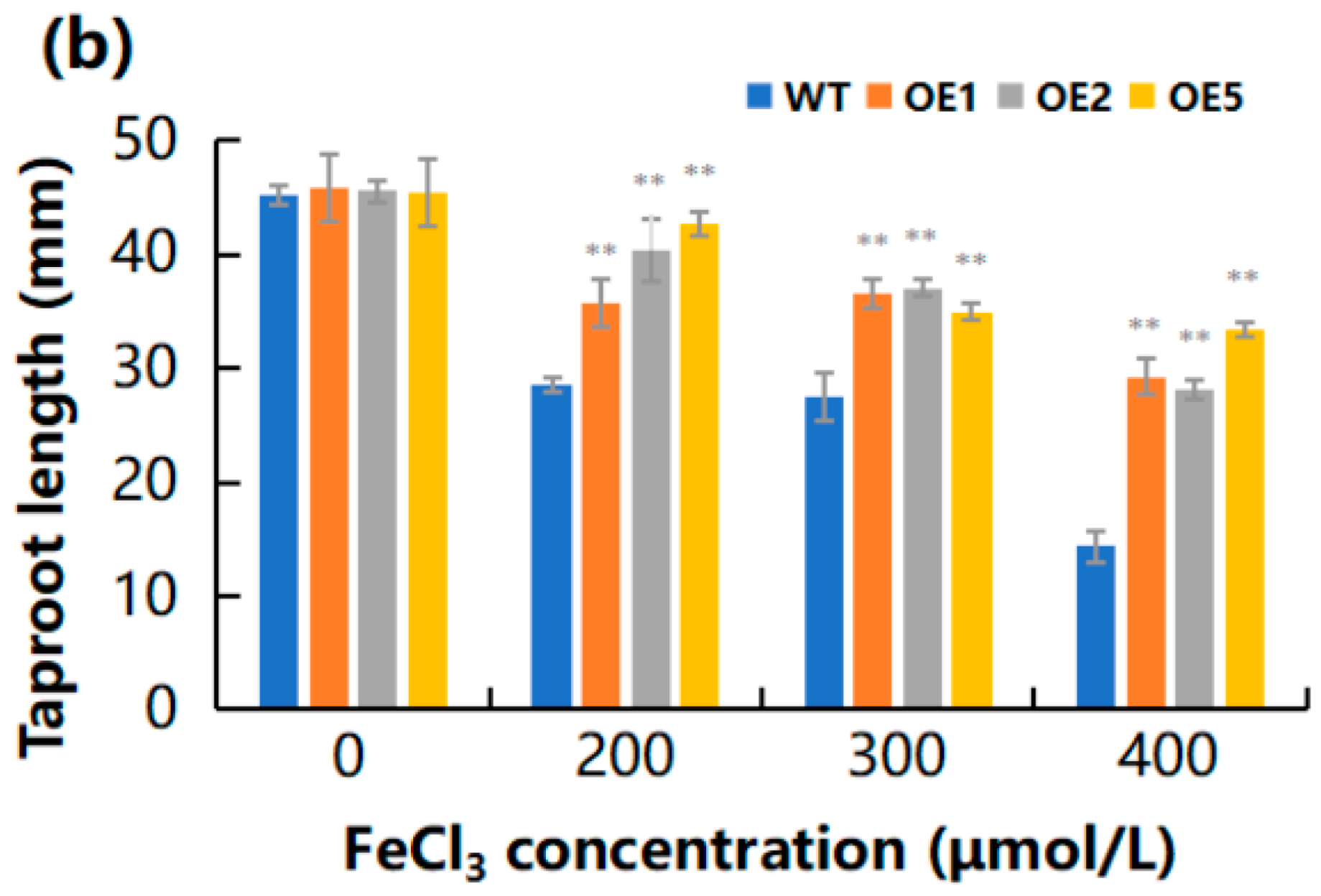
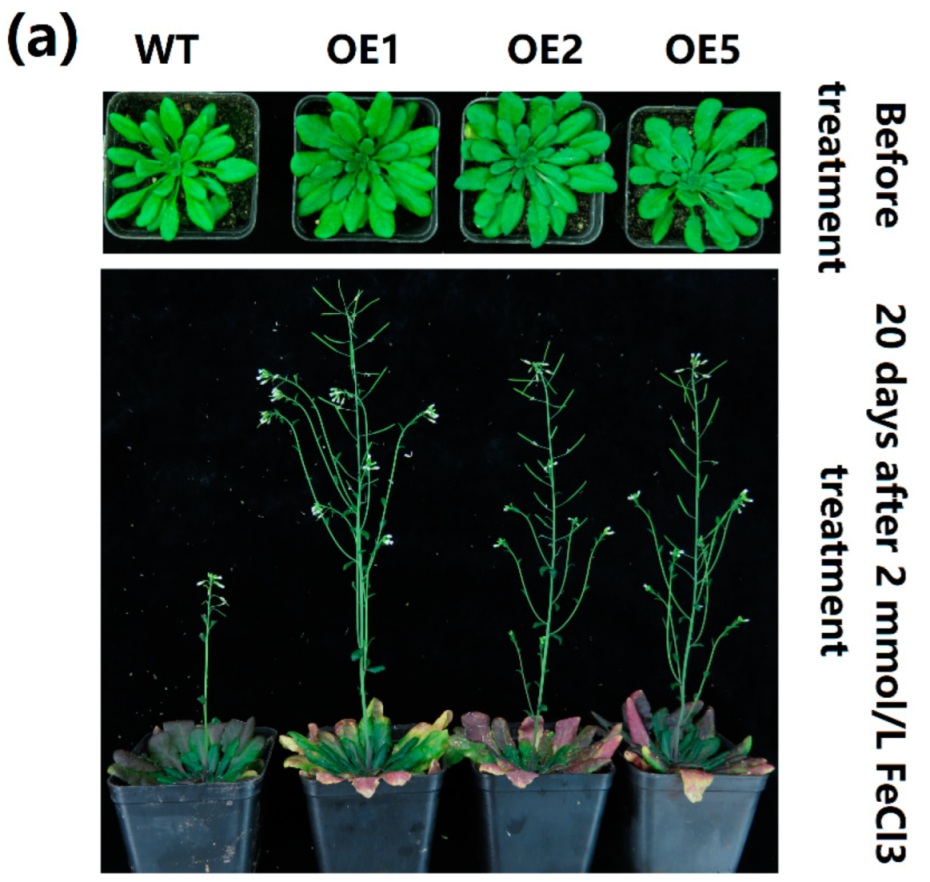
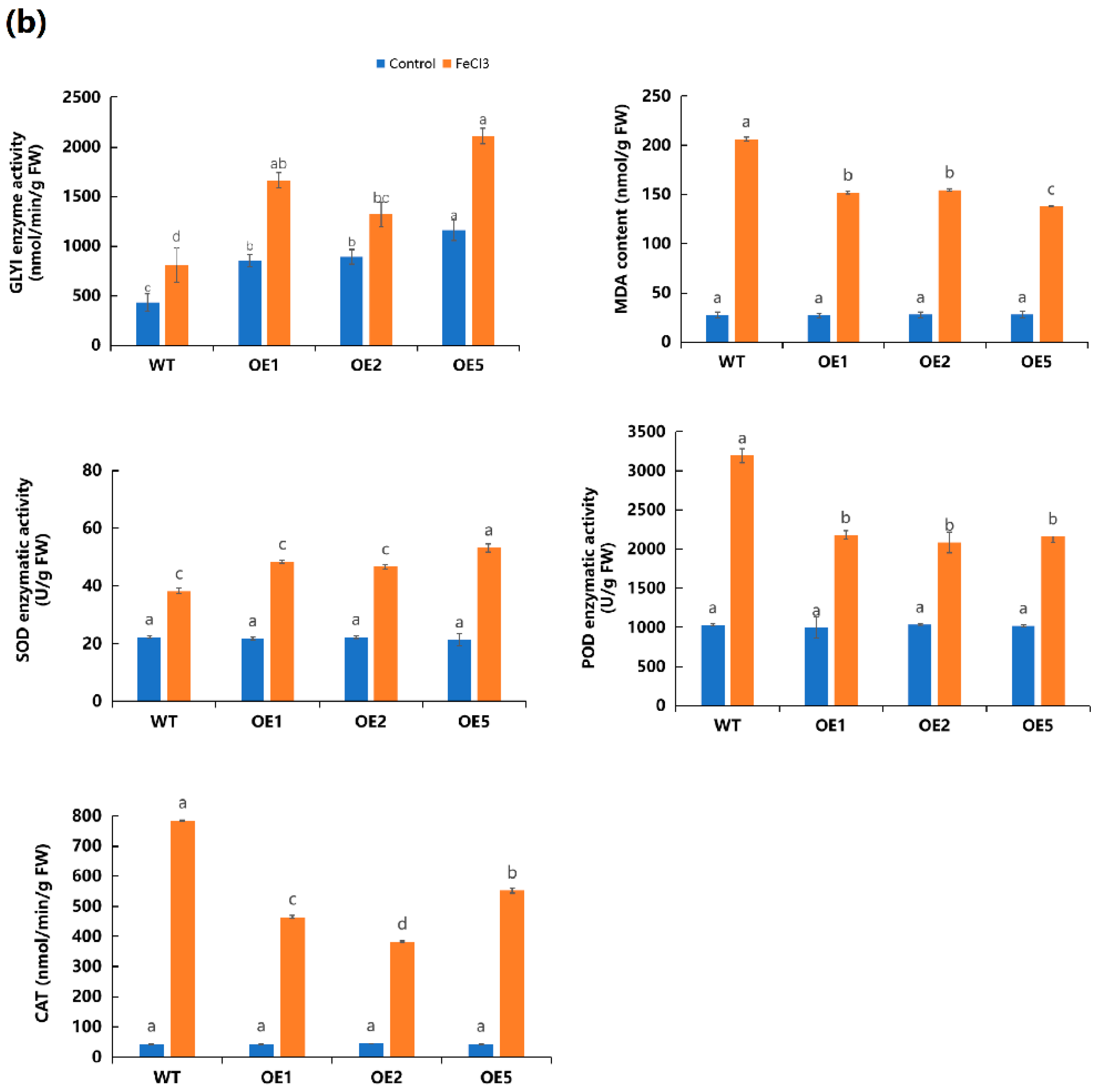
2.5. Overexpression of MeGLYI-13 in Arabidopsis Raised the Tolerance to Iron Stress
3. Discussion
4. Materials and Methods
4.1. Identification and Analysis of GLYI Genes in Cassava
4.2. Expression Analysis of MeGLYIs in Different Tissues and Organs
4.3. Expression Analysis of MeGLYI-13 in Response to Iron Stress
4.4. Cloning of MeGLYI-13 and Functional Analysis in Yeast and Arabidopsis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singla-Pareek, S.L.; Kaur, C.; Kumar, B.; Pareek, A.; Sopory, S.K. Reassessing plant glyoxalases: Large family and expanding functions. New Phytol. 2020, 227, 714–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Cheng, X.; Wang, Y.; Yin, X.; Li, Z.; Liu, R.; Liu, G.; Wang, Y.; Xu, Y. Genome-wide analysis of glyoxalase-like gene families in grape (Vitis vinifera L.) and their expression profiling in response to downy mildew infection. BMC Genom. 2019, 20, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, G.; Xiao, X.; Wang, N.; Zhang, F.; Gao, G.; Xu, K.; Chen, B.; Qiao, J.; Wu, X. Genome-wide analysis and expression profiles of glyoxalase gene families in Chinese cabbage (Brassica rapa L.). PLoS ONE 2018, 13, e0191159. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Islam, T. Genome-wide analysis and expression profiling of glyoxalase gene families in soybean (Glycine max) indicate their development and abiotic stress specific response. BMC Plant Biol. 2016, 16, 87. [Google Scholar] [CrossRef] [Green Version]
- Mustafiz, A.; Singh, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Genome-wide analysis of rice and Arabidopsis identifies two glyoxalase genes that are highly expressed in abiotic stresses. Funct. Integr. Genom. 2011, 11, 293–305. [Google Scholar] [CrossRef]
- Kaur, C.; Tripathi, A.K.; Nutan, K.K.; Sharma, S.; Ghosh, A.; Tripathi, J.K.; Pareek, A.; Singla-Pareek, S.L.; Sopory, S.K. A nuclear-localized rice glyoxalase I enzyme, OsGLYI-8, functions in the detoxification of methylglyoxal in the nucleus. Plant J. 2017, 89, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Li, H.; Thapa, S.; Reddy Sangireddy, S.; Pei, X.; Liu, W.; Jiang, Y.; Yang, S.; Hui, D.; Bhatti, S.; et al. Al-induced proteomics changes in tomato plants over-expressing a glyoxalase I gene. Hortic. Res. 2020, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankaranarayanan, S.; Jamshed, M.; Kumar, A.; Skori, L.; Scandola, S.; Wang, T.; Spiegel, D.; Samuel, M.A. Glyoxalase Goes Green: The Expanding Roles of Glyoxalase in Plants. Int. J. Mol. Sci. 2017, 18, 898. [Google Scholar] [CrossRef]
- Jana, G.A.; Yaish, M.W. Functional characterization of the Glyoxalase-I (PdGLX1) gene family in date palm under abiotic stresses. Plant Signal. Behav. 2020, 15, 1811527. [Google Scholar] [CrossRef]
- Zhou, S.; Sauve, R.; Thannhauser, T.W. Proteome changes induced by aluminium stress in tomato roots. J. Exp. Bot. 2009, 60, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Xu, J.; Shi, J.; Li, H.; Li, B. Molecular cloning and characterization of a novel glyoxalase I gene TaGly I in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2010, 37, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Hafeez, M.B.; Shaukat, K.; Wahid, A.; Hasanuzzaman, M. Fe toxicity in plants: Impacts and remediation. Physiol. Plant 2021, 173, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Onyango, D.A.; Entila, F.; Dida, M.M.; Ismail, A.M.; Drame, K.N. Mechanistic understanding of iron toxicity tolerance in contrasting rice varieties from Africa: 1. Morpho-physiological and biochemical responses. Funct. Plant Biol. 2018, 46, 93–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, S.; Pavia, I.; Carvalho, A.; Moutinho-Pereira, J.; Correia, C.; Lima-Brito, J. Seed priming with iron and zinc in bread wheat: Effects in germination, mitosis and grain yield. Protoplasma 2018, 255, 1179–1194. [Google Scholar] [CrossRef] [PubMed]
- Nozoe, T.; Tachibana, M.; Uchino, A.; Yokogami, N. Effects of ferrous iron (Fe) on the germination and root elongation of paddy rice and weeds. Weed Biol. Manag. 2009, 9, 20–26. [Google Scholar] [CrossRef]
- Zhang, L.; Li, G.; Wang, M.; Di, D.; Sun, L.; Kronzucker, H.J.; Shi, W. Excess iron stress reduces root tip zone growth through nitric oxide-mediated repression of potassium homeostasis in Arabidopsis. New Phytol. 2018, 219, 259–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audebert, A.; Fofana, M. Rice Yield Gap due to Iron Toxicity in West Africa. J. Agron. Crop. Sci. 2009, 195, 66–76. [Google Scholar] [CrossRef]
- Becker, M.; Asch, F. Iron toxicity in rice—Conditions and management concepts. J. Plant Nutr. Soil Sci. 2005, 168, 558–573. [Google Scholar] [CrossRef]
- Onyango, D.A.; Entila, F.; Egdane, J.; Pacleb, M.; Katimbang, M.L.; Dida, M.M.; Ismail, A.M.; Drame, K.N. Mechanistic understanding of iron toxicity tolerance in contrasting rice varieties from Africa: 2. Root oxidation ability and oxidative stress control. Funct. Plant Biol. 2020, 47, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Aung, M.S.; Masuda, H.; Kobayashi, T.; Nishizawa, N.K.J.S.S.; Nutrition, P. Physiological and transcriptomic analysis of responses to different levels of iron excess stress in various rice tissues. Soil Sci. Plant Nutr. 2018, 64, 370–385. [Google Scholar] [CrossRef]
- Aung, M.S.; Kobayashi, T.; Masuda, H.; Nishizawa, N.K. Rice HRZ ubiquitin ligases are crucial for the response to excess iron. Physiol Plant 2018, 163, 282–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathima, A.A.; Sanitha, M.; Tripathi, L.; Muiruri, S. Cassava (Manihot esculenta) dual use for food and bioenergy: A review. Food Energy Secur. 2022, e380. [Google Scholar] [CrossRef]
- Okogbenin, E.; Setter, T.L.; Ferguson, M.; Mutegi, R.; Ceballos, H.; Olasanmi, B.; Fregene, M. Phenotypic approaches to drought in cassava: Review. Front. Physiol. 2013, 4, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.C.; Mutka, A.M.; Hummel, A.W.; Berry, J.; Chauhan, R.D.; Vijayaraghavan, A.; Taylor, N.J.; Voytas, D.F.; Chitwood, D.H.; Bart, R.S. Gene expression atlas for the food security crop cassava. New Phytol. 2017, 213, 1632–1641. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Chen, J.; Chang, J.; Xia, B. Using bioenergy crop cassava (Manihot esculenta) for reclamation of heavily metal-contaminated land. Int. J. Phytoremed. 2020, 22, 1313–1320. [Google Scholar] [CrossRef]
- Ghosh, A. Genome-Wide Identification of Glyoxalase Genes in Medicago truncatula and Their Expression Profiling in Response to Various Developmental and Environmental Stimuli. Front. Plant Sci. 2017, 8, 836. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Mukherjee, S.; Kumar, R.; Alansi, S.; Shah, A.A.; Kalaji, H.M.; Javed, T.; Raza, A. Potassium and melatonin-mediated regulation of fructose-1,6-bisphosphatase (FBPase) and sedoheptulose-1,7-bisphosphatase (SBPase) activity improve photosynthetic efficiency, carbon assimilation and modulate glyoxalase system accompanying tolerance to cadmium stress in tomato seedlings. Plant Physiol. Biochem. 2022, 171, 49–65. [Google Scholar]
- Wu, Q.; Gao, S.; Pan, Y.B.; Su, Y.; Grisham, M.P.; Guo, J.; Xu, L.; Que, Y. Heterologous expression of a Glyoxalase I gene from sugarcane confers tolerance to several environmental stresses in bacteria. PeerJ 2018, 6, e5873. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Xiong, F.; Yu, X.; Gong, X.; Luo, J.; Jiang, Y.; Kuang, H.; Gao, B.; Niu, X.; Liu, Y. Overexpression of a glyoxalase gene, OsGly I, improves abiotic stress tolerance and grain yield in rice (Oryza sativa L.). Plant Physiol. Biochem. 2016, 109, 62–71. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, F.; Li, R.; Zhou, Y.; Wang, S.; Zhou, Q.; Ding, Z.; Yao, Y.; Liu, J.; Wang, Y.; Hu, X.; et al. Genome-Wide Identification of Cassava Glyoxalase I Genes and the Potential Function of MeGLYⅠ-13 in Iron Toxicity Tolerance. Int. J. Mol. Sci. 2022, 23, 5212. https://doi.org/10.3390/ijms23095212
Tang F, Li R, Zhou Y, Wang S, Zhou Q, Ding Z, Yao Y, Liu J, Wang Y, Hu X, et al. Genome-Wide Identification of Cassava Glyoxalase I Genes and the Potential Function of MeGLYⅠ-13 in Iron Toxicity Tolerance. International Journal of Molecular Sciences. 2022; 23(9):5212. https://doi.org/10.3390/ijms23095212
Chicago/Turabian StyleTang, Fenlian, Ruimei Li, Yangjiao Zhou, Shijia Wang, Qin Zhou, Zhongping Ding, Yuan Yao, Jiao Liu, Yajie Wang, Xinwen Hu, and et al. 2022. "Genome-Wide Identification of Cassava Glyoxalase I Genes and the Potential Function of MeGLYⅠ-13 in Iron Toxicity Tolerance" International Journal of Molecular Sciences 23, no. 9: 5212. https://doi.org/10.3390/ijms23095212