
Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
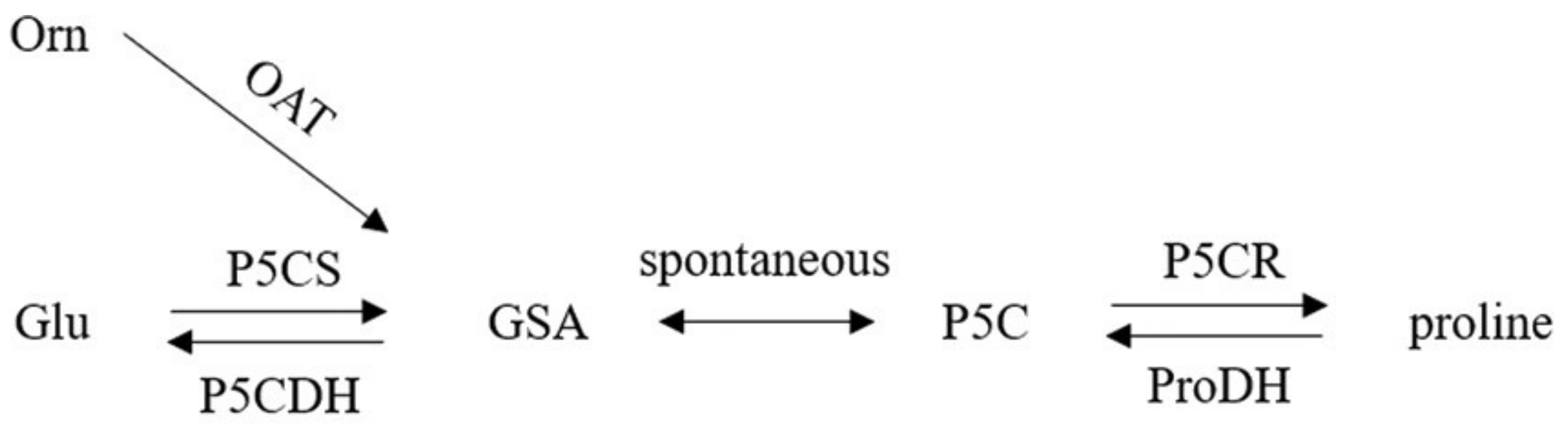
2. Proline Biosynthesis
3. Proline Sensing, Signaling and Transportation
4. The Physiological Function of Proline
5. Application of Proline through Seed Priming
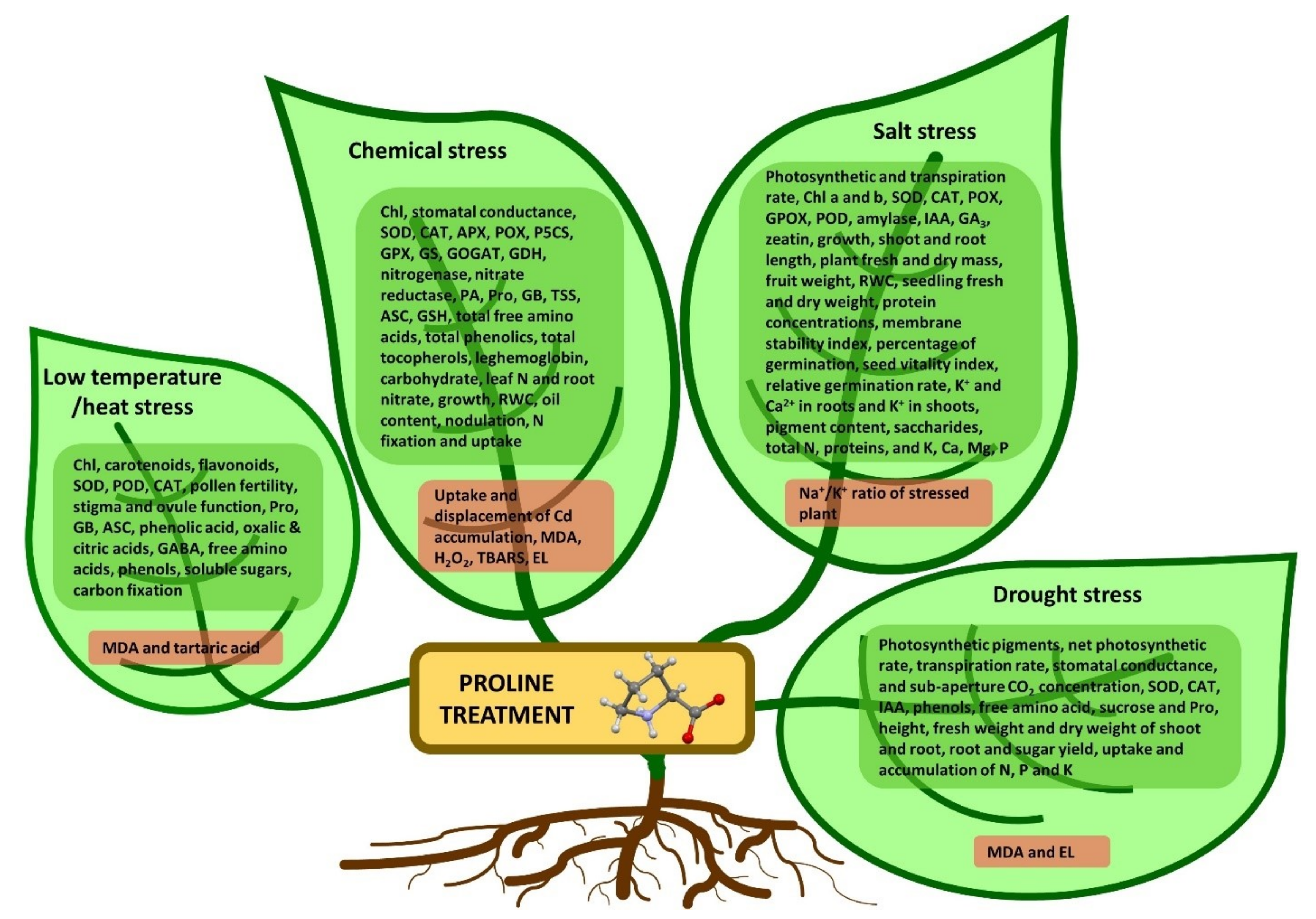
6. Foliar Application of Proline
6.1. Effect of Proline Foliar Application in Drought Stress
6.2. Effect of Proline Foliar Application in Salinity Stress
6.3. Effect of Proline Foliar Application in Temperature Stress
6.4. Effect of Proline Foliar Application in Heavy Metals Stress
7. Conclusions and Future Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stress in Plants; de Oliveira, A.B., Ed.; IntechOpen: Rijeka, Croatia, 2019; Chapter 1. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.S. Plant Productivity and Environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- FAO. The Impact of Disasters and Crises on Agriculture and Food Security: 2021; Report; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Lin, K.H.; Ho, S.L.; Chiang, C.M.; Yang, C.M. Enhancing the abiotic stress tolerance of plants: From chemical treatment to biotechnological approaches. Physiol. Plant. 2018, 164, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Penella, C.; Hernández, J.A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Navarro, J.M.; Gómez-Bellot, M.J.; Barba-Espín, G. Towards a sustainable agriculture: Strategies involving phytoprotectants against salt stress. Agronomy 2020, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Arjeh, J.; Pirzad, A.; Tajbakhsh, M.; Mohammadzadeh, S. Biochemical responses of sugar beet plant to phytoprotectants and vermicompost under moisture stress. J. Plant Physiol. Breed. 2021, 11, 17–31. [Google Scholar]
- Hossain, M.A.; Kumar, V.; Burritt, D.J.; Fujita, M.; Mäkelä, P. Osmoprotectant-mediated abiotic stress tolerance in plants. In Proline Metabolism and Its Functions in Development and Stress Tolerance; Springer Nature: Cham, Switzerland, 2019; pp. 41–72. [Google Scholar]
- Bougouffa, S.; Radovanovic, A.; Essack, M.; Bajic, V.B. DEOP: A database on osmoprotectants and associated pathways. Database 2014, 2014, bau100. [Google Scholar] [CrossRef]
- Omari Alzahrani, F. Metabolic engineering of osmoprotectants to elucidate the mechanism (s) of salt stress tolerance in crop plants. Planta 2021, 253, 1–17. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Bio Technol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Burritt, D.J. Proline and the cryopreservation of plant tissues: Functions and practical applications. Curr. Front. Cryopreserv. 2012, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Sadeghipour, O. Cadmium toxicity alleviates by seed priming with proline or glycine betaine in cowpea (Vigna unguiculata (L.) Walp.). Egypt. J. Agron. 2020, 42, 163–170. [Google Scholar] [CrossRef]
- Elewa, T.A.; Sadak, M.S.; Saad, A.M. Proline treatment improves physiological responses in quinoa plants under drought stress. Biosci. Res. 2017, 14, 21–33. [Google Scholar]
- Qirat, M.; Shahbaz, M.; Perveen, S. Beneficial role of foliar-applied proline on carrot (Daucus carota L.) under saline conditions. Pak. J. Bot. 2018, 50, 1735–1744. [Google Scholar]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline accumulation in plants: Roles in stress tolerance and plant development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: Berlin/Heidelberg, Germany, 2016; pp. 155–166. [Google Scholar]
- Adejumo, S.A.; Oniosun, B.; Akpoilih, O.A.; Adeseko, A.; Arowo, D.O. Anatomical changes, osmolytes accumulation and distribution in the native plants growing on Pb-contaminated sites. Environ. Geochem. Health 2021, 43, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Butt, M.; Ayyub, C.; Amjad, M.; Ahmad, R. Proline application enhances growth of chilli by improving physiological and biochemical attributes under salt stress. Pak. J. Agric. Sci. 2016, 53, 43–49. [Google Scholar]
- Orsini, F.; Pennisi, G.; Mancarella, S.; Al Nayef, M.; Sanoubar, R.; Nicola, S.; Gianquinto, G. Hydroponic lettuce yields are improved under salt stress by utilizing white plastic film and exogenous applications of proline. Sci. Hortic. 2018, 233, 283–293. [Google Scholar] [CrossRef]
- Priya, M.; Sharma, L.; Singh, I.; Bains, T.; Siddique, K.H.; Bindumadhava, H.; Nair, R.M.; Nayyar, H. Securing reproductive function in mungbean grown under high temperature environment with exogenous application of proline. Plant Physiol. Biochem. 2019, 140, 136–150. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Bruce, T.J.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.S. Proline metabolism and molecular cloning of AmP5CS in the mangrove Avicennia marina under heat stress. Ecotoxicology 2020, 29, 698–706. [Google Scholar] [CrossRef]
- Delaunay, A.; Verma, D. Proline accumulation and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar] [CrossRef]
- Szepesi, Á.; Szőllősi, R. Mechanism of proline biosynthesis and role of proline metabolism enzymes under environmental stress in plants. In Plant Metabolites and Regulation under Environmental Stress; Elsevier: Amsterdam, The Netherlands, 2018; pp. 337–353. [Google Scholar]
- Da Rocha, I.M.A.; Vitorello, V.A.; Silva, J.S.; Ferreira-Silva, S.L.; Viégas, R.A.; Silva, E.N.; Silveira, J.A.G. Exogenous ornithine is an effective precursor and the δ-ornithine amino transferase pathway contributes to proline accumulation under high N recycling in salt-stressed cashew leaves. J. Plant Physiol. 2012, 169, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Armengaud, P.; Thiery, L.; Buhot, N.; Grenier-de March, G.; Savouré, A. Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol. Plant. 2004, 120, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funck, D.; Baumgarten, L.; Stift, M.; Von Wirén, N.; Schönemann, L. Differential contribution of P5CS isoforms to stress tolerance in Arabidopsis. Front. Plant Sci. 2020, 11, 565134. [Google Scholar] [CrossRef] [PubMed]
- Schertl, P.; Cabassa, C.; Saadallah, K.; Bordenave, M.; Savouré, A.; Braun, H.P. Biochemical characterization of proline dehydrogenase in Arabidopsis mitochondria. FEBS J. 2014, 281, 2794–2804. [Google Scholar] [CrossRef] [Green Version]
- Ribarits, A.; Abdullaev, A.; Tashpulatov, A.; Richter, A.; Heberle-Bors, E.; Touraev, A. Two tobacco proline dehydrogenases are differentially regulated and play a role in early plant development. Planta 2007, 225, 1313–1324. [Google Scholar] [CrossRef]
- Brugière, N.; Dubois, F.; Limami, A.M.; Lelandais, M.; Roux, Y.; Sangwan, R.S.; Hirel, B. Glutamine synthetase in the phloem plays a major role in controlling proline production. Plant Cell 1999, 11, 1995–2011. [Google Scholar] [CrossRef] [Green Version]
- Mondal, R.; Kumar, A.; Chattopadhyay, S.K. Structural property, molecular regulation and functional diversity of Glutamine Synthetase in higher plants: A data-mining bioinformatics approach. Plant J. 2021, 108, 1565–1584. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Jalil, S.U.; Chopra, P.; Chhillar, H.; Ferrante, A.; Khan, N.A.; Ansari, M.I. Role of GABA in plant growth, development and senescence. Plant Gene 2021, 26, 100283. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Gagneul, D.; Aïnouche, A.; Duhazé, C.; Lugan, R.; Larher, F.R.; Bouchereau, A. A reassessment of the function of the so-called compatible solutes in the halophytic Plumbaginaceae Limonium latifolium. Plant Physiol. 2007, 144, 1598–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, D.; Grewal, S.; Kaur, J.; Singh, S. Differential proline metabolism in vegetative and reproductive tissues determine drought tolerance in chickpea. Biol. Plant. 2017, 61, 359–366. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Azeem, F.; Shahriar, S.M.; Islam, M.T. Regulation of proline transporters in salt stress response in plants. In Transporters and Plant Osmotic Stress; Elsevier: Amsterdam, The Netherlands, 2021; pp. 291–306. [Google Scholar]
- Rentsch, D.; Hirner, B.; Schmelzer, E.; Frommer, W.B. Salt stress-induced proline transporters and salt stress-repressed broad specificity amino acid permeases identified by suppression of a yeast amino acid permease-targeting mutant. Plant Cell 1996, 8, 1437–1446. [Google Scholar] [PubMed] [Green Version]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino acid transporters in plants: Identification and function. Plants 2020, 9, 972. [Google Scholar] [CrossRef] [PubMed]
- Khanna-Chopra, R.; Semwal, V.K.; Lakra, N.; Pareek, A. Proline—A key regulator conferring plant tolerance to salinity and drought. In Plant Tolerance to Environmental Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 59–80. [Google Scholar]
- Girousse, C.; Bournoville, R.; Bonnemain, J.L. Water deficit-induced changes in concentrations in proline and some other amino acids in the phloem sap of alfalfa. Plant Physiol. 1996, 111, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Verslues, P.E.; Sharp, R.E. Proline accumulation in maize (Zea mays L.) primary roots at low water potentials. II. Metabolic source of increased proline deposition in the elongation zone. Plant Physiol. 1999, 119, 1349–1360. [Google Scholar] [CrossRef] [Green Version]
- Cha-um, S.; Rai, V.; Takabe, T. Proline, glycinebetaine, and trehalose uptake and inter-organ transport in plants under stress. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2019; pp. 201–223. [Google Scholar]
- Akbudak, M.A.; Filiz, E. Genome-wide investigation of proline transporter (ProT) gene family in tomato: Bioinformatics and expression analyses in response to drought stress. Plant Physiol. Biochem. 2020, 157, 13–22. [Google Scholar] [CrossRef]
- Yamada, N.; Sakakibara, S.; Tsutsumi, K.; Waditee, R.; Tanaka, Y.; Takabe, T. Expression and substrate specificity of betaine/proline transporters suggest a novel choline transport mechanism in sugar beet. J. Plant Physiol. 2011, 168, 1609–1616. [Google Scholar] [CrossRef]
- Hare, P.; Cress, W.; Van Staden, J. Proline synthesis and degradation: A model system for elucidating stress-related signal transduction. J. Exp. Bot. 1999, 50, 413–434. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Z.; Gao, J.; Wang, P.; Hu, T.; Wang, Z.; Hou, Y.J.; Wan, Y.; Liu, W.; Xie, S.; et al. Arabidopsis duodecuple mutant of PYL ABA receptors reveals PYL repression of ABA-independent SnRK2 activity. Cell Rep. 2018, 23, 3340–3351. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Juste, J.; Alrefaei, A.F.; Rodriguez, P.L. Plant osmotic stress signaling: MAPKKKs meet SnRK2s. Trends Plant Sci. 2020, 25, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Zlobin, A.; Bloodworth, J.C.; Osipo, C. Mitogen-activated protein kinase (MAPK) signaling. In Predictive Biomarkers in Oncology; Springer: Berlin/Heidelberg, Germany, 2019; pp. 213–221. [Google Scholar]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Soma, F.; Takahashi, F.; Suzuki, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Plant Raf-like kinases regulate the mRNA population upstream of ABA-unresponsive SnRK2 kinases under drought stress. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Zhang, J.; Hsu, P.K.; Ceciliato, P.H.; Zhang, L.; Dubeaux, G.; Munemasa, S.; Ge, C.; Zhao, Y.; Hauser, F.; et al. MAP3Kinase-dependent SnRK2-kinase activation is required for abscisic acid signal transduction and rapid osmotic stress response. Nat. Commun. 2020, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Rajasheker, G.; Jawahar, G.; Jalaja, N.; Kumar, S.A.; Kumari, P.H.; Punita, D.L.; Karumanchi, A.R.; Reddy, P.S.; Rathnagiri, P.; Sreenivasulu, N.; et al. Role and regulation of osmolytes and ABA interaction in salt and drought stress tolerance. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 417–436. [Google Scholar]
- Vandana, U.K.; Singha, B.; Gulzar, A.; Mazumder, P. Chapter 18 - Molecular mechanisms in plant growth promoting bacteria (PGPR) to resist environmental stress in plants. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 221–233. [Google Scholar] [CrossRef]
- Venekamp, J.; Koot, J. The distribution of free amino acids, especially of proline, in the organs of field bean plants, Vicia faba L., during development in the field. J. Plant Physiol. 1984, 116, 343–349. [Google Scholar] [CrossRef]
- Funck, D.; Winter, G.; Baumgarten, L.; Forlani, G. Requirement of proline synthesis during Arabidopsis reproductive development. BMC Plant Biol. 2012, 12, 191. [Google Scholar] [CrossRef] [Green Version]
- Fujita, T.; Maggio, A.; Garcia-Rios, M.; Bressan, R.A.; Csonka, L.N. Comparative analysis of the regulation of expression and structures of two evolutionarily divergent genes for Delta 1-pyrroline-5-carboxylate synthetase from tomato. Plant Physiol. 1998, 118, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Momayezi, M.; Zaharah, A.; Hanafi, M.; Mohd Razi, I. Seed germination and proline accumulation in rice (Oryza sativa L.) as affected by salt concentrations. Pertanika J. Trop. Agric. Sci 2009, 32, 247–259. [Google Scholar]
- Mattioli, R.; Falasca, G.; Sabatini, S.; Altamura, M.M.; Costantino, P.; Trovato, M. The proline biosynthetic genes P5CS1 and P5CS2 play overlapping roles in Arabidopsis flower transition but not in embryo development. Physiol. Plant. 2009, 137, 72–85. [Google Scholar] [CrossRef]
- Székely, G.; Abrahám, E.; Cséplo, A.; Rigó, G.; Zsigmond, L.; Csiszár, J.; Ayaydin, F.; Strizhov, N.; Jásik, J.; Schmelzer, E.; et al. Duplicated P5CS genes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. Plant J. Cell Mol. Biol. 2008, 53, 11–28. [Google Scholar] [CrossRef] [Green Version]
- Meinke, D.; Muralla, R.; Sweeney, C.; Dickerman, A. Identifying essential genes in Arabidopsis thaliana. Trends Plant Sci. 2008, 13, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Costantino, P.; Trovato, M. Proline accumulation in plants: Not only stress. Plant Signal. Behav. 2009, 4, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- KAVI KISHOR, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Hima Kumari, P.; Sunita, M.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Kamiyama, M.; Iwata, T.; Makita, N.; Furukawa, H.; Ikeda, H. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot. 2006, 97, 731–738. [Google Scholar] [CrossRef]
- Lansac, A.; Sullivan, C.; Johnson, B. Accumulation of free proline in sorghum (Sorghum bicolor) pollen. Can. J. Bot. 1996, 74, 40–45. [Google Scholar] [CrossRef]
- Špoljarević, M.; Agić, D.; Lisjak, M.; Gumze, A.; Wilson, I.D.; Hancock, J.T.; Teklić, T. The relationship of proline content and metabolism on the productivity of maize plants. Plant Signal. Behav. 2011, 6, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Nepi, M.; von Aderkas, P.; Wagner, R.; Mugnaini, S.; Coulter, A.; Pacini, E. Nectar and pollination drops: How different are they? Ann. Bot. 2009, 104, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Schwacke, R.; Grallath, S.; Breitkreuz, K.E.; Stransky, E.; Stransky, H.; Frommer, W.B.; Rentsch, D. LeProT1, a transporter for proline, glycine betaine, and γ-amino butyric acid in tomato pollen. Plant Cell 1999, 11, 377–391. [Google Scholar]
- Majewska-Sawka, A.; Nothnagel, E.A. The multiple roles of arabinogalactan proteins in plant development. Plant Physiol. 2000, 122, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.C.; Long, F.; Maruoka, E.M.; Cooper, J.B. A new proline-rich early nodulin from Medicago truncatula is highly expressed in nodule meristematic cells. Plant Cell 1994, 6, 1265–1275. [Google Scholar]
- Aranega-Bou, P.; de la O Leyva, M.; Finiti, I.; García-Agustín, P.; González-Bosch, C. Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front. Plant Sci. 2014, 5, 488. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed priming: New comprehensive approaches for an old empirical technique. In New Challenges in Seed Biology-Basic and Translational Research Driving Seed Technology; IntechOpen: London, UK, 2016; pp. 1–46. [Google Scholar]
- Kranner, I.; Minibayeva, F.V.; Beckett, R.P.; Seal, C.E. What is stress? Concepts, definitions and applications in seed science. New Phytol. 2010, 188, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Paparella, S.; Araújo, S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Perveen, S.; Mahmood, S. Dynamic proline metabolism: Importance and regulation in water-limited environments. In Plant Metabolites and Regulation under Environmental Stress; Elsevier: Amsterdam, The Netherlands, 2018; pp. 323–336. [Google Scholar]
- Venegas-Molina, J.; Proietti, S.; Pollier, J.; Orozco-Freire, W.; Ramirez-Villacis, D.; Leon-Reyes, A. Induced tolerance to abiotic and biotic stresses of broccoli and Arabidopsis after treatment with elicitor molecules. Sci. Rep. 2020, 10, 10319. [Google Scholar] [CrossRef] [PubMed]
- Wiszniewska, A. Priming Strategies for Benefiting Plant Performance under Toxic Trace Metal Exposure. Plants 2021, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Sako, K.; Nguyen, H.M.; Seki, M. Advances in chemical priming to enhance abiotic stress tolerance in plants. Plant Cell Physiol. 2020, 61, 1995–2003. [Google Scholar] [CrossRef]
- Agami, R. Applications of ascorbic acid or proline increase resistance to salt stress in barley seedlings. Biol. Plant. 2014, 58, 341–347. [Google Scholar] [CrossRef]
- Karalija, E.; Selović, A. The effect of hydro and proline seed priming on growth, proline and sugar content, and antioxidant activity of maize under cadmium stress. Environ. Sci. Pollut. Res. 2018, 25, 33370–33380. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, A.; Nehal, N.; Sharma, N. Effect of proline on germination and seedling growth of rice (Oryza sativa L.) under salt stress. J. Pharmacogn. Phytochem. 2018, 7, 2449–2452. [Google Scholar]
- Shafiq, F.; Raza, S.; Bibi, A.; Khan, I.; Iqbal, M. Influence of proline priming on antioxidative potential and ionic distribution and its relationship with salt tolerance of wheat. Cereal Res. Commun. 2018, 46, 287–300. [Google Scholar] [CrossRef]
- Demiralay, M.; Altuntaş, C.; Sezgin, A.; Terzi, R.; Kadioğlu, A. Application of proline to root medium is more effective for amelioration of photosynthetic damages as compared to foliar spraying or seed soaking in maize seedlings under short-term drought. Turk. J. Biol. 2017, 41, 649–660. [Google Scholar] [CrossRef]
- Hua-long, L.; Han-jing, S.; Jing-guo, W.; Yang, L.; De-tang, Z.; Hong-wei, Z. Effect of seed soaking with exogenous proline on seed germination of rice under salt stress. J. Northeast Agric. Univ. 2014, 21, 1–6. [Google Scholar] [CrossRef]
- Rady, M.M.; Hemida, K.A. Sequenced application of ascorbate-proline-glutathione improves salt tolerance in maize seedlings. Ecotoxicol. Environ. Saf. 2016, 133, 252–259. [Google Scholar] [CrossRef]
- Yaqoob, H.; Akram, N.A.; Iftikhar, S.; Ashraf, M.; Khalid, N.; Sadiq, M.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Seed pretreatment and foliar application of proline regulate morphological, physio-biochemical processes and activity of antioxidant enzymes in plants of two cultivars of quinoa (Chenopodium quinoa Willd.). Plants 2019, 8, 588. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- AlKahtani, M.D.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Husnain, L.A.; AlGwaiz, H.I.; Abdelaal, K.A. Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef]
- Farooq, M.; Nawaz, A.; Chaudhry, M.; Indrasti, R.; Rehman, A. Improving resistance against terminal drought in bread wheat by exogenous application of proline and gamma-aminobutyric acid. J. Agron. Crop Sci. 2017, 203, 464–472. [Google Scholar] [CrossRef]
- Merwad, A.R.M.; Desoky, E.S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M.; Shahbaz, M.; Humera, H. Ameliorating effect of foliar applied proline on nutrient uptake in water stressed maize (Zea mays L.) plants. Pak. J. Bot 2008, 40, 211–219. [Google Scholar]
- Rozema, J.; Flowers, T. Crops for a salinized world. Science 2008, 322, 1478–1480. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Wani, A.; Ahmad, A.; Hayat, S.; Tahir, I. Is foliar spray of proline sufficient for mitigation of salt stress in Brassica juncea cultivars? Environ. Sci. Pollut. Res. 2016, 23, 13413–13423. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Kharal, M.A.; Ahmad, M.; Abbasi, G.H.; Nazli, F.; Hussain, A.; Akhtar, M.F.u.Z. Inducing salinity tolerance in red pepper (Capsicum annuum L.) through exogenous application of proline and L-tryptophan. Soil Environ. 2018, 37, 160–168. [Google Scholar]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.S.P. Impact of salt-induced toxicity on growth and yield-potential of local wheat cultivars: Oxidative stress and ion toxicity are among the major determinants of salt-tolerant capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.K.; Kumar, M.; Li, W.; Luo, Y.; Burritt, D.J.; Alkan, N.; Tran, L.S.P. Enhancing salt tolerance of plants: From metabolic reprogramming to exogenous chemical treatments and molecular approaches. Cells 2020, 9, 2492. [Google Scholar] [CrossRef]
- Abdelhamid, M.T.; Rady, M.M.; Osman, A.S.; Abdalla, M.A. Exogenous application of proline alleviates salt-induced oxidative stress in Phaseolus vulgaris L. plants. J. Hortic. Sci. Biotechnol. 2013, 88, 439–446. [Google Scholar] [CrossRef]
- Habib-ur Rehman, A.; Muhammad, A.; Abdul, W.; Amer, J. Inducing salt tolerance in canola (Brassica napus L.) by exogenous application of glycinebetaine and proline: Response at the initial growth stages. Pak. J. Bot. 2009, 41, 1311–1319. [Google Scholar]
- El-Sayed, O.M.; El-Gammal, O.; Salama, A. Effect of ascorbic acid, proline and jasmonic acid foliar spraying on fruit set and yield of Manzanillo olive trees under salt stress. Sci. Hortic. 2014, 176, 32–37. [Google Scholar] [CrossRef]
- Wu, G.; Feng, R.; Li, S.; Du, Y. Exogenous application of proline alleviates salt-induced toxicity in sainfoin seedlings. J. Anim. Plant Sci. 2017, 27, 246–251. [Google Scholar]
- Shaddad, M. The effect of proline application on the physiology ofraphanus sativus plants grown under salinity stress. Biol. Plant. 1990, 32, 104–112. [Google Scholar] [CrossRef]
- Shahbaz, M.; Mushtaq, Z.; Andaz, F.; Masood, A. Does proline application ameliorate adverse effects of salt stress on growth, ions and photosynthetic ability of eggplant (Solanum melongena L.)? Sci. Hortic. 2013, 164, 507–511. [Google Scholar] [CrossRef]
- Mahboob, W.; Khan, M.A.; Shirazi, M. Induction of salt tolerance in wheat (Triticum aestivum L.) seedlings through exogenous application of proline. Pak. J. Bot 2016, 48, 861–867. [Google Scholar]
- Dawood, M.; Taie, H.; Nassar, R.; Abdelhamid, M.; Schmidhalter, U. The changes induced in the physiological, biochemical and anatomical characteristics of Vicia faba by the exogenous application of proline under seawater stress. S. Afr. J. Bot. 2014, 93, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Imran, Q.M.; Falak, N.; Hussain, A.; Mun, B.G.; Yun, B.W. Abiotic stress in plants; stress perception to molecular response and role of biotechnological tools in stress resistance. Agronomy 2021, 11, 1579. [Google Scholar] [CrossRef]
- Argosubekti, N. A review of heat stress signaling in plants. In IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd Location: Makassar City, Indonesia, 2020; Volume 484, p. 012041. [Google Scholar]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Hussain, R.; Ayyub, C.M.; Shaheen, M.R.; Rashid, S.; Nafees, M.; Ali, S.; Butt, M.; Ali, M.; Maqsood, A.; Fiaz, S.; et al. Regulation of Osmotic Balance and Increased Antioxidant Activities under Heat Stress in Abelmoschus esculentus L. Triggered by Exogenous Proline Applications. Agronomy 2021, 11, 685. [Google Scholar] [CrossRef]
- Posmyk, M.; Janas, K. Effects of seed hydropriming in presence of exogenous proline on chilling injury limitation in Vigna radiata L. seedlings. Acta Physiol. Plant. 2007, 29, 509–517. [Google Scholar] [CrossRef]
- Nawaz, G.; Han, Y.; Usman, B.; Liu, F.; Qin, B.; Li, R. Knockout of OsPRP1, a gene encoding proline-rich protein, confers enhanced cold sensitivity in rice (Oryza sativa L.) at the seedling stage. 3 Biotech 2019, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mohammadrezakhani, S.; Hajilou, J.; Rezanejad, F.; Zaare-Nahandi, F. Assessment of exogenous application of proline on antioxidant compounds in three Citrus species under low temperature stress. J. Plant Interact. 2019, 14, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Koç, E. The effect of exogenous proline and salicylic acid application on proline and apoplastic protein in cold tolerance of pepper callus cultures. Int. J. Agric. Biol. 2013, 15, 382–385. [Google Scholar]
- Tangahu, B.V.; Sheikh Abdullah, S.R.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M. A review on heavy metals (As, Pb, and Hg) uptake by plants through phytoremediation. Int. J. Chem. Eng. 2011, 2011, 939161. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Dubey, G.; Prasad, S.M. Exogenous proline application ameliorates toxic effects of arsenate in Solanum melongena L. seedlings. Ecotoxicol. Environ. Saf. 2015, 117, 164–173. [Google Scholar] [CrossRef]
- Shahid, M.A.; Balal, R.M.; Pervez, M.A.; Abbas, T.; Aqeel, M.A.; Javaid, M.M.; Garcia-Sanchez, F. Exogenous proline and proline-enriched Lolium perenne leaf extract protects against phytotoxic effects of nickel and salinity in Pisum sativum by altering polyamine metabolism in leaves. Turk. J. Bot. 2014, 38, 914–926. [Google Scholar] [CrossRef] [Green Version]
- Alyemeni, M.N.; Hayat, Q.; Hayat, S.; Faizan, M.; Faraz, A. Exogenous proline application enhances the efficiency of nitrogen fixation and assimilation in chickpea plants exposed to cadmium. Legume Res. Int. J. 2016, 39, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Zouari, M.; Ahmed, C.B.; Elloumi, N.; Bellassoued, K.; Delmail, D.; Labrousse, P.; Abdallah, F.B.; Rouina, B.B. Impact of proline application on cadmium accumulation, mineral nutrition and enzymatic antioxidant defense system of Olea europaea L. cv Chemlali exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2016, 128, 195–205. [Google Scholar] [CrossRef]
- Yan, L.; Riaz, M.; Jiang, C. Exogenous application of proline alleviates B-deficiency-induced injury while aggravates aluminum toxicity in trifoliate orange seedlings. Sci. Hortic. 2020, 268, 109372. [Google Scholar] [CrossRef]
- Aggarwal, M.; Sharma, S.; Kaur, N.; Pathania, D.; Bhandhari, K.; Kaushal, N.; Kaur, R.; Singh, K.; Srivastava, A.; Nayyar, H. Exogenous proline application reduces phytotoxic effects of selenium by minimising oxidative stress and improves growth in bean (Phaseolus vulgaris L.) seedlings. Biol. Trace Elem. Res. 2011, 140, 354–367. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Plant | Stress | Observed Effects | Reference |
|---|---|---|---|
| Chenopodium quinoa Willd. | Low-temperature (Seeds kept at 4 °C for 20 h before sowing) | Plant growth improved by increasing the content of chlorophyll, carotenoids, proline, ascorbic acid, total free amino acids, phenols, total soluble sugars, the activity of SOD, POD and CAT and reducing MDA | Yaqoob et al., 2019 [90] |
| Hordeum vulgare | Salinity (0, 100 and 200 mM NaCl) | Proline treatment decreased the salinity impacts by increasing the activity of antioxidant enzymes (SOD, CAT and POX) and affecting leaf anatomy | Agami, 2014 [83] |
| Oryza sativa L. | Salinity (0 and 100 mM NaCl) | Rice germination, relative germination rate and, the amylase activities under stress heve been increased | Hua-long et al., 2014 [88] |
| Oryza sativa L. | Salinity (0, 50, 100, 150 and 200 mM NaCl) | The percentage of germination, seed vitality index, and alpha-amylase activity significantly increased | Singh et al., 2018 [85] |
| Triticum aestivum L. | Salinity (0 and 150 mM NaCl) | The growth and activity of the antioxidant enzyme improved, which depends in part on the plant’s ability to absorb sodium and distribute it to the roots | Shafiq et al., 2018 [86] |
| Vigna unguiculata L. | Cadmium (0 and 100 M cadmium chloride) | The uptake and displacement of cadmium and MDA reduced, the amount of chlorophyll, stomatal conductance, and RWC, the activity of SOD, CAT and APX increased | Sadeghipour, 2020 [12] |
| Zea mays L. | Drought (0 and −0.5 MPa by polyethylene glycol (PEG 6000)) | Root treatment by 1 mM PRO were more effective than foliar spraying on water potential, chlorophyll content, proline level, net photosynthetic rate, transpiration rate, stomatal conductance, and sub-aperture concentration | Demiralay et al., 2017 [87] |
| Zea mays L. | Salinity (0 and 100 mM NaCl) | IAA, GA3, zeatin, SOD, GPOX Activities, protein concentrations, RWC, membrane stability index increased and ABA and CAT decreased under salt stress | Rady and Hemida, 2016 [89] |
| Zea mays L. | Cadmium (0, 5 and 50 M Cd ions obtained from CdCl2.5HO) | Improving defensive mechanisms, like proline and sugar synthesis and increasing Cd accumulation induced by proline priming | Karalija and Selović, 2018 [84] |
| Plant | Stress | Observed Effects | Reference |
|---|---|---|---|
| Beta vulgaris | Normal irrigation and 50% field capacity | The application of silicon and proline increased CAT and SOD activity, root and sugar yield, sucrose%, Chl content, phenolic compounds and reduced RWC, MDA and EL | Alkahtani et al., 2021 [92] |
| Chenopodium quinoa | Normal irrigation and skipping two irrigation times at 45 and 52 days after sowing | Exogenous proline has the greatest effect on photosynthetic pigments, IAA, phenols, free amino acid and proline content, plant height, fresh and dry weight of shoot and root | Elewa et al., 2017 [13] |
| Triticum aestivum L. | Normal irrigation and drought at 35% water holding capacity | Application of proline at 150 ppm and GABA at 100 ppm were recommended as the most effective concentrations on chl, proline, glycine betaine and total soluble phenolics contents and reduced MDA | Farooq et al., 2017 [93] |
| Zea mays L. | Full field and 60% field capacity | Application of proline in modulating drought stress by promoting the uptake and accumulation of N, P and K | Ali et al., 2008 [95] |
| Plant | Stress | Observed Effects | Reference |
|---|---|---|---|
| Brassica juncea | 2.8, 4.2 and 5.6 ds·m NaCl | Although proline (20 mM) improved yield in plants without stress and under mild stress, it was not effective at high salinity stress levels | Wani et al., 2016 [99] |
| Capsicum annuum L. | 0 and 50 mM NaCl | The greatest improvement in SOD and CAT activities, photosynthetic and transpiration rate, shoot and root length, plant fresh and dry mass was obtained at a concentration of 0.8 mM proline | Butt et al., 2016 [18] |
| Capsicum annuum L. | 0.6, 4.04 and 6.11 ds·m | Proline and L-tryptophan treatment improved shoot dry weight, fruit weight, RWC, Photosynthetic pigments and decreased the Na/K ratio | Jamil et al., 2018 [100] |
| Daucus carota L. | 0 and 150 NaCl | Proline application increased the activity of SOD, POD, free proline content, and K and Ca in roots and K in shoots | Qirat et al., 2018 [14] |
| Phaseolus vulgaris L. | 1.84, 6.03 and 8.97 ds·m | SOD, CAT and POD, carotenoids, ascorbic acid, endogenous proline, the concentrations of P and the K:Na ratio under salt stress by exogenous proline. | Abdelhamid et al., 2013 [104] |
| Raphanus sativus | 0, 40, 80, 120 and 160 mM NaC1 | Transpiration rate, stomatal frequency, pigment content, saccharides, total nitrogen, proteins, and some nutrients (K, Ca, Mg, P) improved at low and moderate salt stress | Shaddad, 1990 [108] |
| Solanum melongena L. | 0 and 150 mM NaC1 | Proline spraying did not affect the modulation of salinity stress impacts | Shahbaz et al., 2013 [109] |
| Triticum aestivum L. | 0, 60 and 120 Mm NaCl | Root and shoot length, seedling fresh and dry weight, Chl a and b content, TSS, Pro, GB, TPC, and K content, and K:Na ratio improved in stressed plants | Mahboob et al., 2016 [109] |
| Vicia faba | 0.23 ds·m tap water, 3.13 and 6.25 ds·m diluted seawater | Photosynthetic pigments, N, P, K, and Ca content, K:Na ratio, and total soluble carbohydrates increased, while Na, Cl, phenolic content, free amino acids, and proline reduced | Dawood et al., 2014 [111] |
| Plant | Stress | Observed Effects | Reference |
|---|---|---|---|
| Abelmoschus esculentus L. | Heat (Gradually increasing the temperature of the growth chamber from 28/22 °C to 45/35 °C (day/night)) | Shoot length, leaves per plant, SOD, POD and CAT activity, leaf Pro, GB, total free amino acids, and Chl content increased | Hussain et al., 2021 [115] |
| Citrus reticulata/ C. sinensis/ C. paradisi | low-temperature (shoots exposed to temperatures 1, −1 and −3 °C for three hours) | The amount of phenolic acid, flavonoids, oxalic, citric, and ascorbic acids, gamma-aminobutyric acid, endogenous proline, APX, and CAT increased and tartaric acid reduced | Mohammadrezakhani et al., 2019 [118] |
| Lactuca sativa | Salinity/Light/Heat (0 and 15 mM NaCl. Transparent or white film Late spring, summer and fall) | The combine both white covering film and 5 M proline application under the salinity of 15 mM, improved efficient preservation of plant growth, photosynthetic rate, Chl content and yield | Orsini et al., 2018 [19] |
| Capsicum annuum L. | Cold (Callus tissues were developed at 4, 8, 16 and 24 °C) | The maximum amount of endogenous proline and apoplastic protein at the lowest temperature (4 °C) was observed by the simultaneous application of proline (24 mM) and SA (0.25 mM) | Koç, 2013 [119] |
| Vigna radiata L. | high-temperature (30-day old plants were exposed to 35/23 °C, 40/28 °C and 45/33 °C (day/night)) | Pollen fertility, stigma and ovule function, carbon fixation, and assimilative capacity of heat-stressed mung bean plants were improved | Priya et al., 2019 [20] |
| Plant | Stress | Application | Observed Effects | Reference |
|---|---|---|---|---|
| Cicer arietinum L. | Cadmium (0, 25, 50 or 100 mg of cadmium per kg of soil) | Foliar | Nodulation, nitrogen fixation and uptake, leghemoglobin, carbohydrate, content of nitrogen in leaf and nitrate in root, the activity of nitrogenase, nitrate reductase, GS, GOGAT and GDH increased | Alyemeni et al., 2016 [123] |
| Olea europaea L. cv Chemlali | Cadmium (0, 10 and 30 mg CdCl per kg of soil) | Irrigation | The concentration of proline 20 mM enhancing oil and proline content, SOD, CAT, APX and GPX and reducing HO, TBARS and EL | Zouari et al., 2016 [124] |
| Phaseolus vulgaris L. | Selenium (0, 1, 2, 4, and 6 ppm) | Culture medium | Chl and endogenous proline content, RWC, SOD, CAT, APX activity, ASC and GSH content increased and HO reduced | Aggarwal et al., 2011 [126] |
| Pisum sativum L. | Salinity/Nickel (0 and 100 mM NaCl 0 and 100 M Nickel) | Foliar | Pure proline and natural proline (Lolium perenne L. leaf extract) detoxified the stress caused by NiCl and/or NaCl. LP extract can be used as a cheap source of proline to enhance growth, polyamine metabolism, photosynthetic activity, RWC, organic osmolytes. | Shahid et al., 2014 [122] |
| Poncirus trifoliata | Boron deficiency/ aluminum toxicity (0.1 M B, 300 M Al and 0.1 M B + 300 M Al) | Nutrient solution | Exogenous proline enhanced cellulose and protein content. In contrast, higher MDA and HO contents inhibited the plant growth | Yan et al., 2020 [125] |
| Solanum melongena L. | Arsenate (0, 5 and 25 M) | Seedling treatment | Exogenous proline decreased the accumulation of As and attenuated its toxicity by increasing the activity of SOD, CAT, POX, and P5CS as well as endogenous Proine | Singh et al., 2015 [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. https://doi.org/10.3390/ijms23095186
Hosseinifard M, Stefaniak S, Ghorbani Javid M, Soltani E, Wojtyla Ł, Garnczarska M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. International Journal of Molecular Sciences. 2022; 23(9):5186. https://doi.org/10.3390/ijms23095186
Chicago/Turabian StyleHosseinifard, Marjanossadat, Szymon Stefaniak, Majid Ghorbani Javid, Elias Soltani, Łukasz Wojtyla, and Małgorzata Garnczarska. 2022. "Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review" International Journal of Molecular Sciences 23, no. 9: 5186. https://doi.org/10.3390/ijms23095186