1. Introduction
The Gram-positive bacterium
Cutibacterium acnes (
C. acnes), previously known as
Propionibacterium acnes, is a commensal, anaerobic, aerotolerant, lipophilic bacterium found mostly in sebum-rich areas within the pilosebaceous unit (PSU) in normal human skin [
1,
2].
C. acnes strains are currently classified into eight phylotypes (IA-1, IA-2, IB-1, IB-2, IB-3, IC, II, III), with the IA-1 and IA-2 clades preferentially found in acne lesions, whereas phylotypes II and III are found in deeper infections [
3,
4,
5,
6,
7].
C. acnes has been implicated in inflammatory acne, but its pathophysiological role remains a matter of debate. Until recently, it was thought that the obstruction of the pilosebaceous unit was due to an increase in sebum production associated with hyperkeratinization, favoring hypoxic conditions and resulting in the proliferation of
C. acnes strains; however, the
C. acnes load is similar in PSUs from healthy and acne-affected skin [
8]. Moreover,
C. acnes appears to be the major bacterial genus present on skin. It plays a crucial role in the maintenance of skin health, whereas dysbiosis in the cutaneous microbiota can lead to the selection of specific
C. acnes lineages, such as the IA1/IA2 phylotypes found predominantly in acne lesions. This phenomenon could be amplified by hyperseborrhea and the change in sebum composition observed in acne, with an increase in proinflammatory lipid content [
9,
10,
11].
However, comparisons of the genomes of
C. acnes phylotype IA1 isolates from healthy individuals and acne patients revealed no differences [
12]. We can, therefore, hypothesize that the expression of specific components by
C. acnes strains may influence their capacity to colonize the host and stimulate the host’s innate immune system [
13].
C. acnes is currently considered to be an opportunistic pathogen of low pathogenicity, as it is also isolated in deeper infections, such as endocarditis and postsurgical infections.
C. acnes strains can induce inflammatory responses via the toll like receptor (TLR)-2 and TLR-4 pathways, by activating the nuclear factor kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) cascades, thereby inducing the production of proinflammatory molecules, such as interleukin (IL)-1α/β, IL-6, C-X-C motif chemokine ligand interleukin (CXCL)8/(IL)-8, IL-12, interferon (IFN), granulocyte macrophage-colony stimulating factor (GM-CSF), tumor necrosis factor (TNF)-α and human beat defensin (hBD)-2 in keratinocytes, sebocytes and monocytes in vitro, but also in acne lesions ex vivo [
14,
15,
16,
17,
18,
19,
20].
C. acnes strains can produce virulence factors (biofilm, surface proteins), triggering the immune response in the host or enhancing the ability of
C. acnes to adapt to its environment [
8,
21]. Several genes encoding putative virulence factors have been identified in the
C. acnes genome [
22], and transcriptomic analysis revealed strong expression of potential virulence factors, such as the dermatan-sulfate adhesins (DsA1 and DsA2), the Christie–Atkins–Munch–Petersen (CAMP) hemolytic factors, the polyunsaturated fatty acid isomerase, the HtaA iron acquisition protein and GehA lipase and heat-shock proteins, such as HSP20, DnaK, DnaJ, GrpE and GroEL [
23]. We previously characterized the
C. acnes hemolytic factor CAMP1, and showed that it was specifically recognized by TLR-2, with a high degree of polymorphism of the CAMP1 protein sequence associated with strong CAMP1-TLR-2 binding in
C. acnes strains producing large amounts of CXCL8/IL-8 in vitro [
24].
In this study, we produced polyclonal antibodies directed against the CAMP1 protein, which we used to analyze the expression levels of this protein in C. acnes surface protein extracts. We also cloned and produced non-mutated and mutated CAMP1 proteins and assessed their ability to induce the production of proinflammatory molecules in vitro in keratinocytes and ex vivo in human skin explants. Finally, we identified the CAMP1 peptide sequences involved in TLR-2 binding and CAMP1-related peptides capable of decreasing TLR-2 binding and the production of proinflammatory molecules in vitro and ex vivo.
3. Discussion
Christie–Atkins–Munch–Petersen (CAMP) factors are proteins that can form pores in host membranes, contributing to tissue damage. They were discovered through a synergic interaction between a CAMP factor from
Streptococcus agalactiae and the β-toxin (sphingomyelinase or SMase) of
Staphylococcus aureus, inducing the formation of pores in sheep erythrocyte membranes, leading to the lysis of these cells [
27]. SMases can hydrolyze sphingomyelin, a component of the erythrocyte membrane. The membrane is vulnerable if it has a sphingomyelin content of at least 45%; rabbit, mouse and human erythrocytes are therefore not susceptible to CAMP factors, whereas goat, sheep and cow erythrocytes are [
28].
An analysis of the
C. acnes genome has revealed the presence of five CAMP factor genes (
camp1,
camp2,
camp3,
camp4 and
camp5), all considered to be putative virulence factors [
29]. The hemolytic properties of
C. acnes in situ in cases of acne remain unclear, but it has been shown that
C. acnes can use the host SMase to enhance its virulence [
30], and that
C. acnes strains displaying hemolytic activity have been isolated from the intervertebral discs of patients with chronic lower back pain [
31].
C. acnes CAMP factor 2 is considered to be a virulence factor because it can induce inflammation in vivo and has been used as an antigen to elicit monoclonal antibodies, decreasing the production of CXCL8/IL-8 and IL-1β in ex vivo skin explants from patients with acne [
32,
33]. By contrast, the role of CAMP1 remains unknown. However, CAMP1 is mostly expressed by
C. acnes in the PSU [
34], suggesting a role in triggering inflammation.
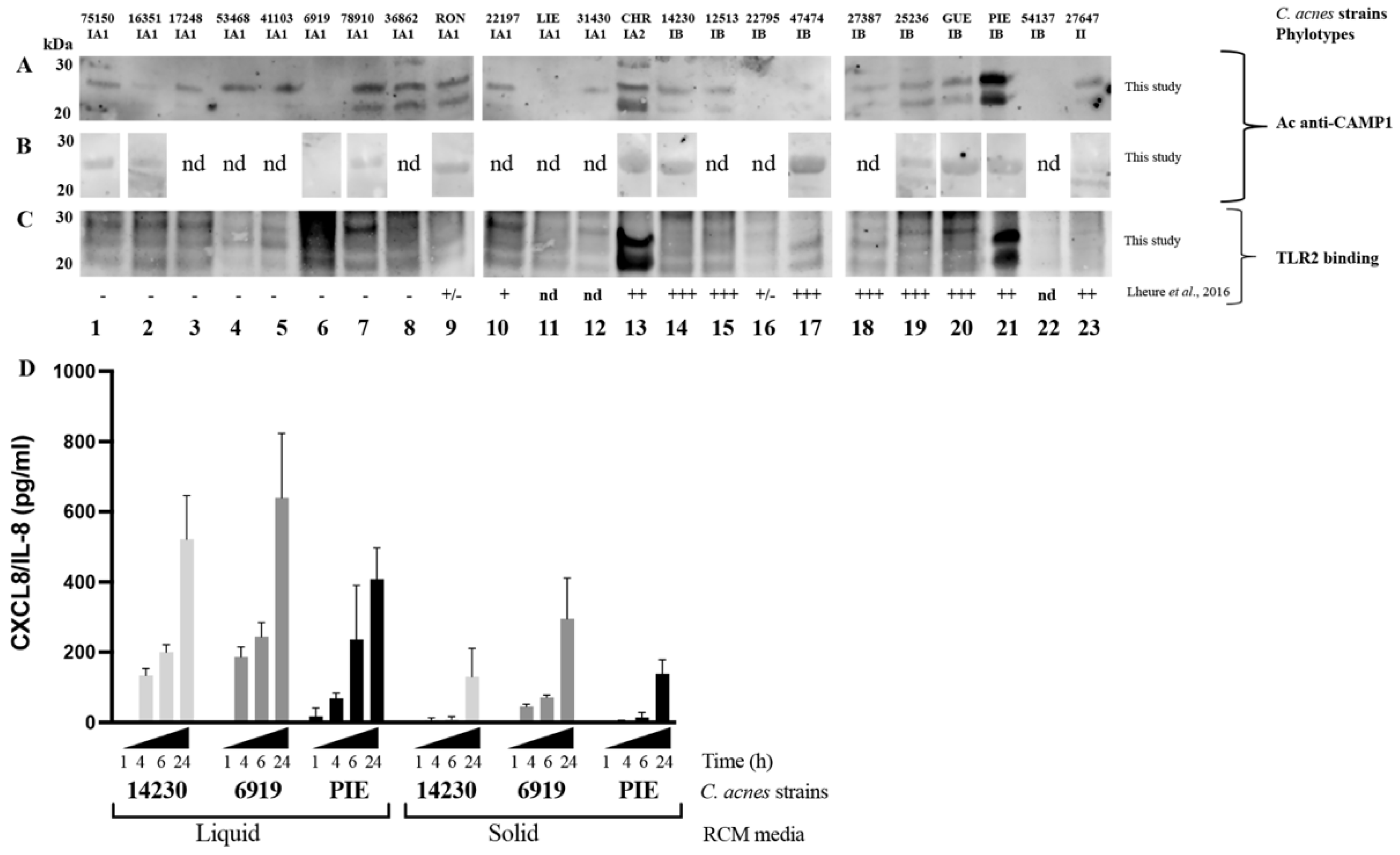
We previously showed that
C. acnes phylotype IA1 strains contain a CAMP1 protein that is only poorly recognized by TLR2, if at all [
24]. In this study, using polyclonal antibodies against CAMP1, we showed that CAMP1 was present in
C. acnes strains of phylotype IA1. We also detected CAMP1 in
C. acnes phylotypes IA2 and IB, consistent with the findings of a previous study [
35]. Our results conflict with those of Lheure et al. [
24] for phylotype II, in which we were unable to detect CAMP1 binding to TLR2, despite performing several experiments; however, a larger number of
C. acnes strains of phylotypes IA2, II and III will need to be analyzed, to evaluate the expression of CAMP1 in this species as a whole. CAMP1-TLR-2 binding activity assays detected CAMP1 in phylotypes IA2 and IB, as in our previous study [
24]. Anti-CAMP1 antibodies detected the presence of a single protein band at about 27 kDa, or two protein bands at about 24 and 27 kDa, as previously described in a study using TLR-2 as the probe [
24]. We investigated whether growth conditions affected CAMP1 production, by growing
C. acnes strains on a solid medium. Surprisingly, CAMP1 was detected as a single 27 kDa band in these conditions, regardless of phylotype. Moreover,
C. acnes seemed to induce higher levels of CXCL8/IL-8 production when grown in a liquid medium, suggesting that growth conditions might influence CAMP1 expression and the ability of strains to induce chemokine production. This finding is consistent with a previous study reporting the differential expression of surface proteins according to the culture medium [
36]. With the anti-CAMP1 antibody, we showed that keratinocytes stimulated with
C. acnes phylotype IB, which expresses a CAMP1 recognized by TLR-2, produced less CXCL8/IL-8 production than those stimulated with other phylotypes. We found that higher levels of CAMP1 in a
C. acnes strain were associated with a stronger effect of the anti-CAMP1 antibody. Conversely, stimulation with
C. acnes phylotype IA1, which produces a CAMP1 that is poorly recognized by TLR-2, if at all, resulted in no decrease in CXCL8/IL-8 production. Control experiments with antibodies against TLR-2 showed that CXCL8/IL-8 production decreased with all phylotypes tested, as in previous studies [
14,
24]. Overall, these results indicate that CAMP1 is expressed in phylotypes IA1, IA2 and IB, and that the CAMP1 protein recognized by TLR-2 is involved in CXCL8/IL-8 expression in keratinocytes; however, more investigations and the generation of Δ
camp1 C. acnes strains will be required to determine the impact of CAMP1 on chemokine production.
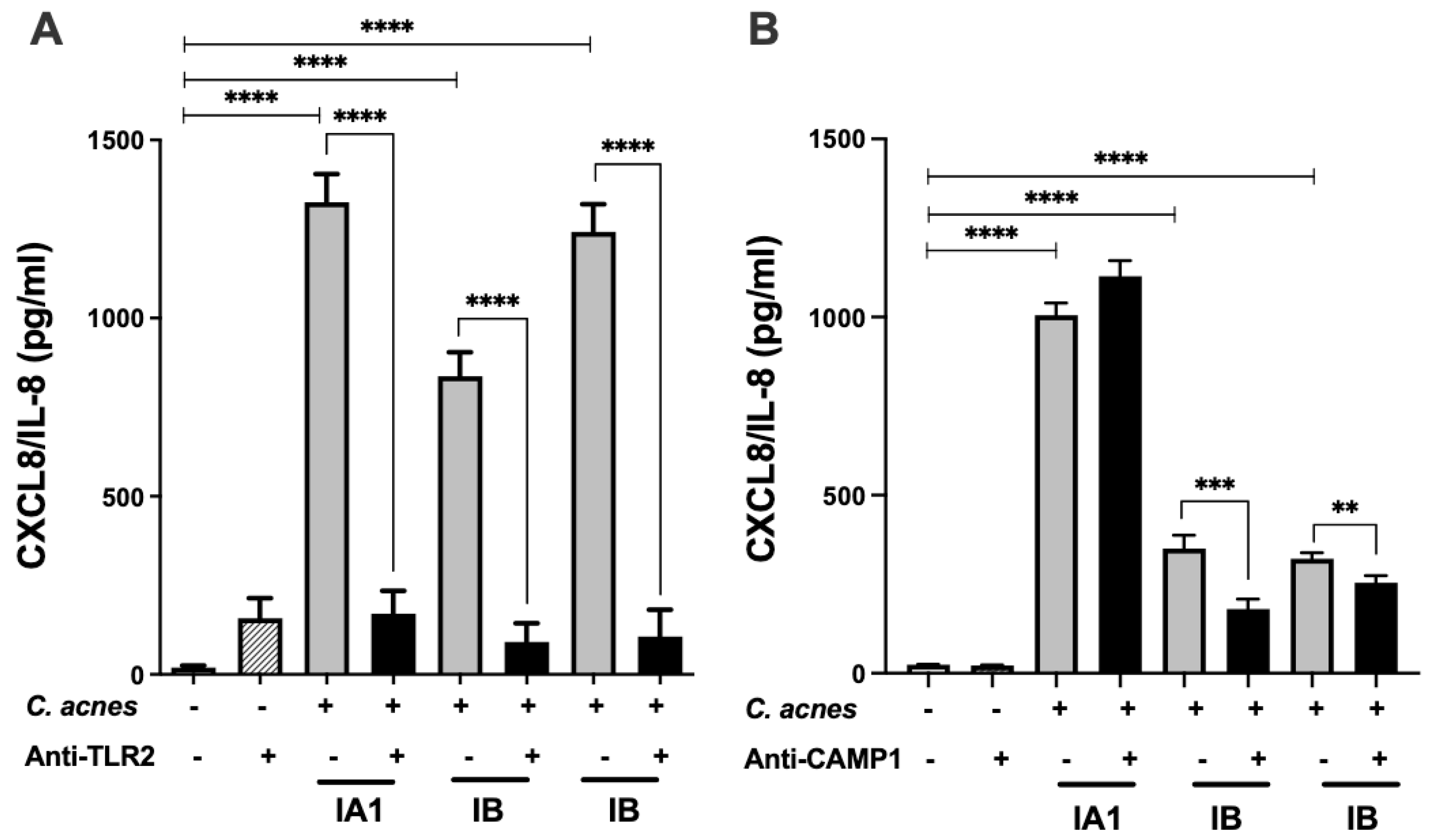
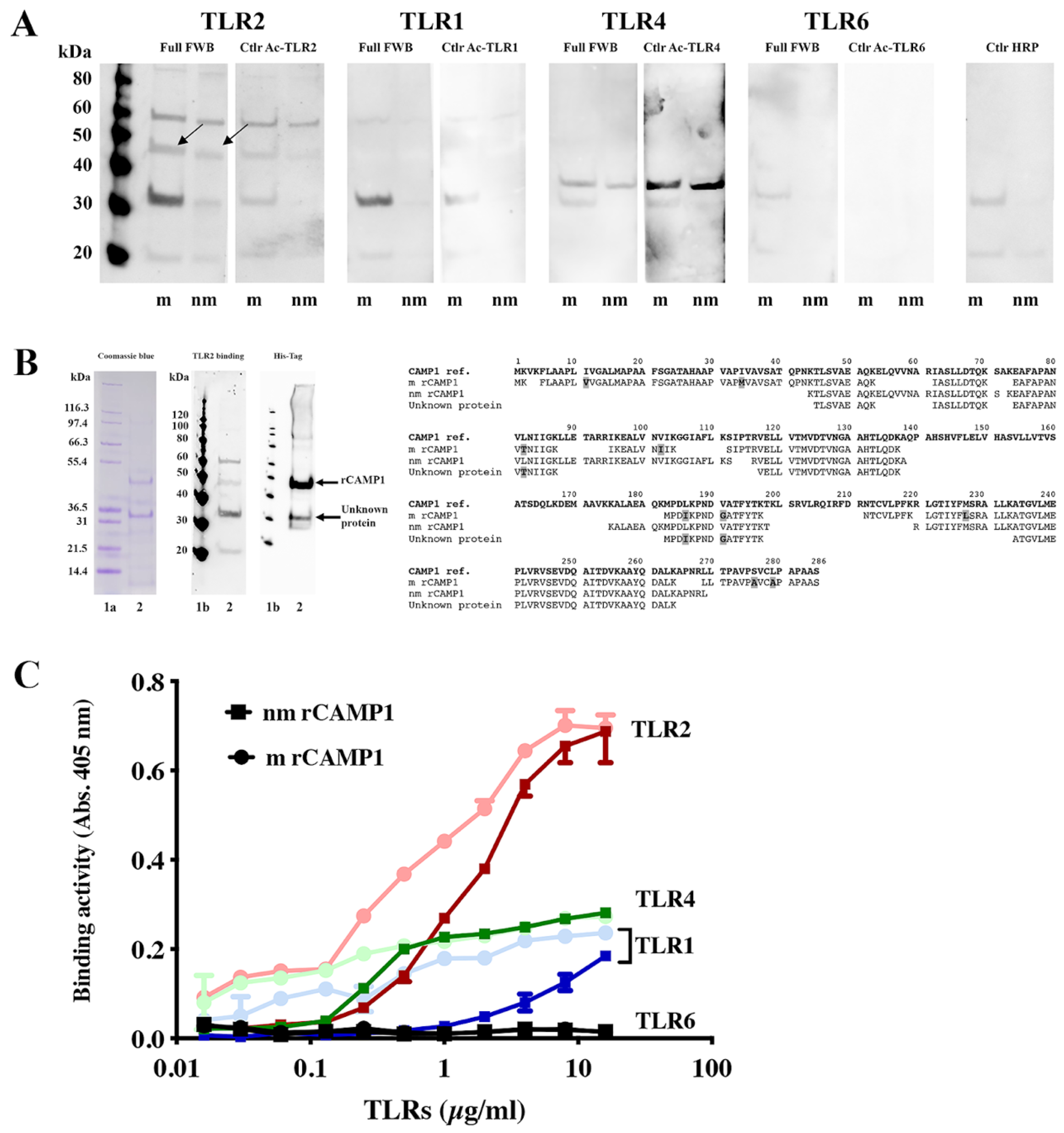
We evaluated the binding of CAMP1 to TLR-2 by producing recombinant proteins from
C. acnes strains expressing non-mutated and mutated CAMP1. Far-Western blotting experiments showed that rCAMP1 was recognized by TLR-2, but not by TLR-1, TLR-4 and TLR-6. This result is consistent with a previous study in which a
C. acnes surface protein was identified as CAMP1 [
24]. The quantitative difference in TLR-2 binding to nmrCAMP1 and mrCAMP1 was not significant, but the intensity of the signal was lower for nmrCAMP1. This result is consistent with a previous study in which
C. acnes strains expressing an unmutated CAMP1 protein displayed weaker TLR-2 binding [
24]. We detected a band at about 60 kDa that was recognized by TLR-2. This protein was also recognized by the anti-TLR2 secondary antibody, so the reaction was considered non-specific. We also detected a protein at about 30 kDa that was recognized strongly by TLR-2, more weakly by TLR-1 and not all by TLR-4 and TLR-6. This 30 kDa protein was recognized by the His-Tag antibody and proteomic analysis showed it to be a component of the CAMP1 protein. This result suggests possible differential splicing of rCAMP1 during its synthesis in
E. coli. The quantification of rCAMP1-TLR binding activities showed that recognition by TLR-2 was dose-dependent and saturable, consistent with a specific binding process. TLR-1 and TLR-4 displayed very moderate binding activities, whereas no binding was detected with TLR-6. This result is consistent with the far-Western blotting experiment and indicates that CAMP1-TLR-2 recognition is specific.
We investigated the CAMP1-TLR-2 interaction further by using a peptide spot array library of the CAMP1 protein sequence to identify the parts of the molecule involved in binding to TLR-2 binding. This method made it possible to screen a large number of peptides, as previously described [
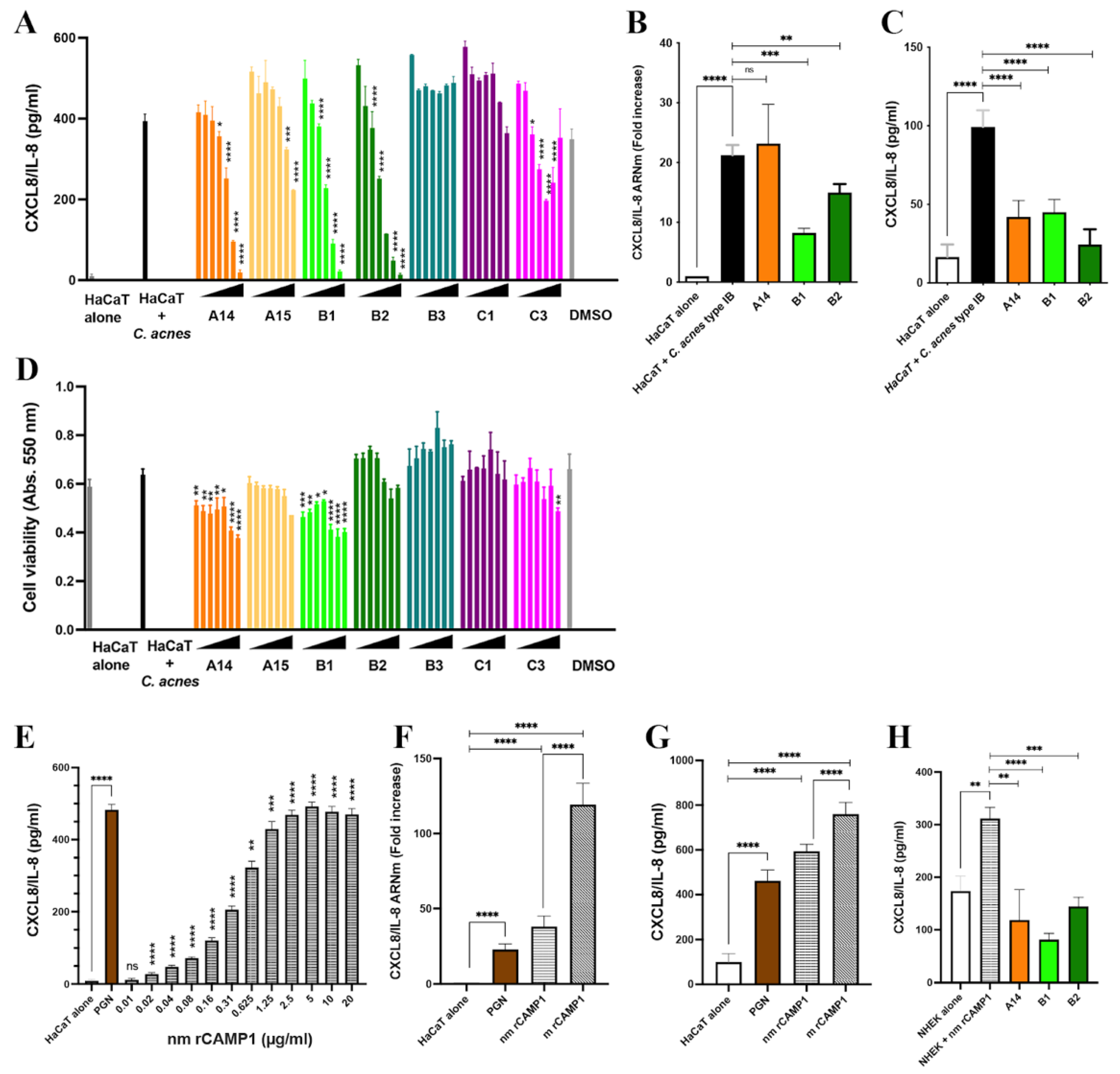
37]. Seven of the 15-mer peptides were specifically recognized by TLR-2. We eliminated the C17 peptide, because it also gave a high level of binding with the control, indicating that the interaction was non-specific. Moreover, the C17 peptide did not belong to the same group as the peptides surrounding its amino acid sequence. Indeed, the selected peptides belonged to three groups (A14-A15, B1-B2-B3, C1-C3) corresponding to overlapping sequences, consistent with a role for a consensus amino acid sequence in binding; however, a competitive binding inhibition experiment showed that A14 and B2 partially inhibited CAMP1-TLR-2 binding in a dose-dependent manner, whereas the other peptides either failed to inhibit this binding or did so only at the highest concentration. The discrepancy between the peptide array and plate inhibition assay results may reflect differences in peptide spatial presentation, as the peptide is immobilized on cellulose in the array and in solution during the inhibition assay on plates. These results suggest that A14 and B2 have the best binding inhibition capacities of the CAMP1 peptides recognized by TLR-2.
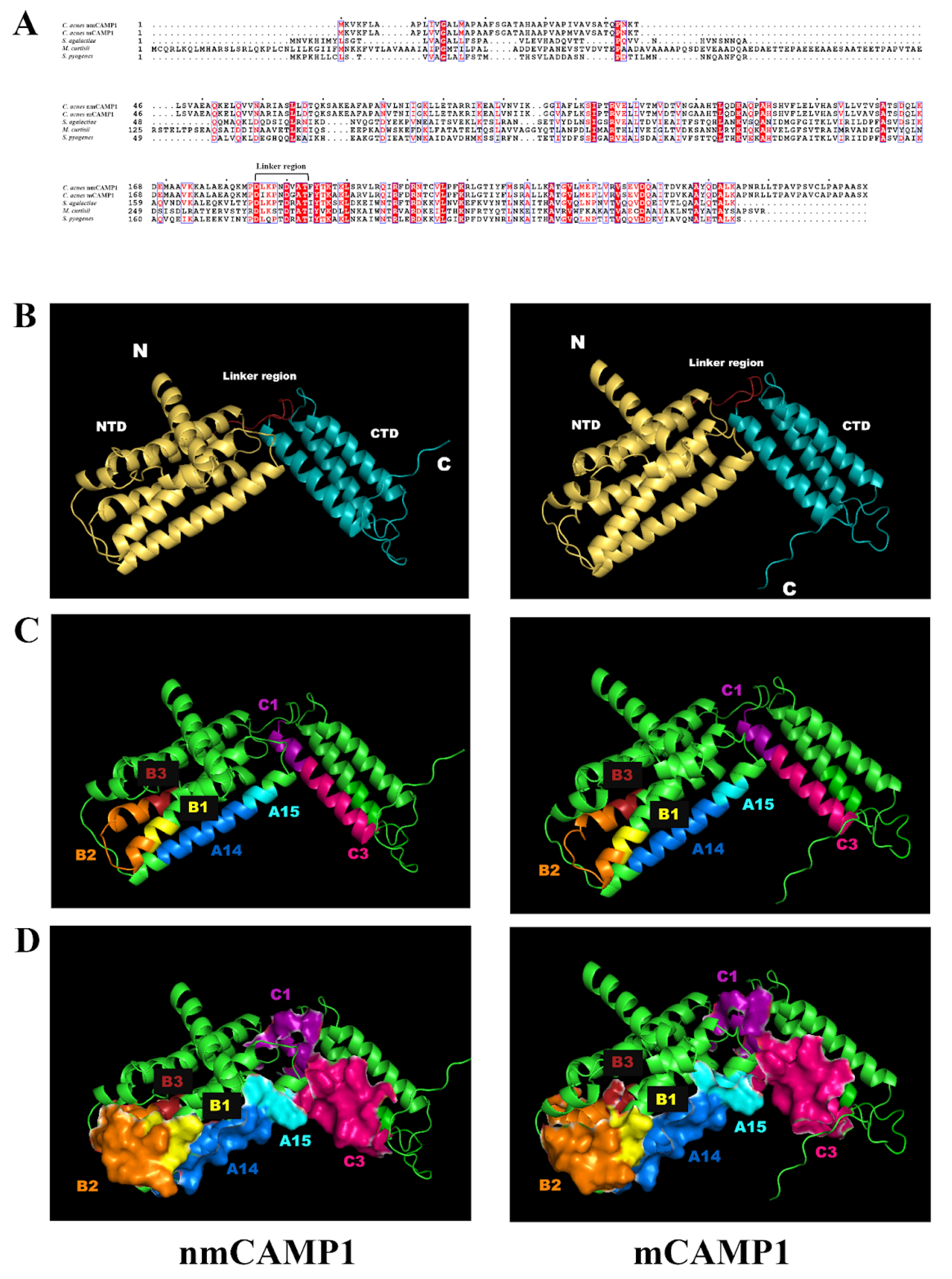
We were unable to crystallize rCAMP1 in this study because its purity was not sufficiently high; however, 3D-crystal structure analyses have already been performed on CAMP factors from
Mobiluncus curtisii and
Streptococcus agalactiae [
38,
39], and a comparison between the sequences of the
S. agalactiae CAMP protein and the
C. acnes CAMP1 protein revealed a sufficient level of sequence identity for in silico 3D structure analysis [
25]. The 3D structure of
C. acnes CAMP1 appears to be similar to that of the
S. agalactiae CAMP protein, with N-terminal and C-terminal domains separated by a linker region. The peptides recognizing TLR-2 were found to be located on the same side of the structure, with the A14 and B2 close to each other spatially. TLR-2 belongs to the Toll-IL-1 receptor superfamily. The receptors of this family have an intracellular Toll interleukin-1 receptor domain (TIR) responsible for signaling pathway activation and an extracellular leucine-rich repeat domain (LRR) responsible for the interaction with lipopeptides. TLR-2 forms heterodimers with TLR-1 and TLR-6, which in turn, recognize di- and tri-acylated lipoproteins via the hydrophobic pocket [
40]; however, several studies have shown that proteins produced by
Neisseria meningitidis or
Vibrio cholerae can also activate TLR-2 to induce proinflammatory cytokine responses in a TLR2-dependent manner [
41,
42]. Polysaccharides also seem to be able to interact with TLR-2 [
43]. Moreover, a small molecule, diprovocim, with a structure very different from the canonical lipostructures recognized by TLR-2, was able to bind to the ligand-binding pocket of the heterodimerized LRR of TLR-2 [
44]. No crystal structure showing the mechanism of TLR-2 binding to proteinaceous ligands has yet been obtained. Further studies will be required to crystallize the
C. acnes CAMP1 for the determination of its 3D structure and investigation of the CAMP1-TLR-2 interaction.
C. acnes can induce high levels of chemokine and cytokine production by the various target cells it encounters in the course of inflammatory acne. We showed that both the nmrCAMP1 and mrCAMP1 proteins induce a strong proinflammatory molecule response in HaCaT and primary keratinocytes. This result confirms previous findings that CAMP1 proteins eluted from electrophoretic gels can induce CXCL8/IL-8 production by activating the TLR-2 signaling pathway [
24]. The A14, B1 and B2 peptides yielded a dose-dependent decrease in CXCL8/IL-8 production, whereas the other peptides recognized by TLR-2 (A15, B3, C1, C3) did not, suggesting that a specific amino acid sequence and/or conformation is involved in this inhibition. No major impact on cell viability was observed, indicating that the decrease in CXCL8/IL-8 production did not result from cell death. Subsequent investigations focused on peptides A14, B1 and B2, which were the only ones with a significant effect on CXCL8/IL-8 production. We used an ex vivo model of
C. acnes-induced inflammation based on human skin explants from healthy individuals. The stimulation of these explants with
C. acnes strains led to the massive production of TNF-α, GM-CSF and IL-1β. This result is consistent with previous reports of the expression of these cytokines in lesional skin from acne patients [
33,
45]; however, metalloproteinase expression was weak following stimulation with
C. acnes. This discrepancy may be due to the use of normal healthy skin explants in our study and lesional skin explants from acne patients in previous reports. We detected the production of the anti-inflammatory IL-10 cytokine, but at levels far lower than for TNF-α, GM-CSF and IL-1β. This result is consistent with previous findings showing IL-10 production in skin explants from acne lesions and from healthy individuals [
46]. IL-17 expression was weak in this model. This result is consistent with previous reports that
C. acnes can induce the production of IL-17 by PBMCs and in acne lesions, indicating the presence of a CD4
+ T-cell infiltrate. The low level of IL-17 in our model may be due to the low counts of CD4
+ cells and CD83
+ mature dendritic cells in non-lesional skin, as previously reported [
45,
47]. Similarly, only very small amounts of IL-1α were produced, whereas this cytokine has been reported to be produced in large amounts in keratinocytes in vitro and in a mouse model in vivo involving a sebum-like application of acne-related
C. acnes strains [
48,
49]. CXCL8/IL-8 levels were very high in the skin explant culture medium due to the presence of SVF in the medium. We, therefore, considered the values obtained for CXCL8/IL-8 to be uninformative in this model. We found no major difference between the
C. acnes phylotypes in terms of the levels of proinflammatory molecules produced, but only one strain per phylotype was tested. The investigation of larger numbers of
C. acnes strains is therefore required for any firm conclusions to be drawn. Moreover, comparisons between healthy- and acne-related
C. acnes strains are likely to be of interest. This model is not directly comparable to acne lesions, but it could, nevertheless, be useful for investigations of the
C. acnes-induced response, as the skin explant may contain some of the cells involved in the immune response.
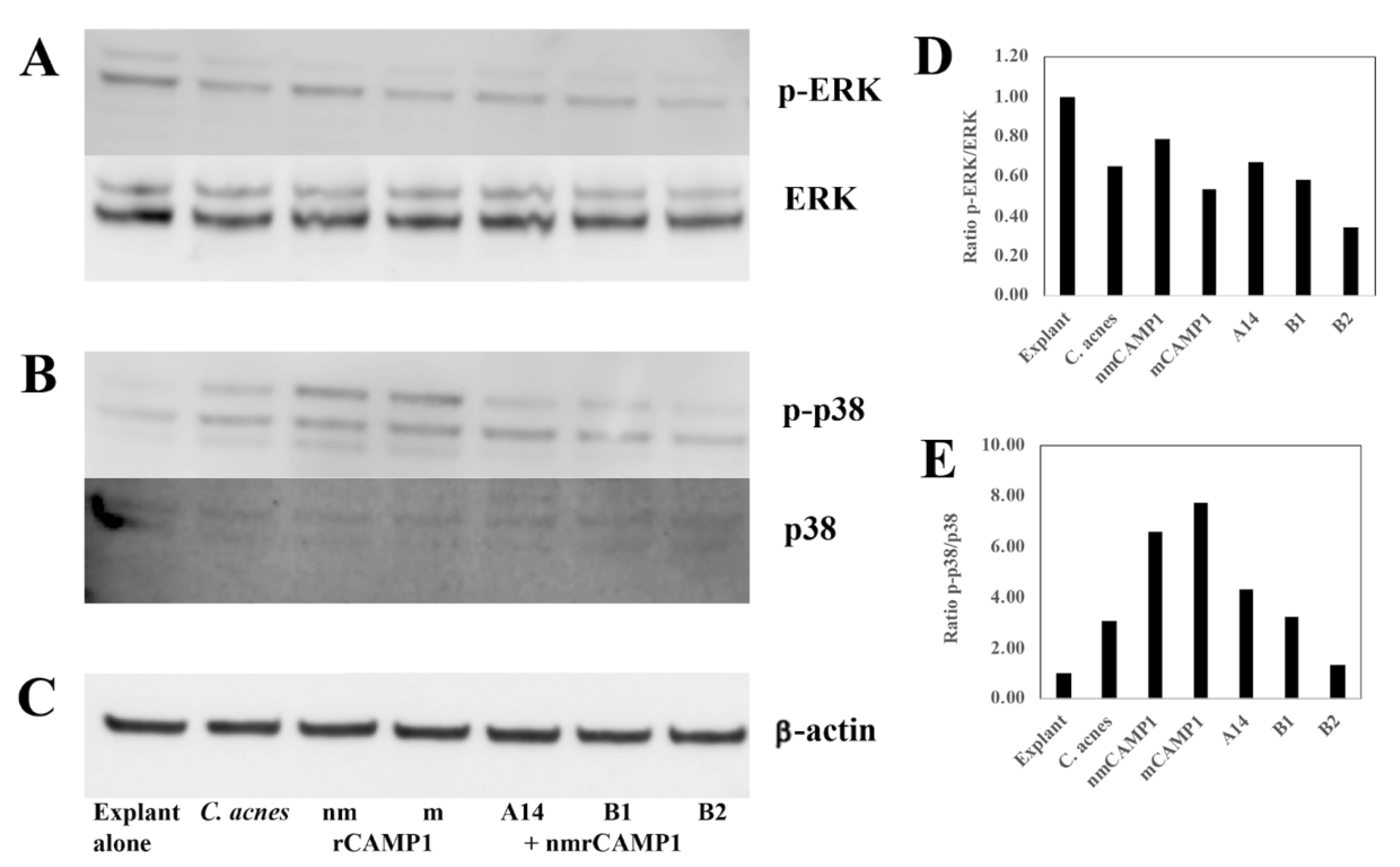
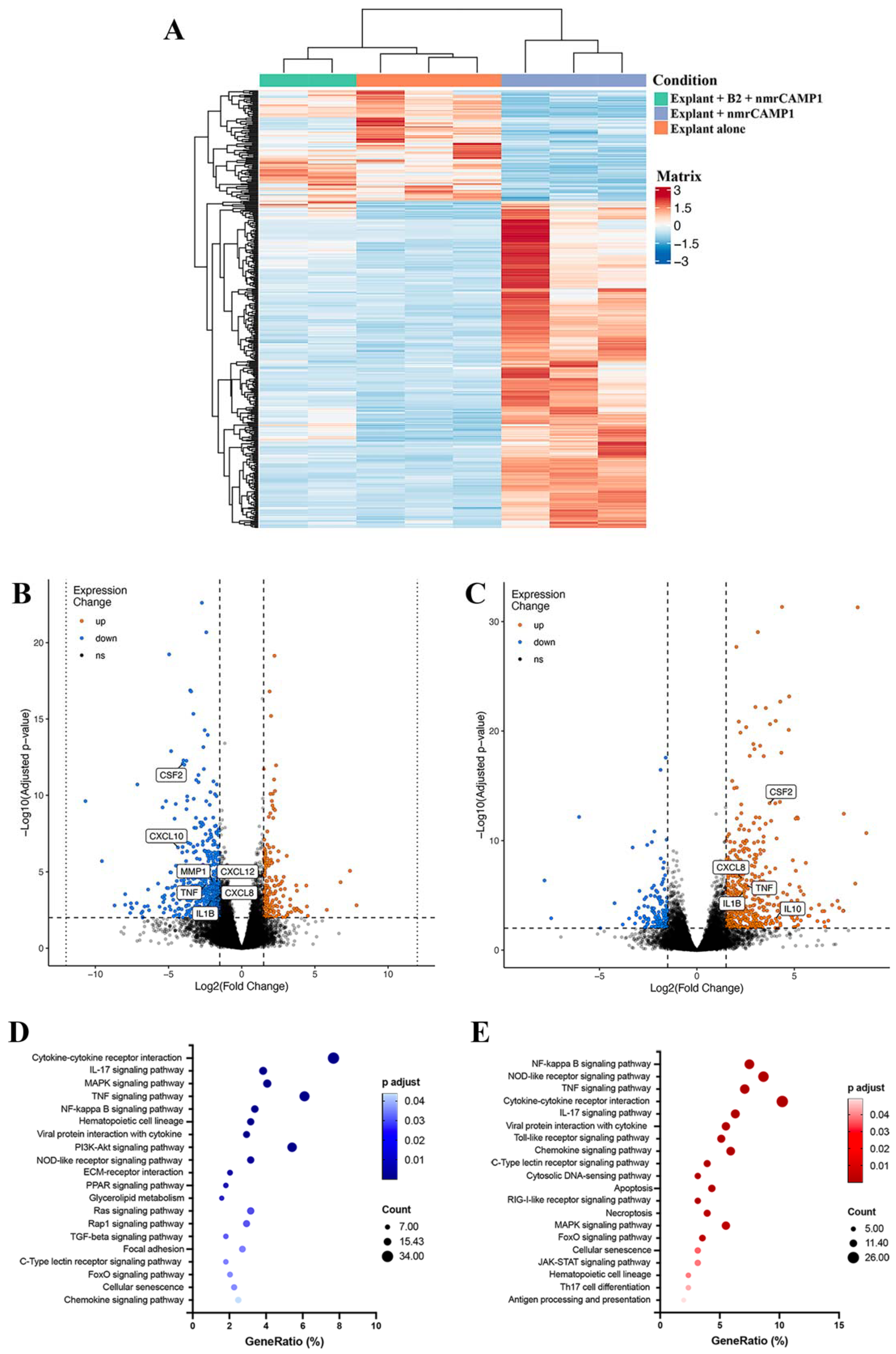
We then assessed the impact of peptides A14, B1 and B2 on skin explants stimulated with both
C. acnes and nmrCAMP1. These explants displayed significantly lower levels of TNF-α, GM-CSF and IL-1β production and also had significantly lower levels of the metalloproteinases MMP1 and MMP3. Metalloproteinases are enzymes with proteolytic activity that are involved in the control of extracellular matrix (ECM) remodeling [
50]. They are induced by reactive oxygen species (ROS) produced in response to several stresses, including UV exposure, TNF-α [
51,
52] and
C. acnes infection [
53]. The ROS activate the MAPK pathway, thereby activating the transcription factor activator protein-1 (AP-1), leading to the MMP expression [
54]. MMPs play a key role in acne scarring and
C. acnes upregulates several of these enzymes [
55]. CAMP1-derived peptides, therefore, seem to be able to downregulate all the major inflammatory components induced by
C. acnes. We further investigated the impact of the B2 peptide on the inflammatory response induced by
C. acnes, by performing an RNA sequencing analysis on skin explants treated with the B2 peptide and stimulated with nmrCAMP1. Most of the inflammatory markers strongly upregulated in the presence of nmrCAMP1 were downregulated in the presence of B2. We identified 12 pathways significantly affected in both untreated and treated conditions: the cytokine–cytokine receptor interaction, the IL-17 signaling pathway, the TNF signaling pathway, the MAPK signaling pathway, the NF-κB signaling pathway, the hematopoietic cell lineage, the viral protein interaction with cytokine and cytokine receptor, the NOD-like receptor signaling pathway, the C-type lectin receptor signaling pathway, the forkhead (Fox)O signaling pathway, cellular senescence and the chemokine signaling pathway. This finding is consistent with previous reports identifying the cytokine–cytokine receptor interaction pathway as active in acne lesions, supporting the notion that this model is a potentially interesting tool for studies of the innate immune molecular pathways induced by
C. acnes [
56,
57]. We identified 10 DEGs from these pathways:
IL6,
TNF,
CXCL1,
CXCL2, CXCL3, CXCL8, IL-1β, CCL2, CCL4 and CSF2. All these genes encode proinflammatory cytokines and chemokines previously detected in acne lesions [
58] and are linked to activation of the NF-κB pathway, inducing the upregulation of TNF-α, CXCL8/IL-8, IL-1β, CXCL1 and CXCL2 [
16,
59,
60]. The B2 peptide decreases the inflammatory response induced by
C. acnes and can therefore be considered a novel modulator of the TLR-2 signaling pathway. The use of an ex vivo human skin explant model also appears to be a useful tool for studies of the impact of candidate anti-inflammatory molecules.
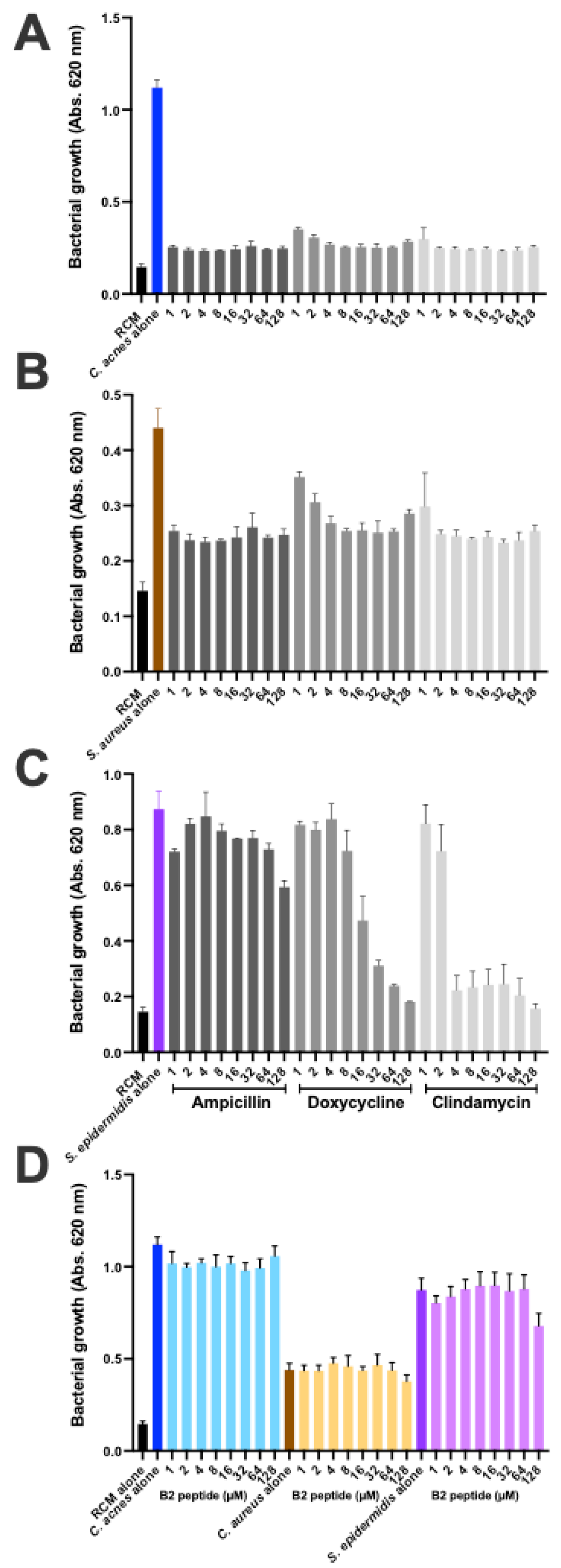
The cutaneous microbiota has been shown to play an essential role in maintaining skin homeostasis and health, with imbalances leading to several diseases.
C. acnes is by far the most abundant of the cutaneous bacteria and the selection of a specific acne-associated lineage may be linked to skin dysbiosis [
61,
62]. The increase in antibiotic resistance in
C. acnes and other skin-related bacteria is making it necessary to develop and test new therapeutic approaches. Peptide-based approaches, in particular, are being considered; however, this technology is based essentially on disruption of the membrane to decrease bacterial viability and the associated inflammatory reaction [
63,
64,
65]. We show here that the CAMP1-related B2 peptide did not alter the growth of
C. acnes,
S. aureus and
S. epidermidis strains at high concentrations. It has been shown that TLR-2 modulators can be used to treat acne without adverse events [
66]. The B2 peptide derived from the
C. acnes CAMP1 protein, which has strong anti-inflammatory properties, is, therefore, a good candidate for clinical testing.
4. Materials and Methods
4.1. Bacterial Strain and Conditions of Growth
We used 24 strains of
C. acnes in the study: strain 6919 (Type IA
1) from the American Type Culture Collection (Manassas, VA) and strains isolated from various infections (13 bone and joint infections, 1 case of pleuritis, 1 case of appendicitis, 1 case of pericarditis, 1 case of pneumonia, 3 cutaneous infections, 1 case of bacteremia and 2 ascitic fluid infections).
C. acnes strains RON (IA
1), TRI (IA
2), LIE (IA
2), CHR (IA
2), GUE (IB) and PIE (IB) were isolated from patients with joint infections. All these strains were kindly provided by Dr. Philippe Morand from the Bacteriology Department at Cochin Hospital, Paris, France [
24].
C. acnes strains were grown under anaerobic conditions in liquid or solidified reinforced clostridial medium (RCM; Difco Laboratories, Detroit, MI, USA), in a GasPak™ EZ Anaerobic Container System (Becton Dickinson & Co, Franklin Lakes, NJ, USA) at 37 °C. For routine culture, 100 mL of RCM was used and the bacteria were harvested after 5 days at 37 °C, by centrifugation at 7,000 x
g for 10 min and 4 °C. The pellets were pooled and washed in about 30 mL cold sterile phosphate-buffered saline (PBS) (1.5 mM KH
2PO
4, 2.7 mM Na
2HPO
4.7H
2O, 0.15 M NaCl (pH 7.4)), and were then centrifuged again, as described above. The final bacterial pellet was suspended in sterile PBS (1:10 based on the volume of the culture).
Staphylococcus aureus (ATCC #25923) and
Staphylococcus epidermidis (ATCC #12228) were grown in tryptic soy broth (TSB) and brain heart infusion (BHI) (Sigma-Aldrich, Saint-Louis, MO, USA) under aerobic conditions at 37 °C.
Two chemically competent Escherichia coli strains (TOP10 and BL21 StarTM DE3; Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) were used. The strains were grown under aerobic conditions at 37 °C in liquid or solidified Luria-Bertani (LB) (Sigma-Aldrich, Saint-Louis, MO, USA) medium with or without ampicillin (50 μg/mL) to screen for transformants. Super optimal broth with catabolite repressor (SOC medium) was used for cell recovery after heat shock during the transformation experiments.
4.2. Total Bacterial Surface Protein Extraction
The
C. acnes total protein extract was obtained by incubating the bacterial pellet with chicken egg lysozyme (2 mg/mL) for 1 h at 37 °C, with gentle mixing. The suspension was centrifuged at 10,000×
g for 15 min at 4 °C and the supernatant containing the proteins was retained. Protein concentration was determined by the Lowry method, with BSA as the standard [
67].
4.3. Production of Polyclonal anti-CAMP1 Antibodies
Antibodies directed against CAMP1 (accession number AAS92206.1) were obtained by conjugating two peptides with keyhole limpet hemocyanin (carrier protein used as a vaccine adjuvant): KMPDLKPNDVAT and VLRQIRFDRNTC. The resulting conjugates were injected into female New Zealand White rabbits (Proteogenix, Schiltigheim, France). Three injections were performed over a period of 70 days. Blood was obtained from the rabbits at the end of the immunization protocol. Antisera were first depleted against immobilized non-phosphorylated peptide, and specific antibodies against CAMP1 were then purified with the phosphorylated KMPDLKPNDVAT and VLRQIRFDRNTC peptides immobilized on a column. The titer of purified antibodies was evaluated by enzyme-linked immunosorbent assay (ELISA) with the phosphorylated and non-phosphorylated peptides and was found to be 1:64,000 (
Figure S1).
4.4. Far-Western Blotting
C. acnes surface proteins (50 μg) were separated by electrophoresis under denaturing conditions in a NuPAGE Novex 4–12% Bis-Tris gel (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France). The separated proteins were transferred onto a nitrocellulose membrane, which was then incubated in saturation buffer containing 1 X tris-buffered saline (TBS), 1.4 M NaCl, 5% non-fat milk, 0.1% Tween 20, 200 mM Tris (pH 7.6) for 1 h at room temperature. The membrane was washed three times, for 15 min each, with TBS/T buffer (1 X TBS, 0.1% Tween-20) and were then incubated overnight with human recombinant TLR-1, TLR-2, TLR-4 or TLR-6 (R&D Systems, Abingdon, UK) diluted to 0.1 μg/mL in TBS/T, at 4 °C, with gentle shaking. The unbound protein probe was removed by washing as described above, and the membranes were incubated with biotinylated human antibodies against TLR-1, TLR-2, TLR-4 or TLR-6 (R&D System, Abingdon, UK) diluted to 0.05 μg/mL in TBS/T supplemented with 5% BSA, for 20 h at 4 °C, with gentle shaking. The membranes were washed to remove unbound antibodies, and the surface protein-TLR1,2,4 or 6-biotinylated antibody complexes were detected by incubation for 1 h at room temperature with HRP (horseradish peroxidase) diluted to 0.5 μg/mL in saturation buffer. Unbound material was removed by washing and peroxidase activity was detected in a chemiluminescence assay (WesternBright ECL, Advansta, Menlo Park, CA, USA).
4.5. CAMP1 Binding Activity
For quantitative analysis, nmrCAMP1 and mrCAMP1 proteins were diluted to 20 μg/mL in 50 mM carbonate–bicarbonate buffer (pH 9.6) and immobilized on 96-well polystyrene plates by overnight incubation at 4 °C. The wells were washed three times with 0.05% Tween-20 in PBS (PBS-Tween). Human recombinant TLR-1, TLR-2, TLR-4 or TLR-6 (R&D System, Abingdon, UK) (0.01 to 16 μg/mL in PBS-Tween) was added and the plates were incubated overnight at 4 °C. The plates were washed three times with PBS-Tween, and biotinylated human antibodies against TLR-1, TLR-2, TLR-4 or TLR-6 (R&D System, Abingdon, UK) diluted to 0.05 μg/mL in PBS-Tween were added and the plates were incubated at room temperature for 1 h. The plates were washed, peroxidase-conjugated streptavidin (0.5 μg/mL in PBS-Tween) was added and the plates were incubated for 30 min at room temperature. After washing, bound peroxidase activity was detected by incubation with the chromogenic peroxidase substrate ABTS. Absorbance was measured at 405 nm. For the binding inhibition experiments, nmrCAMP1 was immobilized on a plate at 20 μg/mL as described above. In parallel, TLR-2 (0.1 μg/mL) was incubated overnight at 4 °C with various concentrations of the A14, A15, B1, B2, B3, C1 or C3 (1.25, 12.5, 125, 1250 and 12500 nM) peptides, which were added to the plate after it had been washed with PBS-Tween to remove excess nmrCAMP1. Binding was then detected as described above. The peptides (chemically synthesized by Proteomic Solutions, Saint Marcel, France) were dissolved in 30% DMSO for A14 and A15 and in water for B1, B2, B3, C1 and C3.
4.6. Cell Culture, Pretreatment and Stimulation
Immortalized human keratinocytes HaCaT (Catalog #300493; GmbH, CLS Cell Lines Services GmbH, Eppelheim, Germany) were grown in Dulbecco’s modified Eagle’s medium-Glutamax-I (DMEM) supplemented with 1 mM sodium pyruvate. The immortalized human monocytic cell line ThP1 (ATCC, Manassas, VA, USA; catalog number TIB-202) was grown in Roswell Park Memorial Institute 1640 Medium-Glutamax-I (RPMI). DMEM and RPMI were supplemented with 0.1% and 10% heat-inactivated fetal calf serum and with an antibiotic/antimycotic solution (10 U/mL penicillin, 10 µg/mL streptomycin, 0.25 μg/mL amphotericin; Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) and were placed at 37 °C in a humidified atmosphere containing 5% CO2. Primary normal human epidermal keratinocytes (NHEK, catalog #00192627) were grown in the KGMTM-Gold Bullet Kit, in accordance with the manufacturer’s instructions (Lonza, Basel, Switzerland). Immortalized cell lines were routinely tested to check for an absence of Mycoplasma infection. Cells cultured in six- or 96-well polystyrene plates were subjected to pretreatment with peptides or antibodies for 24 h in the dark, at 37 °C. The medium was replaced and the cells were stimulated with a C. acnes suspension (MOI = 15), or with the recombinant proteins (nmrCAMP1 or mrCAMP1) for 18 h at 37 °C, under an atmosphere containing 5% CO2.
4.7. Western Blot
After peptide pretreatment (62.5 μM) for 24 h, followed by stimulation with a
C. acnes suspension (OD
620 nm = 1.0) or the recombinant proteins at a concentration of 10 μg/mL for 18 h, human skin explants were washed in cold sterile PBS. They were shredded for 2 × 1 min with a gentleMACS
TM Dissociator in a gentleMACS
TM M tube (Miltenyi Biotec, Bergisch Gladbach, Germany) containing RIPA lysis buffer consisting of 50 mM Tris, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS (pH 8.0), 2 mM EDTA, 2 mM sodium pyrophosphate, 10% glycerol, 1 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride and 10 mg/mL leupeptin. Total protein extract was centrifuged at 10,000×
g for 15 min at 4 °C and protein concentration was determined by the Lowry method, with BSA as the standard, as described by Peterson [
67]. Proteins (30 μg per lane) were fractionated by electrophoresis under denaturing conditions in a NuPAGE Novex 4–12% Bis-Tris gel (1 mm, 10 wells, Invitrogen, UK) and the resulting bands were transferred to nitrocellulose membranes. Membranes were saturated by incubation for 1 h at room temperature in 5% nonfat dry milk in TBS/T buffer (1X TBS, 0.1% Tween-20), and were then incubated for 18 h at 4 °C with rabbit polyclonal IgG antibodies against p-p38 (Catalog #sc-17852-R, Thr 180/Tyr 182, 1:500 BSA), p38 (Catalog #sc-535, C-20), or ERK1 (Catalog #sc94, K-23), with mouse monoclonal IgG antibodies against native and phosphorylated forms of p-ERK (Catalog #sc-7383, E-4), and β-actin (Catalog #sc-47778, C4) used as protein loading controls after dilution to 1:500 in 5% BSA in TBS/T buffer (all antibodies were purchased from Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA otherwise stated). The membranes were washed and bound antibodies were detected by incubation for 1 h at room temperature with polyclonal goat anti-mouse IgG-horse radish peroxidase (HRP) (Catalog #sc-2005) or goat anti-rabbit IgG-HRP (Catalog #sc-2004), antibodies diluted to 1:2000 in saturation buffer. Unbound material was removed by washing, and peroxidase activity was detected by chemiluminescence with ECL Western Blotting Reagent (Advansta Corp., Menio Park, CA, USA).
4.8. Bacterial DNA Extraction
Bacterial DNA was extracted with the E.Z.N.A. Bacterial DNA kit, in accordance with the manufacturer’s recommendations (Omega Bio-Tek, Norcross, GA, USA). The samples were treated with lysozyme to digest the cell wall, and then with protease and RNase. The binding conditions were adjusted and the samples were transferred to spin columns, specifically binding DNA. The columns were washed, the DNA was eluted and its concentration was determined by measuring absorbance at 260 nm (Nanodrop 2000, Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France). The values obtained were in the range of 30 to 220 ng/μL, and the concentration of the solution was adjusted as required.
4.9. Production and Purification of the His-Tag Fusion CAMP1 Proteins
We used a phylotype IA1
C. acnes strain (75150) expressing the non-mutated CAMP1 protein (nmCAMP1) (GenBank Accession number KX581400), and a phylotype II
C. acnes strain (27647) expressing the mutated CAMP1 protein (mCAMP1) (GenBank Accession number KX581395) [
24]. The gene encoding the CAMP1 factor protein was, in each case, amplified with specific CAMP1F1 (forward and reverse) primers (
Table S1) and the
Pfx DNA polymerase in a thermocycler ProFlex PCR System (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France), as follows: initial denaturation at 98 °C for 30 s, 35 cycles of denaturation at 98 °C for 10 s, annealing at 65 °C for 10 s and extension at 72 °C for 30 s. A final extension cycle was performed at 72 °C for 10 min. The CACC sequence was added at the start of the forward primer, to direct the insertion of blunt-ended PCR products into the TOPO cloning vector in accordance with the manufacturer’s instructions (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France). Both amplicons were purified and inserted into the pET102D/TOPO expression vector (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France), according to the manufacturer’s instructions, to obtain the vectors pET102-nmCAMP1 and pET102-mCAMP1. For amplification and storage, both recombinant plasmids were introduced into the chemically competent
E. coli TOP10 strain and selected on the basis of their resistance to ampicillin added to the LB medium (50 μg/mL). Both plasmids were sequenced on a 3730xl DNA Analyzer (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) to confirm their sequences. Both recombinant plasmids had the correct sequence and were, therefore, introduced into the BL21 Star
TM (DE3)
E. coli strain for overexpression of the recombinant proteins, nmrCAMP1 and mrCAMP1, with a poly-histidine (6-His) tag at their N-termini. Transformed bacteria were cultured overnight in 1 L of LB medium supplemented with 1% glucose and 50 μg/mL ampicillin. The overnight culture was then used to inoculate 6 L of Overnight Express
TM Instant LB medium (Merck, Darmstadt, Germany), which was incubated at 37 °C for four hours for protein production. Bacteria were harvested by centrifugation at 5000×
g for 10 min and the pellet was resuspended in Lysis Equilibration Buffer (50 mM NaH
2PO
4, 300 mM NaCl, pH 8.0 and 1 mg/mL lysozyme). The suspension was subjected to 30-s pulses (10 × 30 bursts with a 30 s period between bursts) of sonication on ice. The lysate was centrifuged for 30 min at 10,000×
g and the pellet was suspended in a denaturing solubilization buffer (DNS) containing 50 mM NaH
2PO
4, 300 mM NaCl and 8 M urea, pH 8.0. The suspension was subjected to sonication (10 pulses of 30 s each), and the resulting lysate was clarified by centrifugation for 30 min at 10,000×
g. The clarified lysate, containing recombinant proteins, was loaded onto a Protino NI-TED resin immobilized metal chelate affinity chromatography (IMAC) column (GE Healthcare, Chicago, IL, USA). The column was washed with the DNS buffer, and the proteins were eluted in a buffer containing 50 mM NaH
2PO
4, 300 mM NaCl, 8 M urea, 250 mM imidazole pH 8.0. The fractions containing the recombinant proteins were analyzed by SDS-PAGE and stored at −20 °C. Before use, recombinant proteins were thoroughly dialyzed against PBS to remove urea and imidazole (
Figure S2).
4.10. Peptide Array Analysis
For identification of the peptide sequences involved in binding to TLR-2, the amino acid sequences of nmCAMP1 (GenBank Accession number AAS92206.1) and mCAMP1 (GenBank Accession number KX581395) were split into 15-amino acid peptides, with overlapping frames of 11 amino acids, to cover the entire protein sequence. Each peptide was immobilized on a glass slide containing a total of 116 spots (A1 to E20). We also included a positive control corresponding to an unrelated biotinylated peptide (F22) and two negative controls corresponding to unrelated peptides: a V5 epitope tag (F23) and a FLAG tag (F24) (
Table S2) (Active Motif, Carlsbad, CA, USA). The glass slide was incubated in a saturation buffer containing 4 M NaCl, 5% nonfat milk, 0.1% Tween 20, 200 mM Tris (pH 7.6) in TBS for 1 h. The slide was washed three times, for 15 min each, in TBS/T buffer (1 X TBS, 0.1% Tween-20), and incubated overnight at 4 °C with human recombinant TLR-2 (R&D System, Abingdon, UK) diluted at 0.1 μg/mL in TBS/T, with gentle shaking. Unbound antibodies were removed by washing, as described above, and slides were incubated with biotinylated human antibodies against TLR-2 (R&D Systems, Abingdon, UK) diluted to 0.05 μg/mL in TBS/T supplemented with 5% BSA, for 20 h at 4 °C, with gentle mixing. After washing to remove unbound antibodies, the surface protein-TLR-2-biotinylated antibody complexes were detected by incubation for 1 h at room temperature with HRP diluted to 0.5 μg/mL in saturation buffer. Unbound material was removed by washing and peroxidase activity was detected in a chemiluminescence assay (WesternBright ECL, Advansta, Menlo Park, CA, USA).
4.11. In Silico CAMP1-3D Structure Modeling
The comparative modeling approach was used to obtain an accurate 3D structure for the nmCAMP1 and mCAMP1 proteins, with the 3D structures of these proteins predicted based on sequence alignments with known protein models in the Protein Data Bank (PDB) database. Reasonable predictions of the structures of these proteins were generated with trRosetta (
https://yanglab.nankai.edu.cn/trRosetta/ accessed on 19 March 2021) [
68]. All the figures were produced with the molecular visualization system PyMOL [
69], Available from:
http://www.pymol.org/pymol accessed on 14 January 2021).
4.12. Cell Viability Assays
Cell viability was estimated in the (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide (MTT) assay, in which cells were incubated with a 0.2% MTT solution in a cell culture medium for 4 h at 37 °C. This solution was removed and dimethyl sulfoxide (DMSO) was added to solubilize the MTT-formazan compound produced in living cells. Absorbance was measured at 550 nm.
4.13. Ex Vivo C. acnes-Induced Inflammation Assay
Full-thickness human skin was obtained as excess tissue from abdominoplasty procedures at the De La Tour Clinique in Paris, France. The tissue was collected in an anonymous manner, in accordance with routine procedures approved by the local ethics committee. The subcutaneous fat was trimmed off the skin tissue and punch biopsies (8 mm) were performed on the skin. The discs obtained were immediately placed in DMEM/RPMI/SVF (50/50/20) culture medium containing 2% penicillin/streptomycin and 1% fungicide, and complemented with 250 μg/mL insulin, 0.1 mg/mL epidermal growth factor (EGF) and 0.1 mg/mL fibroblast growth factor (FGF). They were incubated at 37 °C under an atmosphere containing 5% CO2 for 18 h, to verify for an absence of bacterial contamination. Skin explants were then incubated with 62.5 μM peptide for 24 h at 37 °C and were then stimulated with a C. acnes suspension (OD620 nm = 1.0) or the nmrCAMP1 protein (10 μg/mL) for 18 h at 37 °C. A baseline control was established with untreated and unstimulated skin explants. The positive control was a skin explant stimulated with C. acnes suspension or with nmrCAMP1 protein only.
4.14. Enzyme-Linked Immunosorbent Assay (ELISA)
Human CXCL8/IL-8 and TNF-α protein concentrations were measured in the supernatants of stimulated cells or skin explants with Ready-Set-Go ELISA Sets (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) according to the manufacturer’s instructions. We used serial dilutions of recombinant human CXCL8/IL-8 and TNF-α for the establishment of standard curves. Optical density at 450 nm was determined with a wavelength correction of 570 nm.
4.15. Luminex Assay
ProcartaPlex human 9-plex assays (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) were used to determine cytokine levels (GM-CSF, CXCL8/IL8, IL-10, IL-17A, IL-1α, IL-1β, IL-12, MMP-1, MMP-3, TNF-α) in the supernatant of NHEK and human skin explants stimulated with C. acnes and nmrCAMP1 protein, according to the manufacturer’s specifications.
4.16. RNA Isolation and Quantitative Real-Time PCR
Cells and skin explants were grown in 6- or 96-well plates. They were incubated for 24 h with peptides (62.5 µM) or antibodies (1 μg/mL) and were then stimulated by incubation for 18 h with
C. acnes or nmrCAMP1 (10 μg/mL), as previously described. Total RNA was isolated with TRIzol (Invitrogen, Carlsbad, USA) and the RNeasy
® Mini kit and was treated with DNAse I, according to the manufacturer’s instructions (Qiagen, Hilden, Germany). RNA concentration was determined by measuring absorbance at 260 nm on a Nanodrop spectrometer (Labtech, France) and the A
260/A
280 ratios for all samples were between 1.6 and 1.9. Complementary DNA was generated from 50 ng of total RNA by reverse transcription at 50 °C for 10 min, and was then subjected to quantitative PCR analysis in a QuantStudio™ 5 Real-Time PCR System thermocycler (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) with the iTaq Universal SYBR Green One-Step kit (Bio-Rad Laboratories, Hercules, CA, USA), with two-step cycle conditions as follows: 95 °C for 60 s followed by 40 cycles of 95 °C for 15 s, 68 °C for 60 s, and a final melting curve at 65–95 °C, for 60 s with increments of 0.1 °C/s. The threshold cycle (Ct) values for the genes studied were determined from the amplification curves. The amount of RNA in stimulated cells relative to control cells was calculated by the 2ΔCt method and is expressed as a relative fold change in expression normalized against the expression of an internal control gene (
GAPDH). All the primers used in this study are described in
Supplementary Table S3.
4.17. RNA Sequencing Analysis
Human skin explants were left untreated and unstimulated (negative control group), were stimulated with nmrCAMP1 (10 μg/mL) for 24 h at 37 °C (positive control group) or were treated with the CAMP1-related B2 peptide (62.5 μM) for 24 h at 37 °C and then stimulated with nmrCAMP1 (treated group). Total RNA was extracted with TRIzol (Invitrogen, Carlsbad, USA) and the RNeasy
® Mini kit and was treated with DNAse I, according to the manufacturer’s instructions (Qiagen, Hilden, Germany). We used 250 ng of RNA to generate the cDNA library with the Illumina Truseq mRNA prep ligation kit. RNAseq was performed on an Illumina NovaSeq instrument providing 50 bp paired-end reads. The reads (about 26 to 34 million reads per sample) in the fastq files were aligned with the GRCh38.101 version of the human genome with RSEM (v1.3.3), using STAR aligner (v2.7.6a). Differential expression between the two sample groups was analyzed with the DESeq2 package (
https://github.com/BSGenomique/genomic-rnaseq-pipeline/releases/tag/v1.0420 accessed on 22 October 2021) with an adjusted
p-value threshold of 0.05. RNA sequencing results were analyzed with the GENOM’IC’s application (
https://github.com/GENOM-IC-Cochin/shiny-rnaseq-viz accessed on 26 November 2021). (GENOM’IC platform, Institut Cochin, U1016, Paris, France). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were performed for the upregulated and downregulated genes with the Database for Annotation, Visualization, and Integrated Discovery (DAVID) (
https://david.ncifcrf.gov/ accessed on 21 March 2022). For IL-6, CXCL1, CXCL2, CXCL3, CCL3 and CCL4, differential expression was confirmed by qRT-PCR with TaqMan probes labeled with FAM fluorochromes. Amplifications were performed in the presence of 50 ng cDNA (previously reverse transcribed with the PreSuperScript™ IV VILO™ Master Mix and ezDNase™ Enzyme), 10 μL TaqMan™ Fast Advanced Master Mix, 1 μL TaqMan™ assay 20X (ACTB: Hs99999903_m1, IL-6: Hs00985639_m1, CXCL1: Hs00236937_m1, CXCL2: Hs00236966_m1, CXCL3: Hs00171061_m1, CCL3: Hs00234142_m1 or CCL4: Hs99999148_m1), in a final volume of 20 μL (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France). The reaction was performed on a QuantStudio™ 5 Real-Time PCR System thermocycler (Thermo Fisher Scientific, Life Technologies SAS, Courtaboeuf, France) with an initial UNG incubation for 2 min at 50 °C, one polymerase activation step at 95 °C for 20 s, followed by 40 cycles of denaturation for 1 s at 95 °C, annealing and extension for 20 s at 60 °C. The same method as described above was used to determine the fold-change in expression relative to the internal control ACTB.
4.18. Statistical Analysis
The significance of differences between experimental groups was determined in parametric unpaired t-tests and ANOVA, implemented in GraphPad Prism 9 (GraphPad, La Jolla, CA, USA). A p-value ≤ 0.05 was considered statistically significant, and significance is indicated as follows: * (p ≤ 0.05), ** (p ≤ 0.01), *** (p ≤ 0.001) and **** (p ≤ 0.0001).


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}