
Preclinical In Vitro Model to Assess the Changes in Permeability and Cytotoxicity of Polarized Intestinal Epithelial Cells during Exposure Mimicking Oral or Intravenous Routes: An Example of Arsenite Exposure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. Chemicals
2.3. Treatment Concentration of Arsenic Species
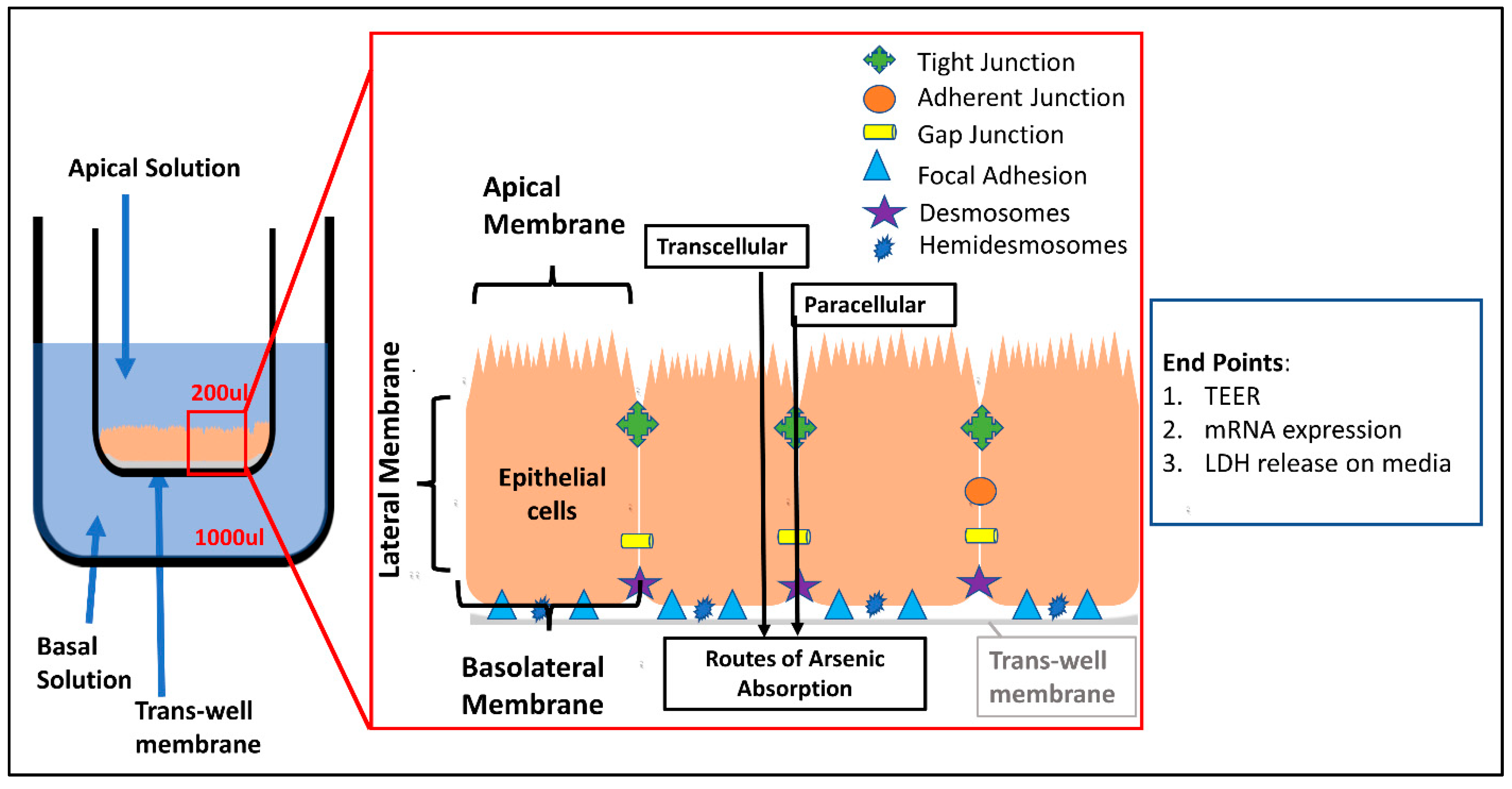
2.4. Culture of T-84 Cells and Trans-Epithelial Electrical Resistance (TEER) Study
2.5. Analysis of Trans-Epithelial Electrical Resistance
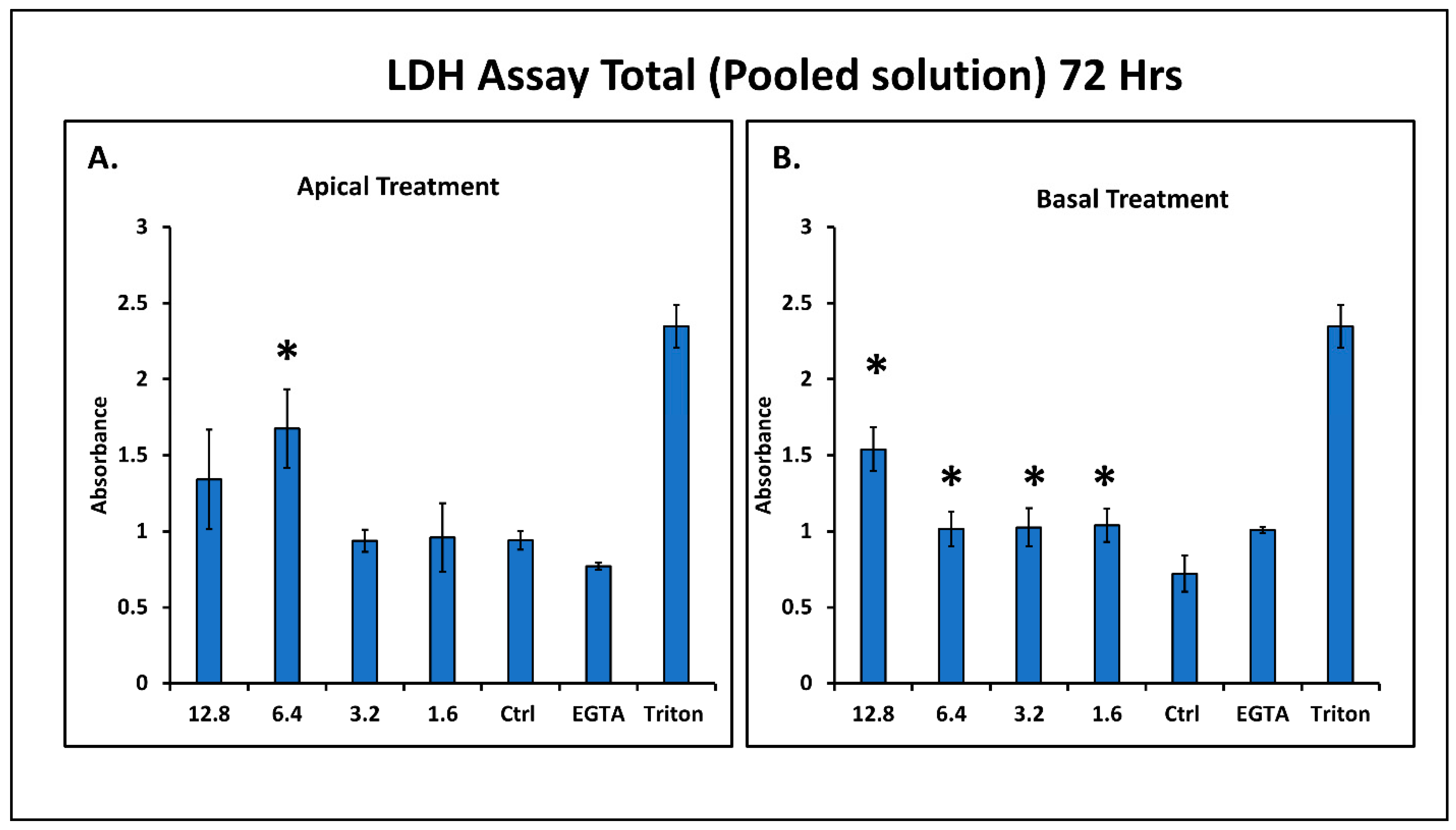
2.6. Lactate Dehydrogenase (LDH) Assay
2.7. Extraction of mRNA, DNAse Treatment and cDNA Conversion T-84 Cells
2.8. Determination of Cell-Junction-Related Gene Expression Using qPCR
3. Statistical Analysis
4. Results
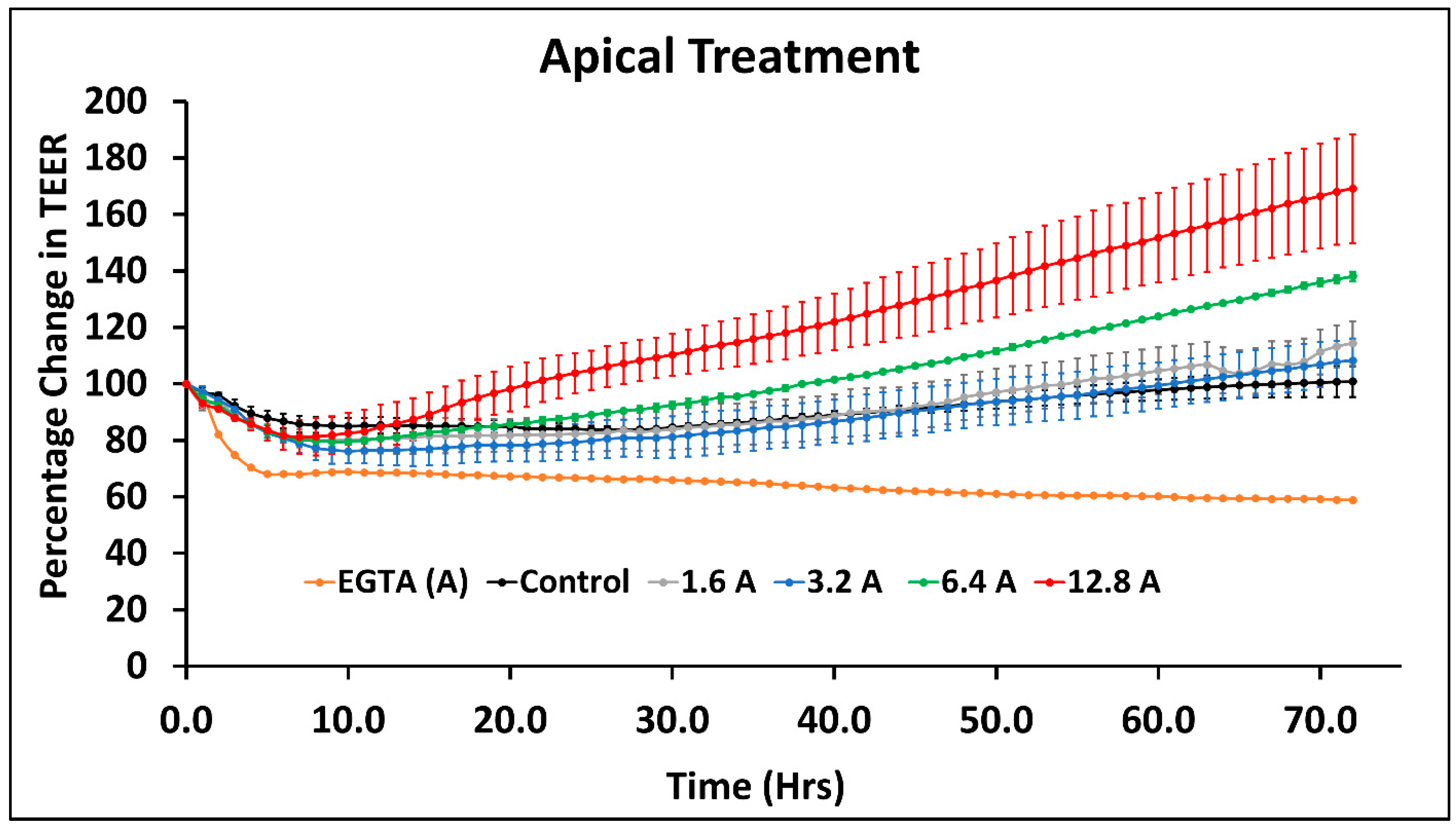
4.1. Effect of Arsenic Species on the Intestinal Cell Monolayer Permeability
4.2. Effect of Sodium Arsenite on T-84 Cell-Junction-Related Genes When Exposed from Apical Side
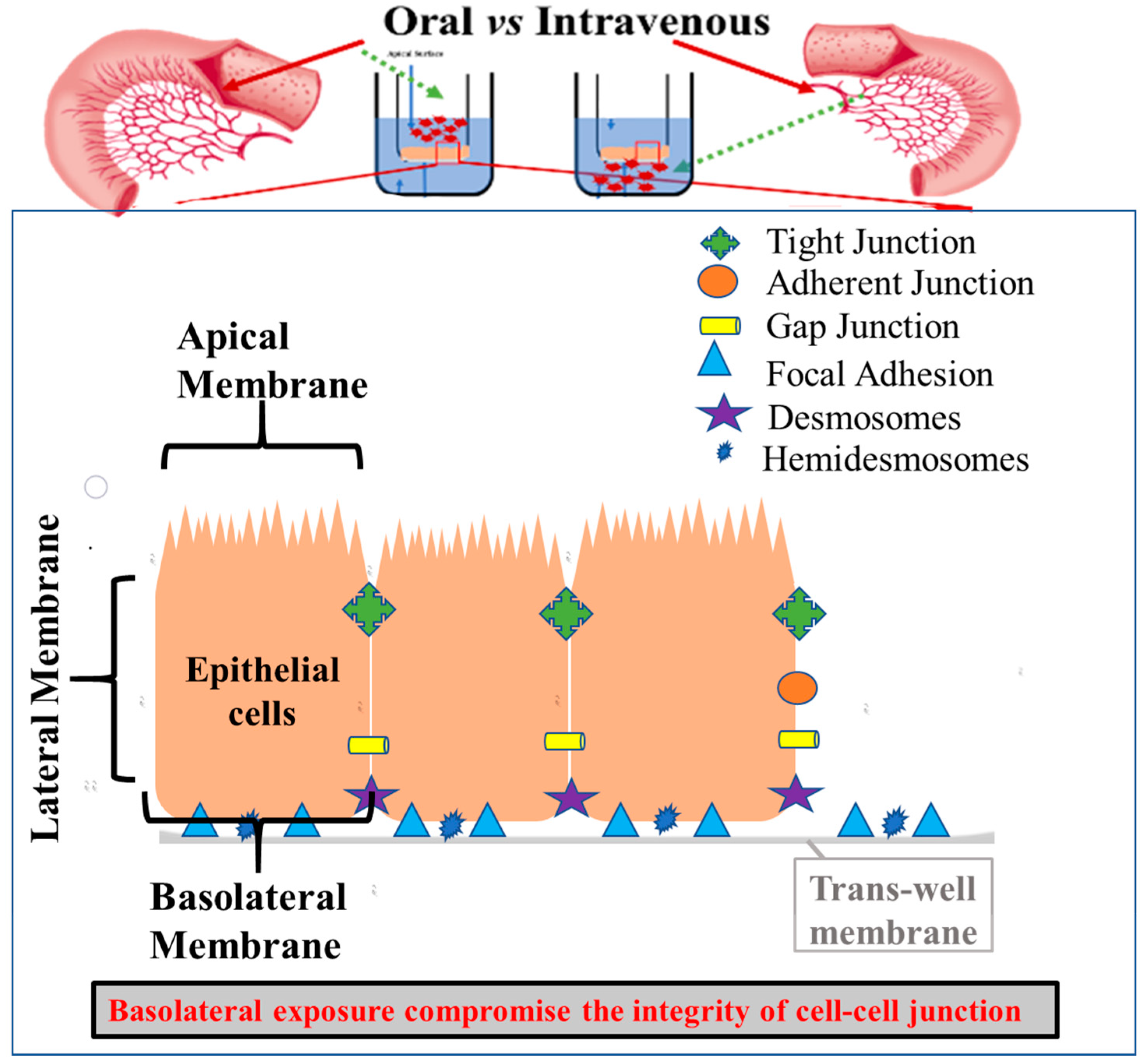
4.3. Effect of Sodium Arsenite Exposure on T-84 Monolayer and Change in Cell-Junction-Related Gene Expression
4.4. Effect of Sodium Arsenite on Cell Cytotoxicity
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease pRev.ention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhadi, A.; Banan, A.; Fields, J.; Keshavarzian, A. Intestinal barrier: An interface between health and disease. J. Gastroenterol. Hepatol. 2003, 18, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Konig, J.; Wells, J.; Cani, P.D.; Garcia-Rodenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human Intestinal Barrier Function in Health and Disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef] [PubMed]
- Schoultz, I.; Keita, A.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef]
- Wang, L.; Llorente, C.; Hartmann, P.; Yang, A.M.; Chen, P.; Schnabl, B. Methods to determine intestinal permeability and bacterial translocation during liver disease. J. Immunol. Methods 2015, 421, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Twaddle, N.C.; Vanlandingham, M.; Beland, F.A.; Doerge, D.R. Metabolism and disposition of arsenic species from controlled dosing with dimethylarsinic acid (DMA(V)) in adult female CD-1 mice. V. Toxicokinetic studies following oral and intravenous administration. Food Chem. Toxicol. 2019, 130, 22–31. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, J.; Lu, X.; Gong, Z.; Le, X.C. Arsenic speciation in urine from acute promyelocytic leukemia patients undergoing arsenic trioxide treatment. Chem. Res. Toxicol. 2004, 17, 95–103. [Google Scholar] [CrossRef]
- Kile, M.L.; Houseman, E.A.; Baccarelli, A.A.; Quamruzzaman, Q.; Rahman, M.; Mostofa, G.; Cardenas, A.; Wright, R.; Christiani, D.C. Effect of prenatal arsenic exposure on DNA methylation and leukocyte subpopulations in cord blood. Epigenetics 2014, 9, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Devesa, V.; Adair, B.M.; Liu, J.; Waalkes, M.P.; Diwan, B.A.; Styblo, M.; Thomas, D.J. Arsenicals in maternal and fetal mouse tissues after gestational exposure to arsenite. Toxicology 2006, 224, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.S.; Song, K.H.; Chung, J.Y. Health effects of chronic arsenic exposure. J. Prev. Med. Public Health 2014, 47, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, M.; Laparra Llopis, J.M. 10-Arsenic Through the Gastrointestinal Tract. In Handbook of Arsenic Toxicology; Flora, S.J.S., Ed.; Academic Press: Oxford, UK, 2015; pp. 281–299. [Google Scholar]
- Chung, J.Y.; Yu, S.D.; Hong, Y.S. Environmental source of arsenic exposure. J. Prev. Med. Public Health 2014, 47, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Nigra, A.E.; Sanchez, T.R.; Nachman, K.E.; Harvey, D.; Chillrud, S.N.; Graziano, J.H.; Navas-Acien, A. The effect of the Environmental Protection Agency maximum contaminant level on arsenic exposure in the USA from 2003 to 2014: An analysis of the National Health and Nutrition Examination Survey (NHANES). Lancet Public Health 2017, 2, e513–e521. [Google Scholar] [CrossRef] [Green Version]
- Stanton, B.A.; Caldwell, K.; Congdon, C.B.; Disney, J.; Donahue, M.; Ferguson, E.; Flemings, E.; Golden, M.; Guerinot, M.L.; Highman, J.; et al. MDI Biological Laboratory Arsenic Summit: Approaches to Limiting Human Exposure to Arsenic. Curr. Environ. Health Rep. 2015, 2, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Ayotte, J.D.; Medalie, L.; Qi, S.L.; Backer, L.C.; Nolan, B.T. Estimating the High-Arsenic Domestic-Well Population in the Conterminous United States. Environ. Sci. Technol. 2017, 51, 12443–12454. [Google Scholar] [CrossRef] [PubMed]
- Gokulan, K.; Arnold, M.G.; Jensen, J.; Vanlandingham, M.; Twaddle, N.C.; Doerge, D.R.; Cerniglia, C.E.; Khare, S. Exposure to Arsenite in CD-1 Mice during Juvenile and Adult Stages: Effects on Intestinal Microbiota and Gut-Associated Immune Status. mBio 2018, 9, e01418-18. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.G.; Gokulan, K.; Doerge, D.R.; Vanlandingham, M.; Cerniglia, C.E.; Khare, S. A single or short time repeated arsenic oral exposure in mice impacts mRNA expression for signaling and immunity related genes in the gut. Food Chem. Toxicol. 2019, 132, 110597. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.; Goodale, B.; Raab, A.; Schwerdtle, T.; Reimer, K.; Conklin, S.; Karagas, M.R.; Francesconi, K.A. Human exposure to organic arsenic species from seafood. Sci. Total Environ. 2017, 580, 266–282. [Google Scholar] [CrossRef]
- Coryell, M.; Roggenbeck, B.A.; Walk, S.T. The Human Gut Microbiome’s Influence on Arsenic Toxicity. Curr. Pharmacol. Rep. 2019, 5, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Abernathy, C.O.; Calderon, R.L.; Chappell, W.R. Arsenic: Exposure and Health Effects; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Shi, Q.; Ju, M.; Zhu, X.; Gan, H.; Gu, R.; Wu, Z.; Meng, Z.; Dou, G. Pharmacokinetic Properties of Arsenic Species after Intravenous and Intragastrical Administration of Arsenic Trioxide Solution in Cynomolgus Macaques Using HPLC-ICP-MS. Molecules 2019, 24, 241. [Google Scholar] [CrossRef] [Green Version]
- Sasijareonrat, N.; Jahn, N.; Ungprasert, P.; Owattanapanich, W. Efficacy and the Adverse Effects of Oral Versus Intravenous Arsenic for Acute Promyelocytic Leukemia: A Meta-Analysis of Randomized-Controlled Studies. Technol. Cancer Res. Treat. 2020, 19, 1533033820937008. [Google Scholar] [CrossRef]
- Breccia, M.; Foa, R. Intravenous arsenic trioxide and all-trans retinoic acid as front-line therapy for low-risk acute promyelocytic leukemia. Expert Rev. Hematol. 2019, 12, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Xi, S.; Li, X.; Lu, C.; Li, G.; Xu, Y.; Qu, C.; Niu, Y.; Sun, G. Arsenic speciation transported through the placenta from mother mice to their newborn pups. Environ. Res. 2006, 101, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Pinyayev, T.S.; Kohan, M.J.; Herbin-Davis, K.; Creed, J.T.; Thomas, D.J. Preabsorptive metabolism of sodium arsenate by anaerobic microbiota of mouse cecum forms a variety of methylated and thiolated arsenicals. Chem. Res. Toxicol. 2011, 24, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.S.; Hoy, K.S.; Schofield, J.R.; Uppal, J.S.; Lin, Y.; Lu, X.; Peng, H.; Le, X.C. Arsenic speciation analysis: A Rev.iew with an emphasis on chromatographic separations. TrAC Trends Anal. Chem. 2020, 123, 115770. [Google Scholar] [CrossRef]
- Cubadda, F.; Jackson, B.P.; Cottingham, K.L.; Van Horne, Y.O.; Kurzius-Spencer, M. Human exposure to dietary inorganic arsenic and other arsenic species: State of knowledge, gaps and uncertainties. Sci. Total Environ. 2017, 579, 1228–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlin, D.J.; Naujokas, M.F.; Bradham, K.D.; Cowden, J.; Heacock, M.; Henry, H.F.; Lee, J.S.; Thomas, D.J.; Thompson, C.; Tokar, E.J.; et al. Arsenic and Environmental Health: State of the Science and Future Research Opportunities. Environ. Health Perspect. 2016, 124, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Kalman, D.A.; Dills, R.L.; Steinmaus, C.; Yunus, M.; Khan, A.F.; Prodhan, M.M.; Yuan, Y.; Smith, A.H. Occurrence of trivalent monomethyl arsenic and other urinary arsenic species in a highly exposed juvenile population in Bangladesh. J. Expo. Sci. Environ. Epidemiol. 2014, 24, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Hirano, S. Metabolism of arsenic and its toxicological relevance. Arch. Toxicol. 2013, 87, 969–979. [Google Scholar] [CrossRef]
- Wang, T.C.; Jan, K.Y.; Wang, A.S.; Gurr, J.R. Trivalent arsenicals induce lipid peroxidation, protein carbonylation, and oxidative DNA damage in human urothelial cells. Mutat. Res. 2007, 615, 75–86. [Google Scholar] [CrossRef]
- Rahman, M.S.; Hughes, M.F. In vitro percutaneous absorption of monosodium methanearsonate and disodium methanearsonate in female B6C3F1 mice. J. Toxicol. Environ. Health 1994, 41, 421–433. [Google Scholar] [CrossRef]
- Nagar, R.; Sarkar, D.; Makris, K.C.; Datta, R.; Sylvia, V.L. Bioavailability and bioaccessibility of arsenic in a soil amended with drinking-water treatment residuals. Arch. Environ. Contam. Toxicol. 2009, 57, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.F. Arsenic toxicity and potential mechanisms of action. Toxicol. Lett. 2002, 133, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhao, P.; Guo, G.; Hu, Z.; Tian, L.; Zhang, K.; Zhang, W.; Xing, M. The Role of Oxidative Stress in Gastrointestinal Tract Tissues Induced by Arsenic Toxicity in Cocks. Biol. Trace Elem. Res. 2015, 168, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, B.R.; Wu, J.; Li, H.S.; Jiang, Z.H.; Zhou, X.M.; Xu, C.H.; Ding, N.; Zha, J.-M.; He, W.Q. In vitro and In Vivo Approaches to Determine Intestinal Epithelial Cell Permeability. J. Vis. Exp. 2018, 140, 57032. [Google Scholar] [CrossRef] [Green Version]
- Twaddle, N.C.; Vanlandingham, M.; Fisher, J.W.; Doerge, D.R. Metabolism and disposition of arsenic species from controlled dosing with sodium arsenite in adult female CD-1 mice. III. Toxicokinetic studies following oral and intravenous administration. Food Chem. Toxicol. 2018, 121, 676–686. [Google Scholar] [CrossRef]
- Williams, K.M.; Gokulan, K.; Cerniglia, C.E.; Khare, S. Size and dose dependent effects of silver nanoparticle exposure on intestinal permeability in an in vitro model of the human gut epithelium. J. Nanobiotechnol. 2016, 14, 62. [Google Scholar] [CrossRef] [Green Version]
- Shaji, E.; Santosh, M.; Sarath, K.V.; Prakash, P.; Deepchand, V.; Divya, B. Arsenic contamination of groundwater: A global synopsis with focus on the Indian Peninsula. Geosci. Front. 2021, 12, 101079. [Google Scholar] [CrossRef]
- Tsuji, J.S.; Chang, E.T.; Gentry, P.R.; Clewell, H.J.; Boffetta, P.; Cohen, S.M. Dose-response for assessing the cancer risk of inorganic arsenic in drinking water: The scientific basis for use of a threshold approach. Crit. Rev. Toxicol. 2019, 49, 36–84. [Google Scholar] [CrossRef]
- Holmgren, G.; Synnergren, J.; Bogestal, Y.; Ameen, C.; Akesson, K.; Holmgren, S.; Lindahl, A.; Sartipy, P. Identification of novel biomarkers for doxorubicin-induced toxicity in human cardiomyocytes derived from pluripotent stem cells. Toxicology 2015, 328, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Balda, M.S.; Matter, K. Epithelial cell adhesion and the regulation of gene expression. Trends Cell Biol. 2003, 13, 310–318. [Google Scholar] [CrossRef]
- Smith, S.M.; Wunder, M.B.; Norris, D.A.; Shellman, Y.G. A simple protocol for using a LDH-based cytotoxicity assay to assess the effects of death and growth inhibition at the same time. PLoS ONE 2011, 6, e26908. [Google Scholar] [CrossRef]
- Moe, B.; Peng, H.; Lu, X.; Chen, B.; Chen, L.W.L.; Gabos, S.; Li, X.-F.; Le, X.C. Comparative cytotoxicity of fourteen trivalent and pentavalent arsenic species determined using real-time cell sensing. J. Environ. Sci. 2016, 49, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Yamaura, Y.; Chapron, B.D.; Wang, Z.; Himmelfarb, J.; Thummel, K.E. Functional Comparison of Human Colonic Carcinoma Cell Lines and Primary Small Intestinal Epithelial Cells for Investigations of Intestinal Drug Permeability and First-Pass Metabolism. Drug Metab. Dispos. 2016, 44, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donato, R.P.; El-Merhibi, A.; Gundsambuu, B.; Mak, K.Y.; Formosa, E.R.; Wang, X.; Abbott, C.A.; Powell, B.C. Studying permeability in a commonly used epithelial cell line: T84 intestinal epithelial cells. Methods Mol. Biol. 2011, 763, 115–137. [Google Scholar] [CrossRef]
- Devriese, S.; Van den Bossche, L.; Van Welden, S.; Holvoet, T.; Pinheiro, I.; Hindryckx, P.; De Vos, M.; Laukens, D. T84 monolayers are superior to Caco-2 as a model system of colonocytes. Histochem. Cell Biol. 2017, 148, 85–93. [Google Scholar] [CrossRef]
- Welcome, M.O. Intercellular network of junctions of the gastrointestinal tract. In Gastrointestinal Physiology; Springer: Cham, Switzerland, 2018; pp. 201–225. [Google Scholar]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef]
- Matter, K.; Balda, M.S. Functional analysis of tight junctions. Methods 2003, 30, 228–234. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukita, S.; Tanaka, H.; Tamura, A. The Claudins: From Tight Junctions to Biological Systems. Trends Biochem. Sci. 2019, 44, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.M.; Myers, L.S.; Kurundkar, A.R.; Maheshwari, A.; Nusrat, A.; Lin, P.W. Probiotic bacteria induce maturation of intestinal claudin 3 expression and barrier function. Am. J. Pathol. 2012, 180, 626–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajjari, M.; Behmanesh, M.; Sadeghizadeh, M.; Zeinoddini, M. Junctional adhesion molecules 2 and 3 may potentially be involved in progression of gastric adenocarcinoma tumors. Med. Oncol. 2013, 30, 380. [Google Scholar] [CrossRef]
- Jones, R.G.; Li, X.; Gray, P.D.; Kuang, J.; Clayton, F.; Samowitz, W.S.; Madison, B.B.; Gumucio, D.L.; Kuwada, S.K. Conditional deletion of beta1 integrins in the intestinal epithelium causes a loss of Hedgehog expression, intestinal hyperplasia, and early postnatal lethality. J. Cell Biol. 2006, 175, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.; Huang, K.; Gao, C.; Lau, Q.C.; Pan, H.; Xie, K.; Li, J.; Liu, R.; Zhang, T.; Xie, N.; et al. Proteomics identification of ITGB3 as a key regulator in reactive oxygen species-induced migration and invasion of colorectal cancer cells. Mol. Cell. Proteom. 2011, 10, M110.005397. [Google Scholar] [CrossRef] [Green Version]
- Goodenough, D.A.; Paul, D.L. Gap junctions. Cold Spring Harb. Perspect. Biol. 2009, 1, a002576. [Google Scholar] [CrossRef]
- Daniel, E.E.; Wang, Y.F. Gap junctions in intestinal smooth muscle and interstitial cells of Cajal. Microsc. Res. Tech. 1999, 47, 309–320. [Google Scholar] [CrossRef]
- King, T.J.; Gurley, K.E.; Prunty, J.; Shin, J.L.; Kemp, C.J.; Lampe, P.D. Deficiency in the gap junction protein connexin32 alters p27Kip1 tumor suppression and MAPK activation in a tissue-specific manner. Oncogene 2005, 24, 1718–1726. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, E.; Shao, Q.; Wang, H.L.; Langlois, S.; Laird, D.W. Connexins act as tumor suppressors in three-dimensional mammary cell organoids by regulating differentiation and angiogenesis. Cancer Res. 2006, 66, 9886–9894. [Google Scholar] [CrossRef] [Green Version]
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim. Biophys. Acta 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kuang, Y.; Wang, Y.; Xu, Q.; Ren, Q. Notch-4 silencing inhibits prostate cancer growth and EMT via the NF-kappaB pathway. Apoptosis 2017, 22, 877–884. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Focal Adhesions | Focal Adhesions | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Apical 12.8 | Apical 6.4 | Apical 3.2 | Apical 1.6 | Apical EGTA | Basal 12.8 | Basal 6.4 | Basal 3.2 | Basal 1.6 | Basal EGTA | ||
| CAV1 | −5.16 | −1.99 | 1.09 | −1.55 | −1.33 | CAV1 | −3.77 | −1.53 | −1.07 | 1.17 | −1.35 |
| CAV2 | −1.86 | −1.16 | −5.81 | −3.52 | −1.37 | CAV2 | 1.71 | 1.57 | 2.73 | 2.91 | 1.41 |
| ITGA1 | −5.09 | −1.22 | −1.60 | 1.28 | 3.41 | ITGA1 | −4.50 | −7.54 | 1.75 | −3.28 | 1.69 |
| ITGB2 | −1.37 | 1.09 | 1.98 | 1.39 | 5.36 | ITGB2 | −5.42 | −5.84 | −3.59 | −1.85 | 1.02 |
| ITGB3 | 6.89 | 5.43 | 1.40 | 1.01 | −2.83 | ITGB3 | 36.90 | 9.43 | 6.20 | 11.40 | 1.24 |
| Tight Junctions | Tight Junctions | ||||||||||
| Apical 12.8 | Apical 6.4 | Apical 3.2 | Apical 1.6 | Apical EGTA | Basal 12.8 | Basal 6.4 | Basal 3.2 | Basal 1.6 | Basal EGTA | ||
| CLDN1 | 7.12 | 7.11 | 2.56 | 2.96 | 1.58 | CLDN1 | 5.17 | 4.37 | 5.22 | 3.74 | 1.11 |
| CLDN15 | 3.96 | 4.00 | 8.61 | 3.32 | 5.88 | CLDN15 | 3.54 | 2.30 | 1.60 | 1.38 | 1.14 |
| CLDN2 | −3.29 | −3.44 | −1.74 | 1.01 | 2.06 | CLDN2 | −37.65 | −2.86 | −1.69 | −1.03 | −1.12 |
| CLDN3 | −5.46 | −8.63 | −19.88 | −2.31 | 1.39 | CLDN3 | −26.09 | −16.75 | −36.27 | −7.14 | 1.25 |
| CLDN4 | −1.82 | −2.22 | −1.08 | −1.17 | 1.44 | CLDN4 | −7.12 | −2.31 | −2.48 | −3.40 | −1.61 |
| CLDN6 | 2.51 | 1.10 | −6.53 | 1.15 | 1.37 | CLDN6 | 7.78 | 1.05 | 1.87 | 2.03 | 1.00 |
| CLDN7 | −1.54 | −1.74 | −1.08 | 1.04 | 1.62 | CLDN7 | −4.25 | −1.81 | −1.48 | −2.25 | −1.34 |
| ESAM | −7.28 | −4.11 | −6.34 | −3.08 | 1.02 | ESAM | −2.66 | −5.22 | −2.84 | −5.07 | −1.05 |
| F11R | 1.26 | −1.99 | 1.41 | 1.57 | 1.84 | F11R | 2.71 | −1.03 | −1.24 | −1.41 | 1.07 |
| ICAM1 | 2.01 | 2.85 | 5.14 | 1.78 | 2.32 | ICAM1 | −2.90 | 1.49 | 1.81 | 2.12 | −1.64 |
| JAM2 | 3.24 | 2.58 | −1.14 | −1.12 | 1.14 | JAM2 | 14.71 | 10.71 | 13.35 | 9.49 | 3.39 |
| OCLN | −1.00 | −3.37 | −3.05 | −1.21 | 1.40 | OCLN | 1.14 | −1.08 | −1.71 | −1.72 | 1.33 |
| TJP2 | 1.49 | 1.10 | −1.05 | 1.53 | 2.50 | TJP2 | −1.54 | −1.07 | 1.12 | −1.05 | 1.44 |
| TJP3 | −6.95 | −3.89 | −1.58 | −2.53 | −1.08 | TJP3 | −8.48 | −3.61 | −2.67 | −2.41 | −1.35 |
| Gap Junctions | Gap Junctions | ||||||||||
| Apical 12.8 | Apical 6.4 | Apical 3.2 | Apical 1.6 | Apical EGTA | Basal 12.8 | Basal 6.4 | Basal 3.2 | Basal 1.6 | Basal EGTA | ||
| GJB1 | −11.35 | −3.07 | −1.58 | −1.05 | 1.69 | GJB1 | −82.02 | −20.71 | −10.19 | −7.14 | −1.35 |
| GJB2 | −2.87 | −2.51 | −1.29 | −1.18 | −1.45 | GJB2 | −12.99 | −3.78 | −2.10 | −3.98 | −1.64 |
| Adherent Junctions | Adherent Junctions | ||||||||||
| Apical 12.8 | Apical 6.4 | Apical 3.2 | Apical 1.6 | Apical EGTA | Basal 12.8 | Basal 6.4 | Basal 3.2 | Basal 1.6 | Basal EGTA | ||
| NOTCH2 | −4.73 | −1.76 | −1.16 | −3.25 | −1.15 | NOTCH2 | −6.25 | 1.02 | 2.35 | 1.15 | −1.66 |
| NOTCH4 | 2.44 | 2.60 | 1.04 | 2.76 | 3.85 | NOTCH4 | 13.22 | 3.34 | 2.42 | 2.37 | 2.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parajuli, P.; Gokulan, K.; Khare, S. Preclinical In Vitro Model to Assess the Changes in Permeability and Cytotoxicity of Polarized Intestinal Epithelial Cells during Exposure Mimicking Oral or Intravenous Routes: An Example of Arsenite Exposure. Int. J. Mol. Sci. 2022, 23, 4851. https://doi.org/10.3390/ijms23094851
Parajuli P, Gokulan K, Khare S. Preclinical In Vitro Model to Assess the Changes in Permeability and Cytotoxicity of Polarized Intestinal Epithelial Cells during Exposure Mimicking Oral or Intravenous Routes: An Example of Arsenite Exposure. International Journal of Molecular Sciences. 2022; 23(9):4851. https://doi.org/10.3390/ijms23094851
Chicago/Turabian StyleParajuli, Pravin, Kuppan Gokulan, and Sangeeta Khare. 2022. "Preclinical In Vitro Model to Assess the Changes in Permeability and Cytotoxicity of Polarized Intestinal Epithelial Cells during Exposure Mimicking Oral or Intravenous Routes: An Example of Arsenite Exposure" International Journal of Molecular Sciences 23, no. 9: 4851. https://doi.org/10.3390/ijms23094851