
Molecular Cloning and Characterization of MbMYB108, a Malus baccata MYB Transcription Factor Gene, with Functions in Tolerance to Cold and Drought Stress in Transgenic Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
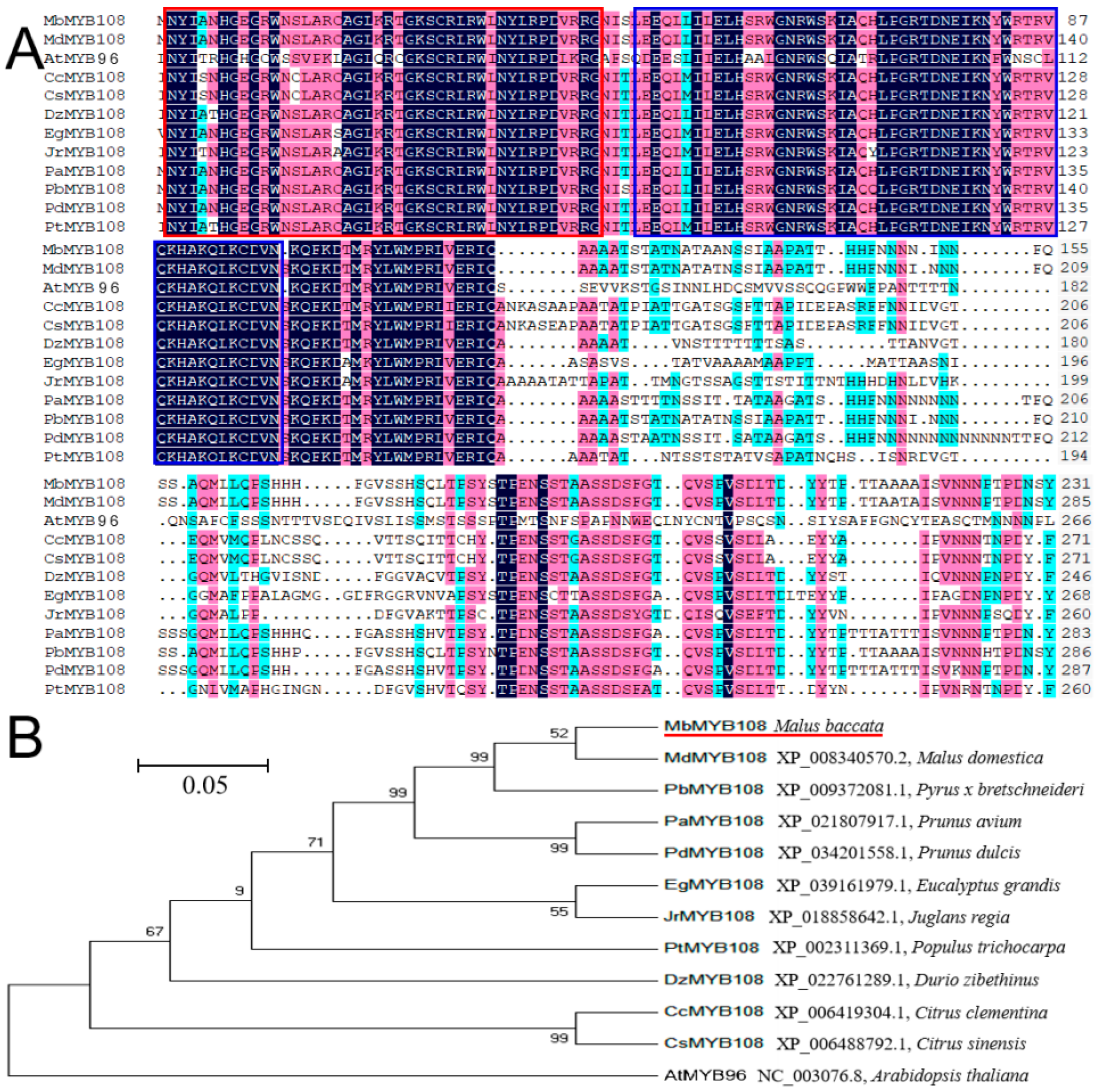
2.1. Isolation of MbMYB108 Gene from M. baccata
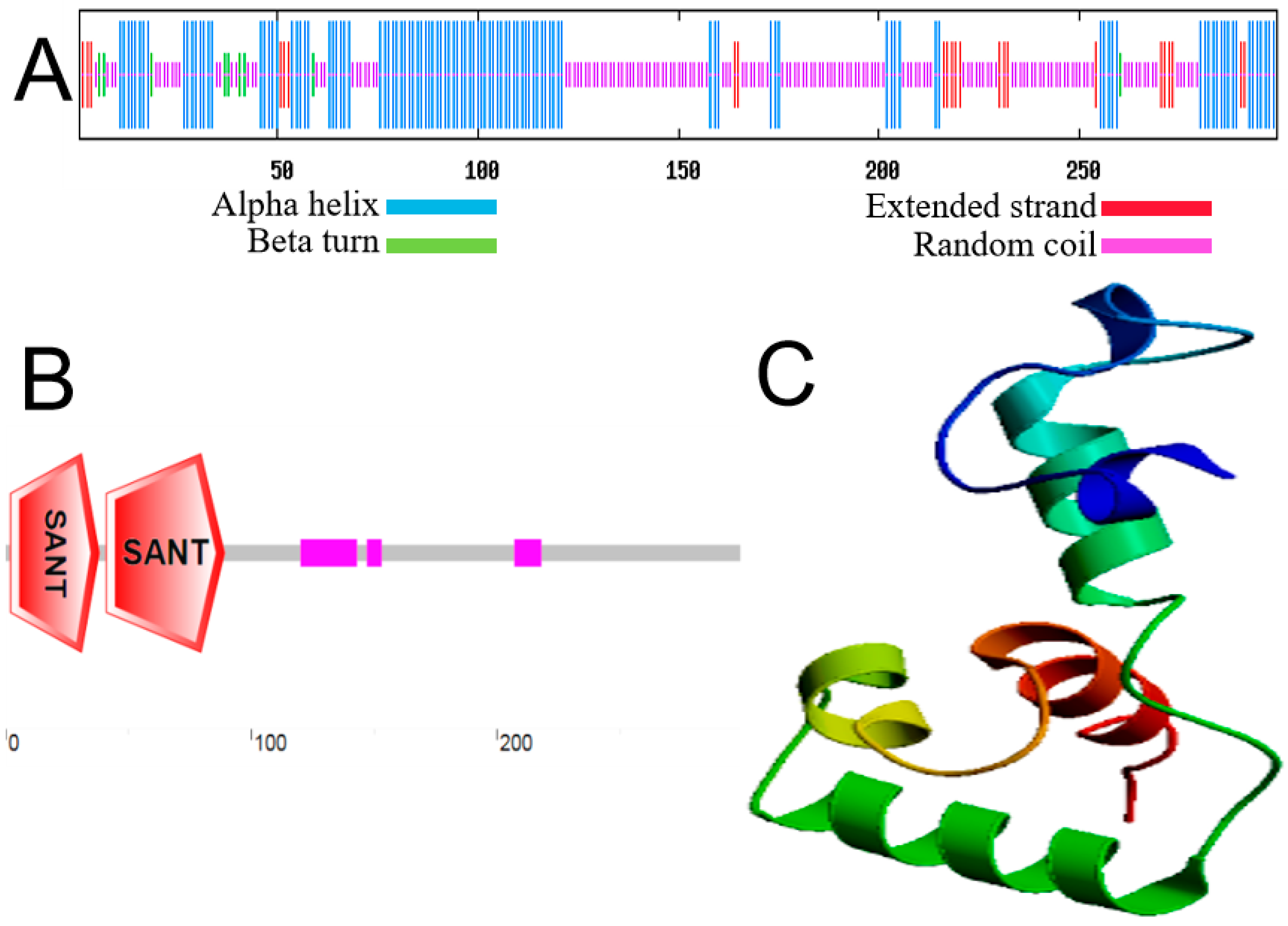
2.2. Phylogenetic Relationship and Structural Prediction of MbMYB108 Protein
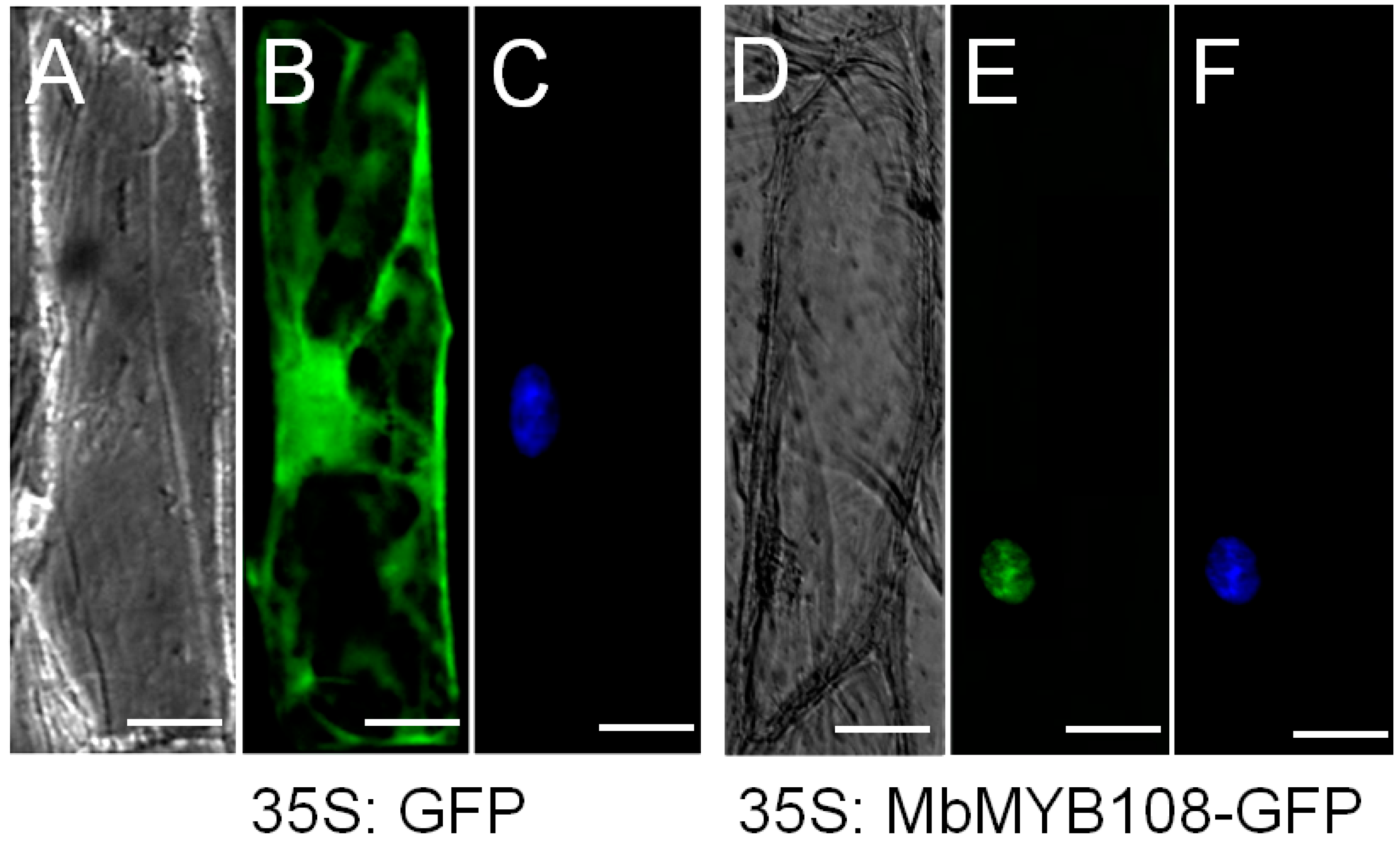
2.3. MbMYB108 Was Localized in the Nucleus
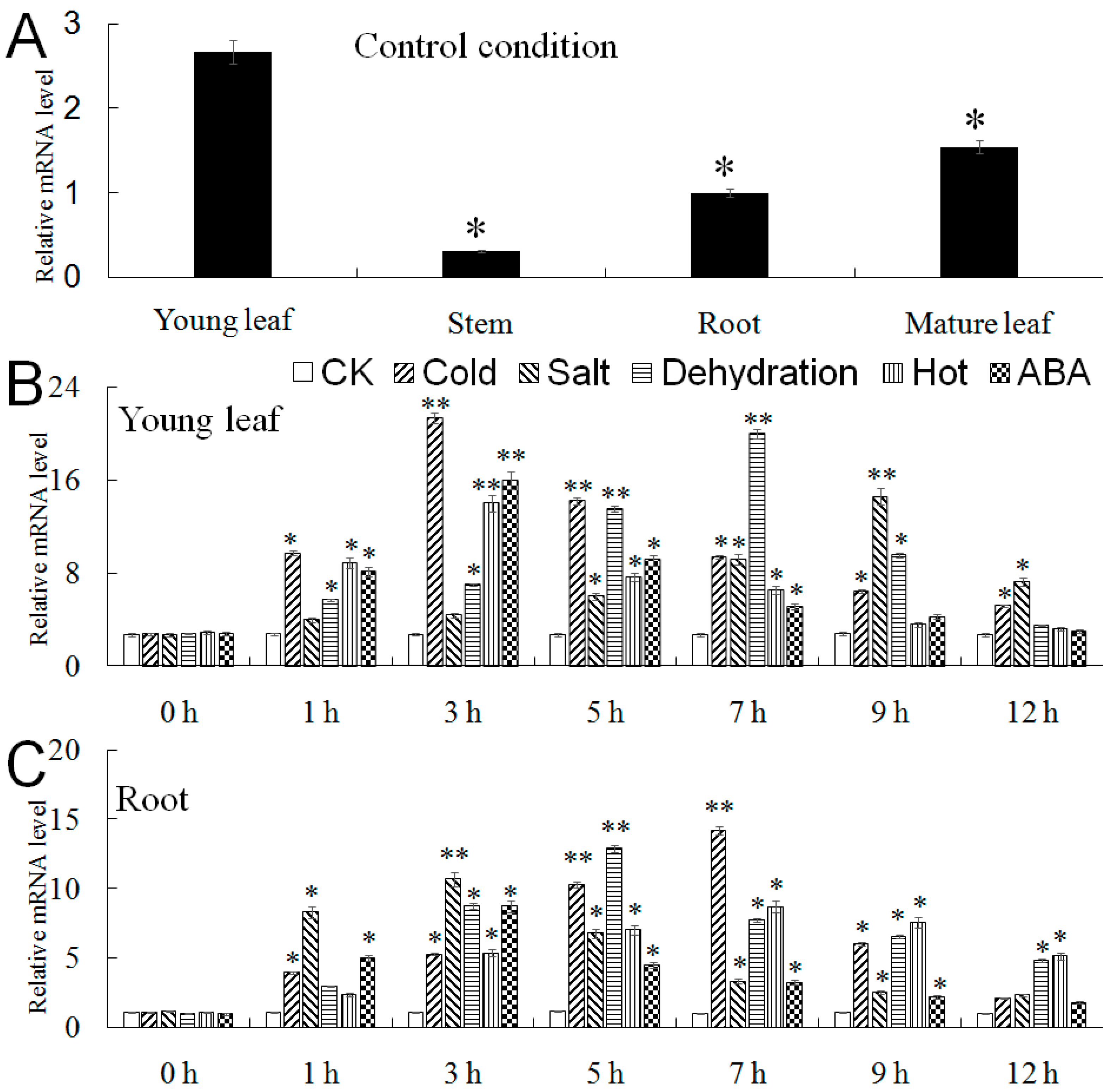
2.4. Expression Level of MbMYB108 in M. baccata
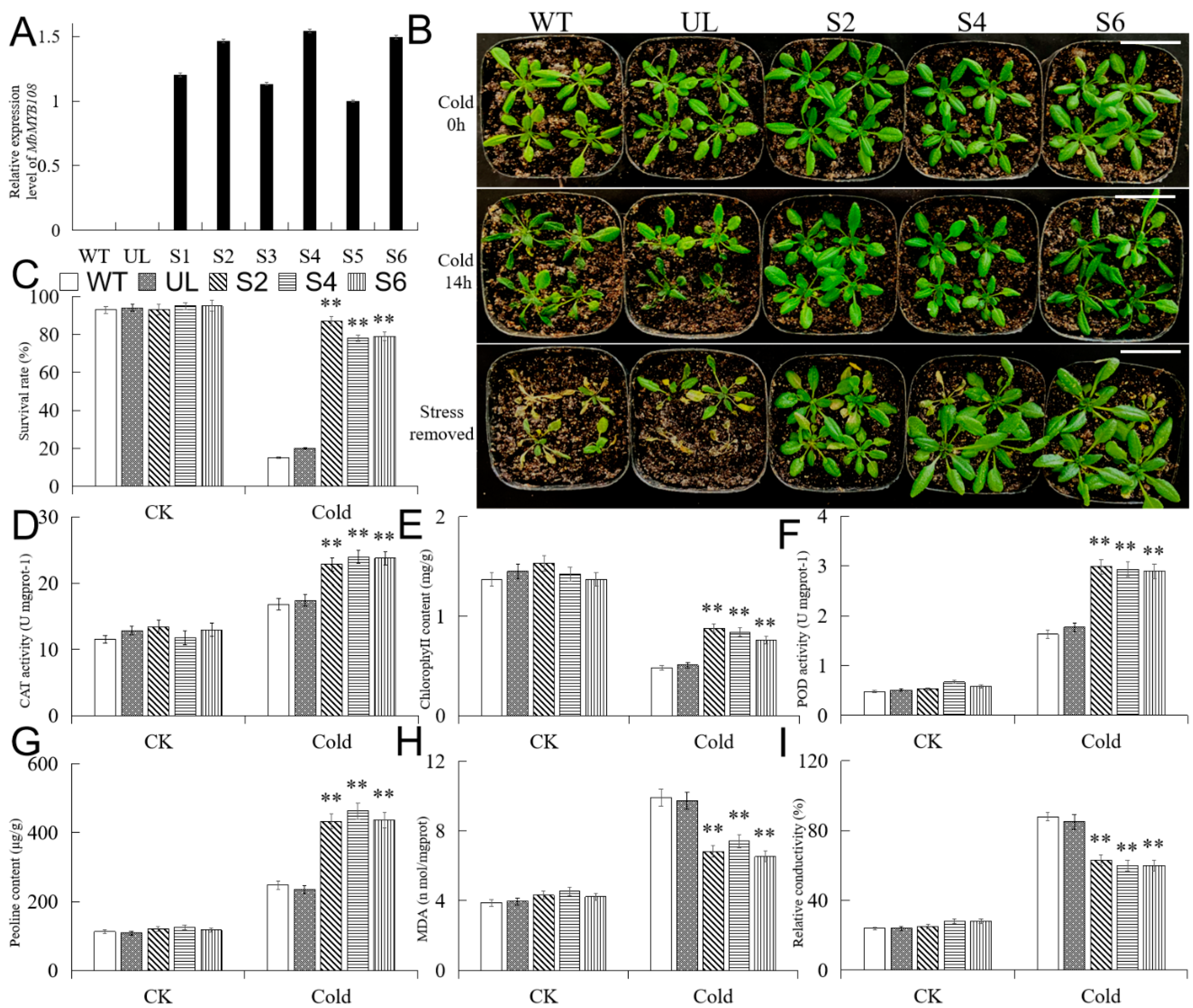
2.5. Overexpression of MbMYB108 Improves Cold Tolerance in Transgenic A. thaliana
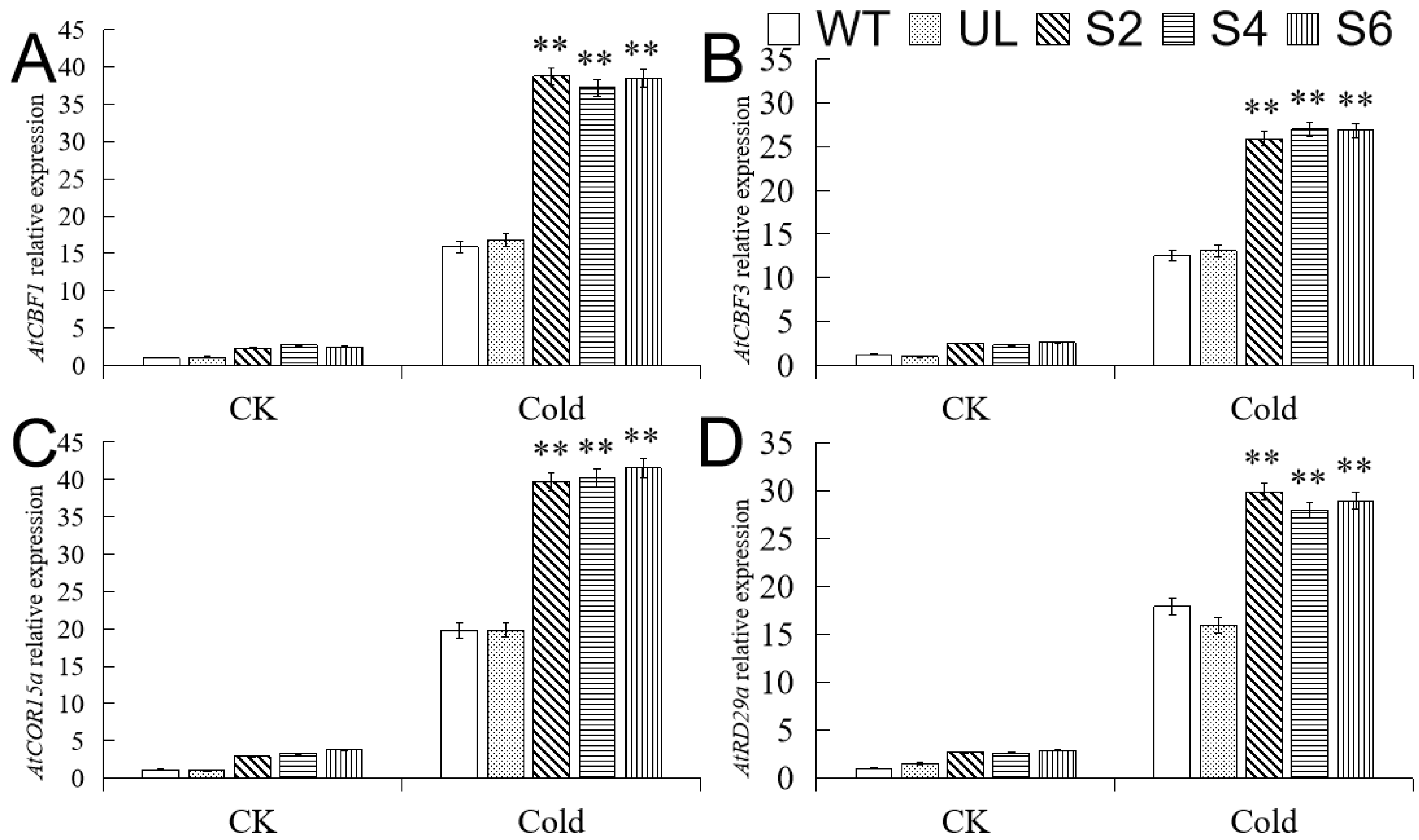
2.6. Overexpression of MbMYB108 Promotes the Expression of Cold Stress-Related Genes
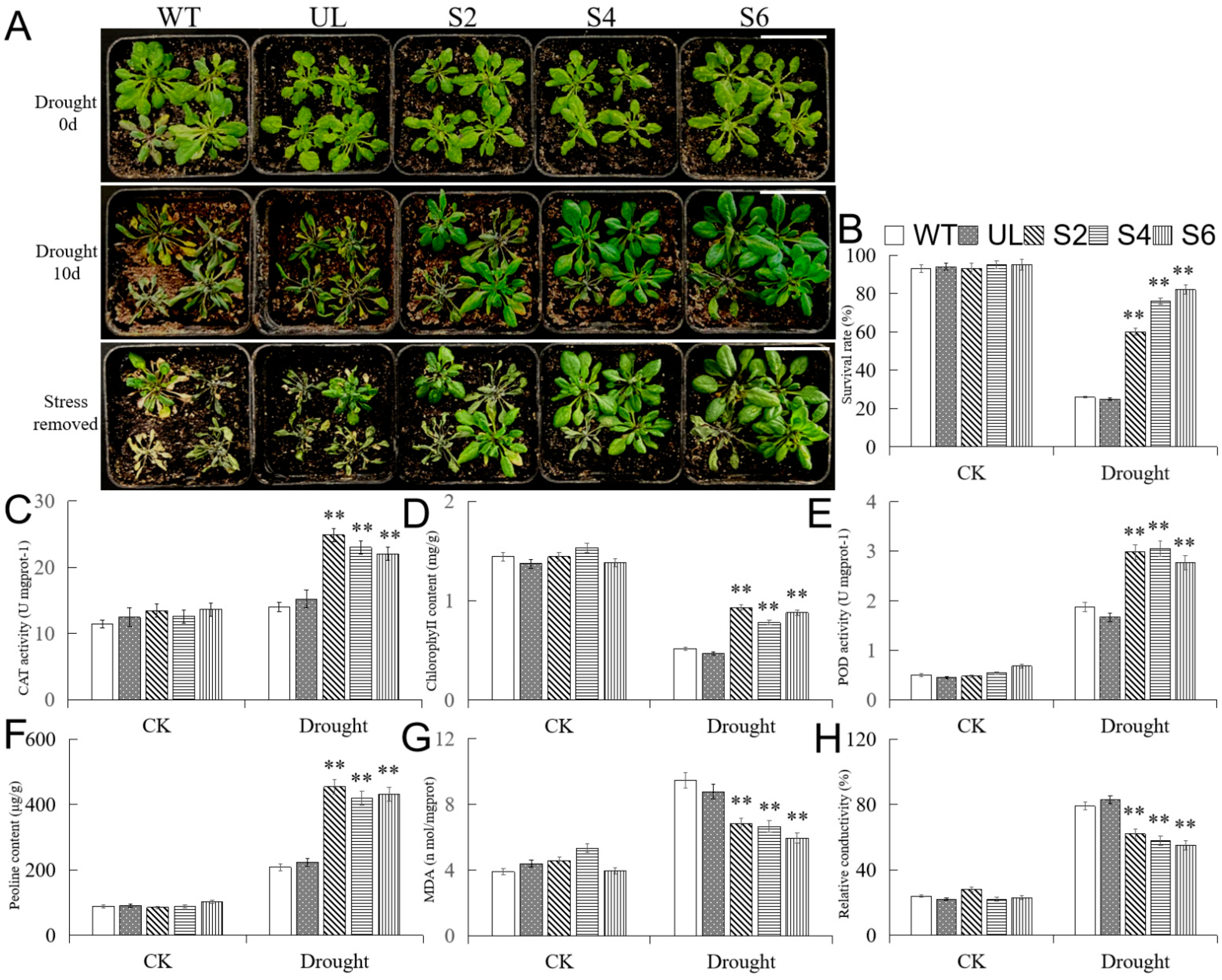
2.7. Overexpression of MbMYB108 Improves Drought Tolerance in Transgenic A. thaliana
2.8. Overexpression of MbMYB108 Promotes the Expression of Drought Stress-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Isolation and Cloning of MbMYB108
4.3. Sequence Analysis and Structure Prediction of MbMYB108
4.4. Subcellular Localization Analysis of MbMYB108 Protein
4.5. Quantitative Real-Time PCR (qPCR) Expression Analysis of MbMYB108
4.6. Generation of Transgenic A. thaliana Overexpressing MbMYB108
4.7. Stress Treatment and Determination of Related Physiological Indexes in A. thaliana
4.8. Expression Analysis of MbMYB108 Downstream Genes
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Feng, P.; Chen, P.; Hong, W.; Zhao, X.; Liu, X. Research Progress of MYB Transcription Factor Family in Arabidopsis thaliana. Life Sci. Res. 2016, 20, 555–560. [Google Scholar]
- Shi, M.Z.; Xie, D.Y. Biosynthesis and metabolic engineering of anthocyanins in Arabidopsis thaliana. Recent Pat. Biotechnol. 2014, 8, 47–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Haga, N.; Kato, K.; Murase, M.; Araki, S.; Kubo, M.; Demura, T.; Suzuki, K.; Muller, I.; Voss, U.; Jurgens, G. R1R2R3-Myb proteins positively regulate cytokinesis through activation of KNOLLE transcription in Arabidopsis thaliana. Development 2007, 134, 1101–1110. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Li, S.; Liu, S.; Hu, C.; Zhang, J.; Moriguchi, T. Genome-wide classification and evolutionary and expression analyses of citrus MYB transcription factor families in sweet orange. PLoS ONE 2014, 9, e112375. [Google Scholar] [CrossRef]
- Qiao, M.; Yu, Y.; Xiang, F. The roles of the Arabidopsis R2R3-MYB transcription factors in the stress responses. Chin. Bull. Life Sci. 2009, 21, 145–150. [Google Scholar]
- Shinozaki, Y.K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stressresponsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB Gene, OsMYB3R-2, Increases Tolerance to Freezing, Drought, and Salt Stress in Transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [Green Version]
- Casaretto, J.A.; El-Kereamy, A.; Zeng, B.; Stiegelmeyer, S.; Chen, X.; Bi, Y.; Rothstein, S. Expression of OsMYB55 in maize activates stress-responsive genes and enhances heat and drought tolerance. BMC Genom. 2016, 17, 312. [Google Scholar] [CrossRef] [Green Version]
- Vannini, C.; Locatelli, F.; Bracale, M.; Magnani, E.; Marsoni, M.; Osnato, M.; Mattana, M.; Baldoni, E.; Coraggio, I. Overexpression of the rice Osmyb4 gene increases chilling and freezing tolerance of Arabidopsis thaliana plants. Plant J. 2004, 37, 115–127. [Google Scholar] [CrossRef]
- Sun, X.; Liu, J.; Yuan, X.; Pan, X.; Du, W.; Ren, H.; Ma, Y.; ABE, J.; Qiu, L.; Liu, B. Cloning and expression analysis of GmMYB gene induced by abiotic stress. J. Crops 2012, 38, 360–368. [Google Scholar]
- Chen, X.; Hao, Y.; Yang, H.; Mao, Z.; Jiang, Y.; Peng, F.; Shen, X.; Zhang, J.; Chen, X.; Shu, H. 10 key technologies for high-quality and efficient development of my country’s apple industry. Chin. Fruit Tree 2010, 4, 65–67. [Google Scholar]
- Yue, Z.; Liu, H.; Ma, F. The Malus carotenoid cleavage dioxygenase 7 is involved in stress response and regulated by basic pentacysteine 1. Sci. Hortic. 2015, 192, 264–270. [Google Scholar] [CrossRef]
- Ding, N.; Chen, J.; Feng, Y.; Sha, J.; Zhang, M.; Jiang, Y. Effects of dwarf apple loading on nitrogen absorption, distribution and utilization. J. Hortic. 2016, 43, 549–556. [Google Scholar]
- Liu, J.; She, L.; Lan, X.; Lu, Y.; Yang, J.; Liu, H.; Lu, C.; Chen, Y. Bioinformatics and expression analysis of the MYB family of Himalayan Mirabilis. Chin. Agric. Sci. Bull. 2020, 36, 54–61. [Google Scholar]
- Xing, Z. Cloning and Functional Analysis of Potato Gene StR2R3-MYB1. Master’s Thesis, Hunan Agricultural University, Haikou, China, 2014; p. 2. [Google Scholar]
- Ren, L.; Zhu, B.; Zhang, Y.; Wang, H.; Li, C.; Su, Y.; Ba, C. The research of applying primer premier 5.0 to design PCR primer. J. Jinzhou Med. Coll. 2004, 25, 43–46. [Google Scholar]
- Boyer, L.; Latek, R.; Peterson, C. The SANT domain: A unique histone-tail-binding module? Nat. Rev. Mol. Cell Biol. 2004, 5, 158–163. [Google Scholar] [CrossRef]
- Lv, B.; Zhang, W.; Li, C.; Ming, F. MYB family transcription factor OsMYB84 participates in salt stress response through ABA signaling pathway. Fudan J. 2015, 54, 591–600. [Google Scholar]
- Xie, Y.P. Mechanism of Apple MdMYB88 and MdMYB124 Transcription Factors in Low Temperature and Drought Stress. Ph.D. Thesis, Northwest A&F University, Xianyang, China, 2018; p. 23. [Google Scholar]
- Xu, C.X. Advances in mechanisms of improving plant cold resistance. J. Ecol. 2012, 32, 7966–7980. [Google Scholar]
- Ke, X.; He, L.; Su, Z. Relative chlorophyll index and distribution of four woody plants in southern China. J. Cent. South Univ. For. Technol. 2010, 30, 82–86. [Google Scholar]
- Pan, R.C.; Wang, X.J.; Li, N.H. Plant Physiology Version 7; High Education Press: Beijing, China, 2012; pp. 438–455. [Google Scholar]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Yao, H. Physiological and Biochemical Study on Cold Resistance of Four Cupressaceae Plants. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2012; p. 2. [Google Scholar]
- Jiao, R.; Liu, H.; Liu, G.; Wang, S.; Hou, N.; Wang, Q.; Liu, Z.; Feng, X.; Hu, X.; Jin, Y. On Proline Accumulation and Plant Resistance to Osmotic Stress. Chin. Agric. Sci. Bull. 2011, 27, 216–221. [Google Scholar]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Penconazole and calcium ameliorate drought stress in canola by upregulating the antioxidative enzymes. Funct. Plant Biol. 2020, 47, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, X.Y. Changes of intracellular Ca2+ levels in plant cells under stress. Anhui Agric. Sci. Bull. 2010, 16, 29–31. [Google Scholar]
- Zhang, J.; Meng, Y.; Song, Y.; Liu, S.; Zhuang, W. Research Developments of Ca2+-CaM Signal System and Its Regulation in Plant. J. Chongqing Norm. Univ. 2005, 22, 49–52. [Google Scholar]
- Liu, G.S.; Chen, J. The role of calcium-dependent protein kinases (CDPKs) in plant calcium signal transduction. Bull. Phys. 2003, 20, 160–167. [Google Scholar]
- Verma, P.; Sanyal, S.K.; Pandey, G.K. Ca2+-CBL-CIPK: A modulator system for efficient nutrient acquisition. Plant Cell Rep. 2021, 40, 2111–2122. [Google Scholar] [CrossRef]
- Li, M.; Lin, L.; Zhang, Y.; Sui, N. ZmMYB31, a R2R3-MYB transcription factor in maize’ positively regulates expression of CBF genes and enhances resistance to chilling and oxidative stress. Mol. Biol. Rep. 2019, 46, 3937–3944. [Google Scholar] [CrossRef]
- Yang, A.; Dai, X.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- Zhou, M.Q.; Shen, C.; Wu, L.H.; Tang, K.X.; Lin, J. CBF-dependent signaling pathway: A key responder to low temperature stress in plants. Crit. Rev. Biotechnol. 2011, 31, 186–192. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Thomashow, M.F. So what’s new in the field of plant cold acclimation? Lots! Plant Physiol. 2001, 125, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, S.S.; Wilhelm, K.S.; Thomashow, M.F. The 5-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought- and ABA-regulated gene expression. Plant Mol. Biol. 1994, 24, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cisacting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.H.; Hong, X.; Agarwal, M.; Zhu, J.K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, M.; Hao, Y.; Kapoor, A.; Dong, C.H.; Fujii, H.; Zheng, X.; Zhu, J.K. A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance. J. Biol. Chem. 2006, 281, 37636–37645. [Google Scholar] [CrossRef] [Green Version]
- Doherty, C.J.; Van-Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for Arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. Plant Cell 2009, 21, 972–984. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, 164376. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells 2020, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Veljovic-Jovanovic, S.; Driscoll, S.; Novitskaya, L.; Foyer, C.H. Drought and oxidative load in the leaves of C3plants: A predominant role for photorespiration? Ann. Bot. 2002, 89, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, Y.; Wang, L.; Hu, P.; Wang, Y.; Jia, Y.; Zhang, C.; Zhang, Y.; Zhang, Y.; Wang, C.; et al. Expression of the MYB transcription factor gene BplMYB46 affects abiotic stress tolerance and secondary cell wall deposition in Betula platyphylla. Plant Biotechnol. J. 2016, 15, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhu, H.; He, S.; Zhai, H.; Liu, Q. A Novel Sweet potato Transcription Factor Gene IbMYB116 Enhances Drought Tolerance in Transgenic Arabidopsis. Front. Plant Sci. 2019, 10, 1025. [Google Scholar] [CrossRef] [Green Version]
- Seo, P.; Xiang, F.; Qiao, M.; Park, J.; Lee, Y.; Kim, S.; Lee, Y.; Park, W.; Park, C. The MYB96 Transcription Factor Mediates Abscisic Acid Signaling during Drought Stress Response in Arabidopsis. Plant Physiol. 2009, 151, 275–289. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N.; Sopory, S.K. Plant signaling in stress: G-protein coupled receptors, heterotrimeric G-proteins and signal coupling via phospholipases. Plant Signal. Behav. 2008, 3, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Gavassi, M.A.; Silva, G.S.; da Silva, C.D.M.S.; Thompson, A.J.; Macleod, K.; Oliveira, P.M.R.; Cavalheiro, M.F.; Domingues, D.S.; Habermann, G. NCED expression is related to increased ABA biosynthesis and stomatal closure under aluminum stress. Environ. Exp. Bot. 2021, 185, 104404. [Google Scholar] [CrossRef]
- Taylor, I.B.; Burbidge, A.; Thompson, A.J. Control of abscisic acid synthesis. J. Exp. Bot. 2000, 51, 1563–1574. [Google Scholar] [CrossRef]
- Yang, Y.; Tan, B. A Distal ABA Responsive Element in AtNCED3 Promoter Is Required for Positive Feedback Regulation of ABA Biosynthesis in Arabidopsis. PLoS ONE 2014, 9, e87283. [Google Scholar]
- Torres, G.A.; Lelandais-Brière, C.; Besin, E.; Jubier, M.F.; Roche, O.; Mazubert, C.; Corre-Menguy, F.; Hartmann, C. Characterization of the expression of Phaseolus vulgaris OCT1, a dehydration-regulated gene that encodes a new type of phloem transporter. Plant Mol. Biol. 2003, 51, 341–349. [Google Scholar] [CrossRef]
- Ren, X.; Chen, Z.; Liu, Y.; Zhang, H.; Zhang, M.; Liu, Q.; Hong, X.; Zhu, J.K.; Gong, Z. ABO3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in Arabidopsis. Plant J. 2010, 63, 417–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.; Merlot, S.; Giraudat, J. The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encode homologous protein phosphatases 2C involved in abscisic acid signal transduction. Plant Cell 1997, 9, 759–771. [Google Scholar] [PubMed] [Green Version]
- Gosti, F.; Beaudoin, N.; Serizet, C.; Webb, A.A.; Vartanian, N.; Giraudat, J. ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. Plant Cell 1999, 11, 1897–1910. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Fujita, Y.; Kanamori, N.; Katagiri, T.; Umezawa, T.; Kidokoro, S.; Maruyama, K.; Yoshida, T.; Ishiyama, K.; Kobayashi, M.; et al. Three arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant Cell Physiol. 2009, 50, 1345–1363. [Google Scholar] [CrossRef] [Green Version]
- Muschietti, J.; McCormick, S. Abscisic acid (ABA) receptors: Light at the end of the tunnel. F1000 Biol. Rep. 2010, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Han, J.; Li, L.; Zhang, Q.; Yang, G.; He, G. Wheat PP2C-a10 regulates seed germination and drought tolerance in transgenic Arabidopsis. Plant Cell Rep. 2020, 39, 635–651. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Wang, Y.; Zhang, L.; Ma, L.; Zhang, X.; Xu, X.; Han, Z. Isolation and functional characterization of MxCS1: A gene encoding a citrate synthase in Malus xiaojinensis. Biol. Plant. 2012, 56, 50–56. [Google Scholar] [CrossRef]
- Hoagland, D.R. Optimum nutrient solutions for plants. Science 1920, 1354, 562–564. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Zhang, Z.; Ding, H.; Chai, L.; Liu, W.; Li, H.; Yang, G. Isolation and characterization of MbWRKY2 gene involved in enhanced drought tolerance in transgenic tobacco. J. Plant Interact. 2018, 13, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Ding, H.; Chai, L.; Liu, W.; Zhang, Z.; Hou, Y.; Yang, G. Isolation and characterization of MbWRKY1, a WRKY transcription factor gene from Malus baccata (L.) Borkh involved in drought tolerance. Can. J. Plant Sci. 2018, 98, 1023–1034. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Li, J.; Liu, W.; Yu, Z.; Shi, Y.; Lv, B.; Wang, B.; Han, D. Molecular cloning and characterization of MxNAS2, a gene encoding nicotianamine synthase in Malus xiaojinensis, with functions in tolerance to iron stress and misshapen flower in transgenic tobacco. Sci. Hortic. 2015, 183, 77–86. [Google Scholar] [CrossRef]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Peirson, S.; Butler, J.; Foster, R. Experimental validation of novel and conventional approaches to quantitative real-time PCR data analysis. Nucleic. Acids. Res. 2003, 31, e73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Han, D.; Han, J.; Xu, T.; Li, X.; Yao, C.; Li, T.; Sun, X.; Wang, X.; Yang, G. Overexpression of MbERF12, an ERF gene from Malus baccata (L.) Borkh increases cold and salt tolerance in Arabidopsis thaliana associated with the ROS scavenging through ethylene signal transduction. Vitr. Cell. Dev. Biol.-Plant 2021, 57, 760–770. [Google Scholar] [CrossRef]
- Han, D.; Wang, Y.; Zhang, Z.; Pu, Q.; Ding, H.; Han, J.; Fan, T.; Bai, X.; Yang, G. Isolation and functional analysis of MxCS3: A gene encoding a citrate synthase in Malus xiaojinensis, with functions in tolerance to iron stress and abnormal flower in transgenic Arabidopsis thaliana. Plant Growth Regul. 2017, 82, 479–489. [Google Scholar] [CrossRef]
- Xu, Y.T.; Xu, Q. Transformation of Arabidopsis thaliana by floral dip. Bio-101 2018, e1010203. [Google Scholar] [CrossRef]
- Ren, L.; Liu, H.; Wang, H.; Zhu, Z. Construction of Expression Vectors of Brassica chinensis Myb28 Gene and Transformation to Arabidopsis thaliana. Biotechnol. Bull. 2012, 3, 80–84. [Google Scholar]
- Yao, C.; Li, X.; Li, Y.; Yang, G.; Liu, W.; Shao, B.; Zhong, J.; Huang, P.; Han, D. Overexpression of a Malus baccata MYB Transcription FactorGene MbMYB4 Increases Cold and Drought Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 1794. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, S.; Liu, Y.; Xu, J.; Liu, T.; Dong, S. Effectiveness of lysozyme coatings and 1-MCP treatments on storage and preservation of kiwifruit. Food Chem. 2019, 288, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Qian, X.; Jiang, T.; Zheng, X. Effect of eugenol fumigation treatment on chilling injury and CBF gene expression in eggplant fruit during cold storage. Food Chem. 2019, 292, 143–150. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Li, P.; Zheng, X.; Liu, Y.; Zhu, Y. Pre-storage application of oxalic acid alleviates chilling injury in mango fruit by modulating proline metabolism and energy status under chilling stress. Food Chem. 2014, 142, 72–78. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Sharma, A.; Yuan, H.; Kumar, V.; Ramakrishnan, M.; Kohli, S.; Kaur, R.; Thukral, A.; Bhardwaj, R.; Zheng, B. Castasterone attenuates insecticide induced phytotoxicity in mustard. Ecotoxicol. Environ. Saf. 2019, 179, 50–61. [Google Scholar] [CrossRef]
- Zhang, L.; Pei, Y.; Wang, H.; Jin, Z.; Liu, Z.; Qiao, Z.; Fang, H.; Zhang, Y. Hydrogen sulfide alleviates cadmium-induced cell death through restraining ROS accumulation in roots of Brassica rapa L. ssp. pekinensis. Oxid. Med. Cell Longev. 2014, 2015, 804603. [Google Scholar]
- Han, Y.; Yang, H.; Wu, M.; Yi, H. Enhanced drought tolerance of foxtail millet seedlings by sulfur dioxide fumigation. Ecotoxicol. Environ. Saf. 2019, 178, 9–16. [Google Scholar] [CrossRef]
- Li, L.; Lin, G.; Gu, H.; Yu, L.; Ni, C. Effects of Srxn1 on growth and Notch signalling of astrocyte induced by hydrogen peroxide. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1917–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Lin, L.; Zhong, S.; Cai, Y.; Zhang, F.; Wang, X.; Miao, R.; Zhang, B.; Gao, S.; Hu, X. Overexpression of novel lncRNA NLIPMT inhibits metastasis by reducing phosphorylated glycogen synthase kinase 3β in breast cancer. J. Cell Physiol. 2018, 234, 10698–10708. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, C.; Li, W.; Liang, X.; Ren, C.; Liu, W.; Yang, G.; Zhao, M.; Yang, T.; Li, X.; Han, D. Molecular Cloning and Characterization of MbMYB108, a Malus baccata MYB Transcription Factor Gene, with Functions in Tolerance to Cold and Drought Stress in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 4846. https://doi.org/10.3390/ijms23094846
Yao C, Li W, Liang X, Ren C, Liu W, Yang G, Zhao M, Yang T, Li X, Han D. Molecular Cloning and Characterization of MbMYB108, a Malus baccata MYB Transcription Factor Gene, with Functions in Tolerance to Cold and Drought Stress in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences. 2022; 23(9):4846. https://doi.org/10.3390/ijms23094846
Chicago/Turabian StyleYao, Chunya, Wenhui Li, Xiaoqi Liang, Chuankun Ren, Wanda Liu, Guohui Yang, Mengfei Zhao, Tianyu Yang, Xingguo Li, and Deguo Han. 2022. "Molecular Cloning and Characterization of MbMYB108, a Malus baccata MYB Transcription Factor Gene, with Functions in Tolerance to Cold and Drought Stress in Transgenic Arabidopsis thaliana" International Journal of Molecular Sciences 23, no. 9: 4846. https://doi.org/10.3390/ijms23094846