
Linoleic Acid Attenuates Denervation-Induced Skeletal Muscle Atrophy in Mice through Regulation of Reactive Oxygen Species-Dependent Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
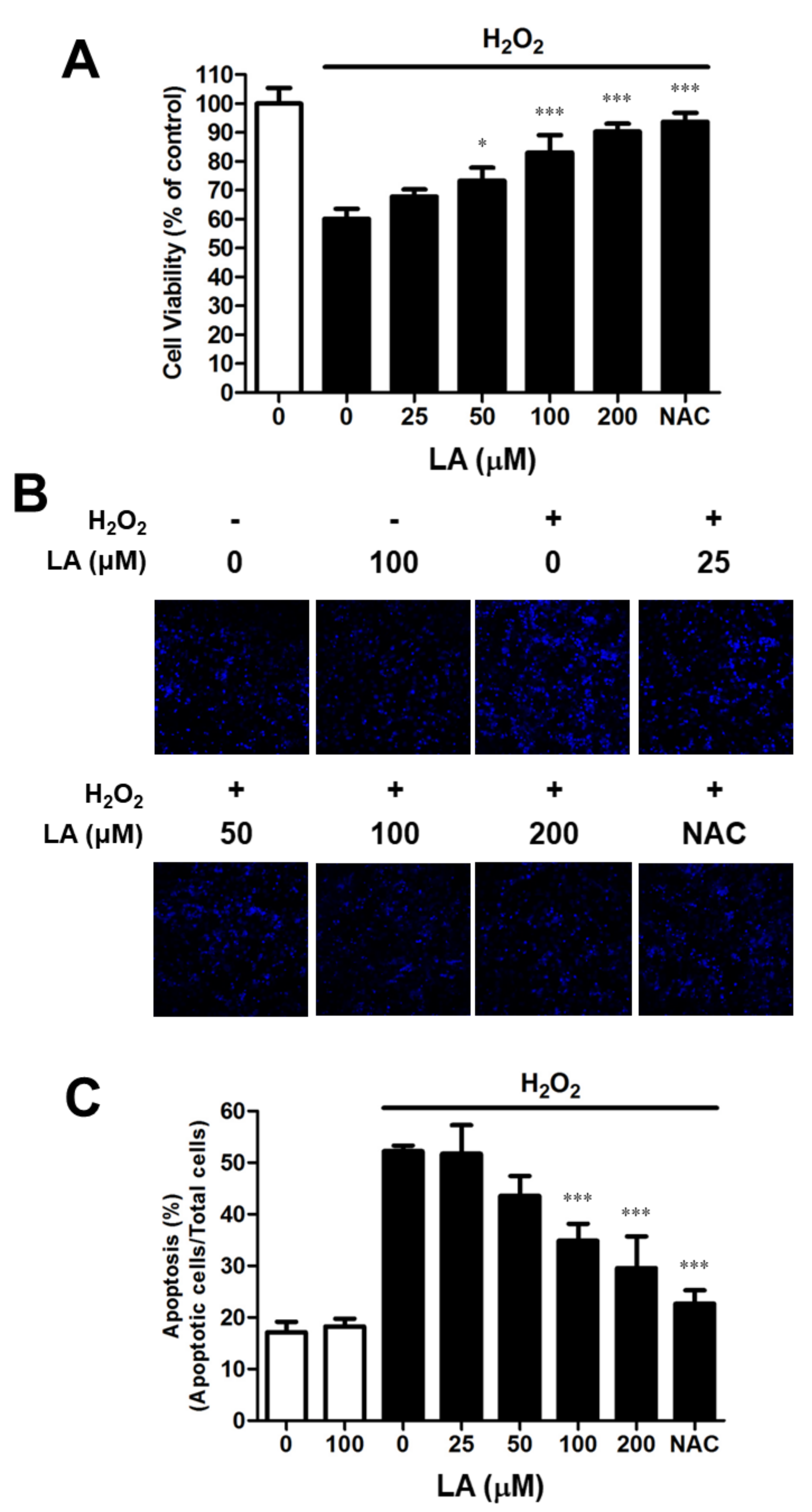
2.1. Linoleic Acid Attenuates the Cell Viability Decrease and Oxidative Stress-Mediated Apoptosis in C2C12 Myoblasts
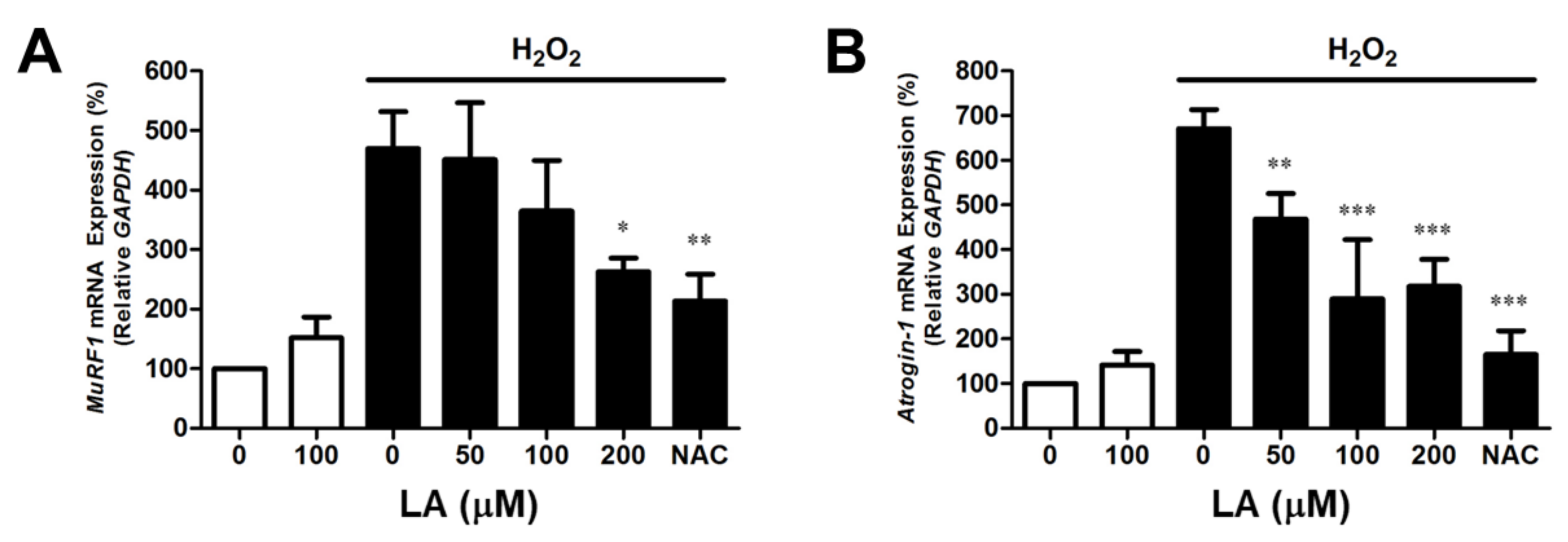
2.2. Linoleic Acid Decreases Oxidative Stress-Induced MuRF1 and Atrogin-1 mRNA Expression in C2C12 Myoblasts
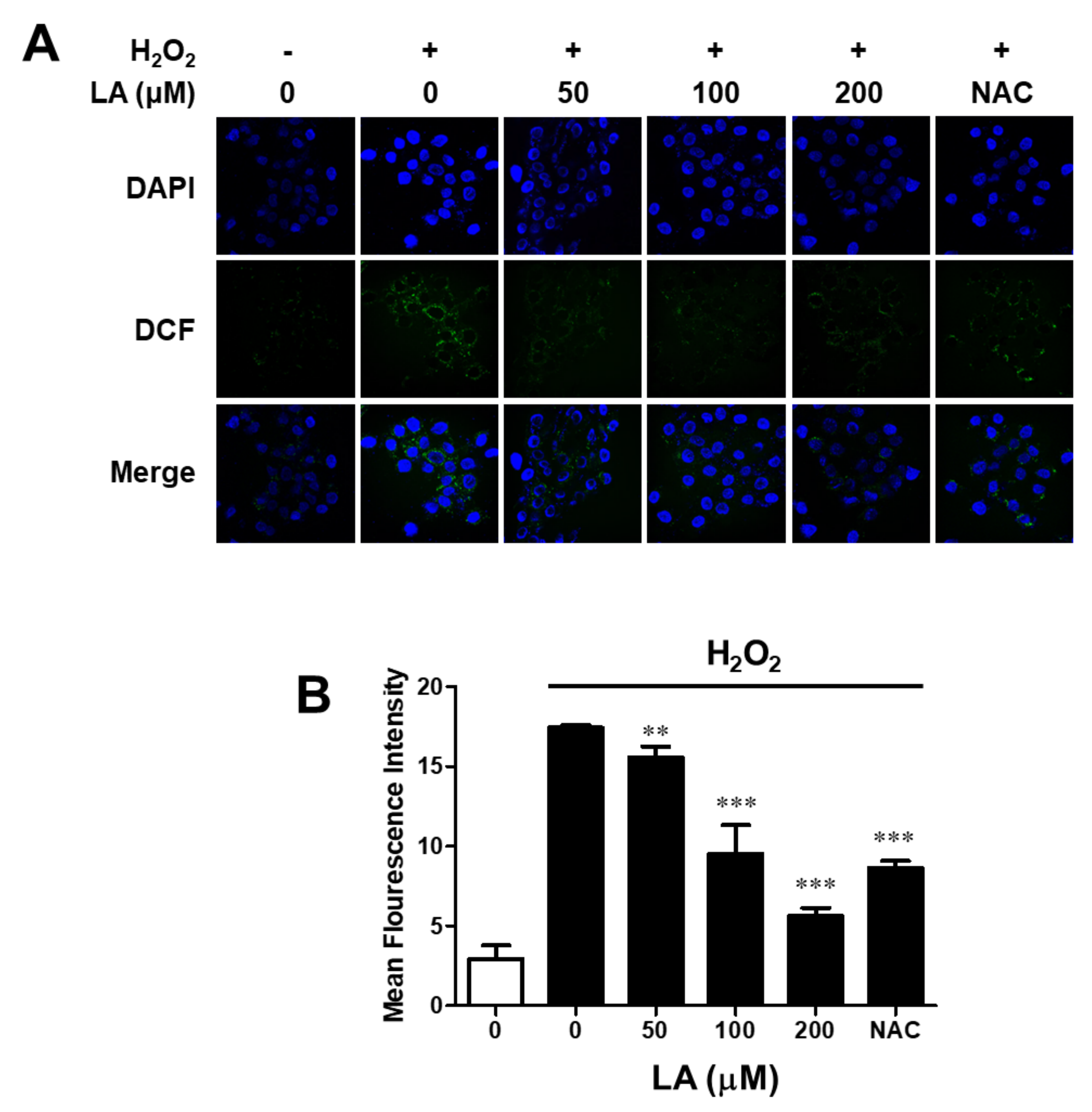
2.3. Linoleic Acid Decreases Intracellular Reactive Oxygen Species Synthesis in C2C12 Myoblasts
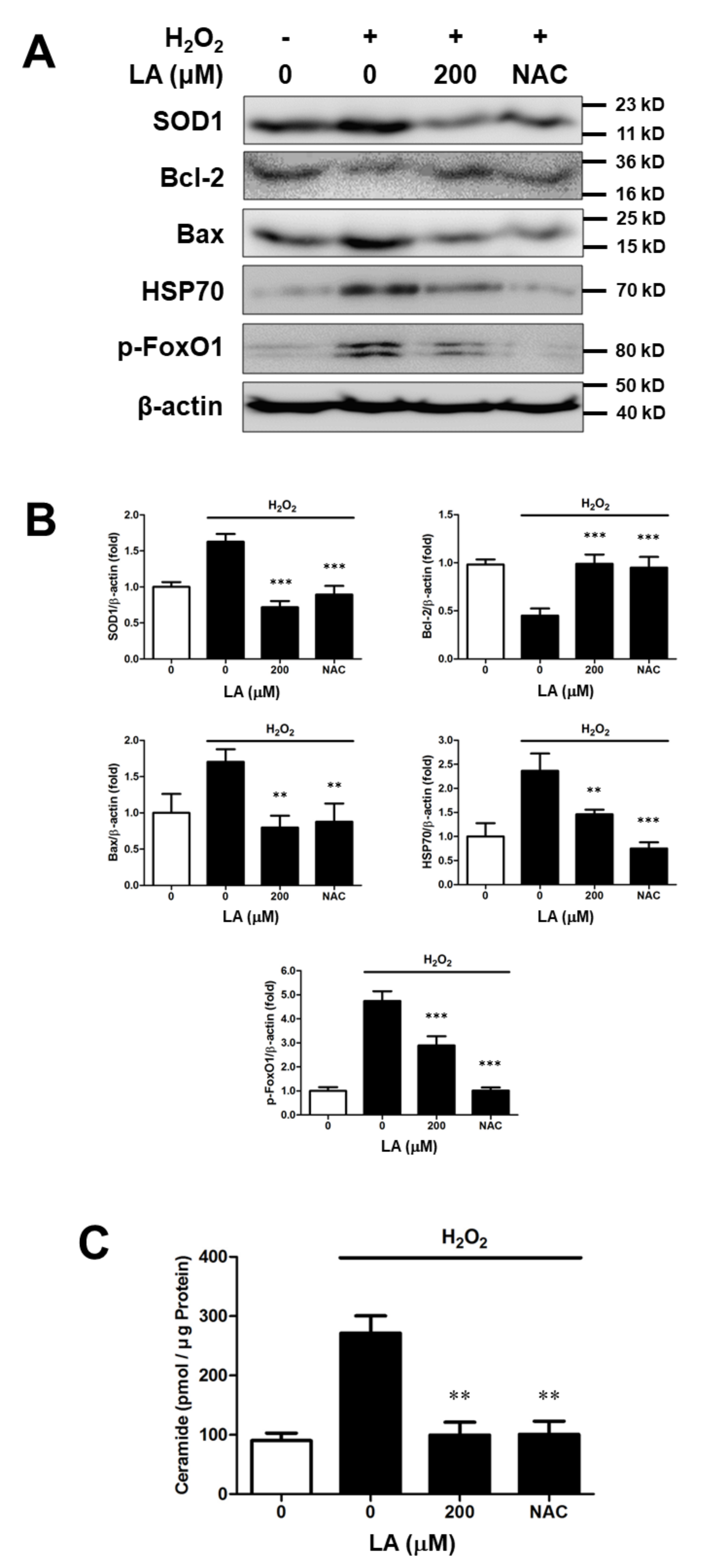
2.4. Effect of Linoleic Acid on the Altered Intracellular Protein Levels Induced by Oxidative Sress in C2C12 Moblasts
2.5. Linoleic Acid Decreases Intracellular Ceramide Level in Oxidative Stress-Induced C2C12 Myoblasts
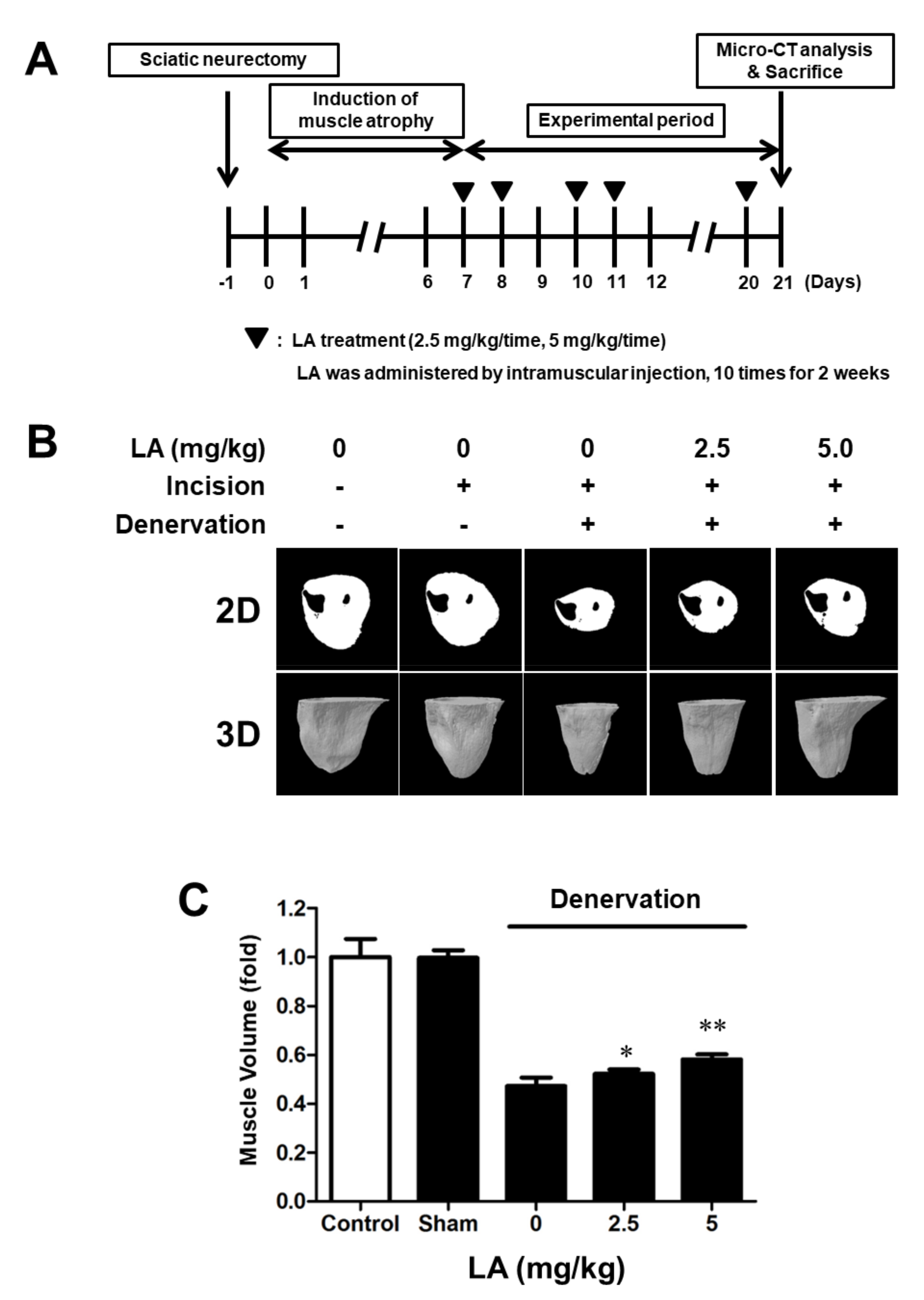
2.6. Linoleic Acid Reduces Muscle Volume on Sciatic Denervation-Induced Muscle Atrophy of C57BL/6 Mice
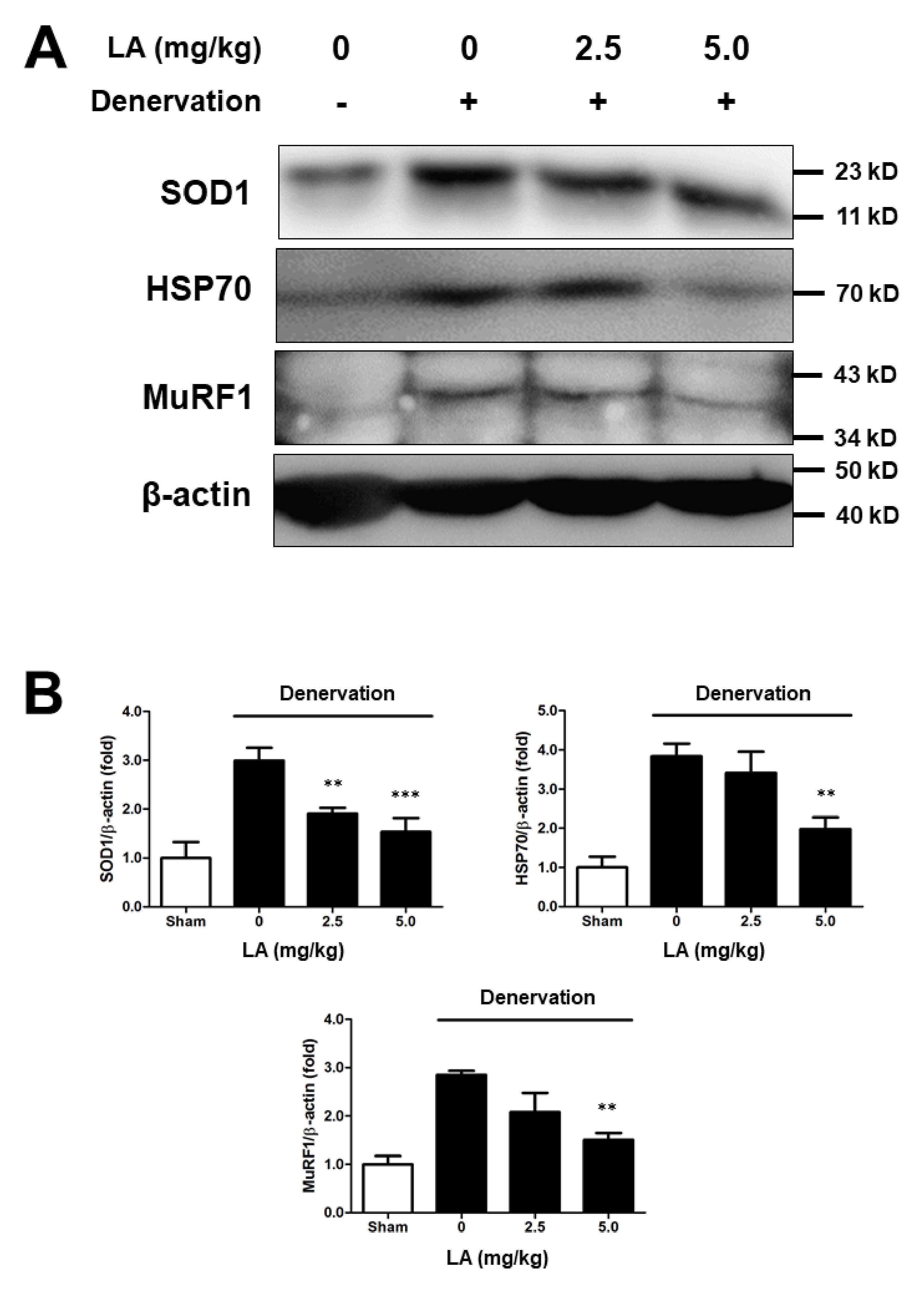
2.7. Linoleic Acid Decreases SOD1, HSP70, and MuRF1 Protein Expression on Sciatic Denervation-Induced Muscle Atrophy of C57BL/6 Mice
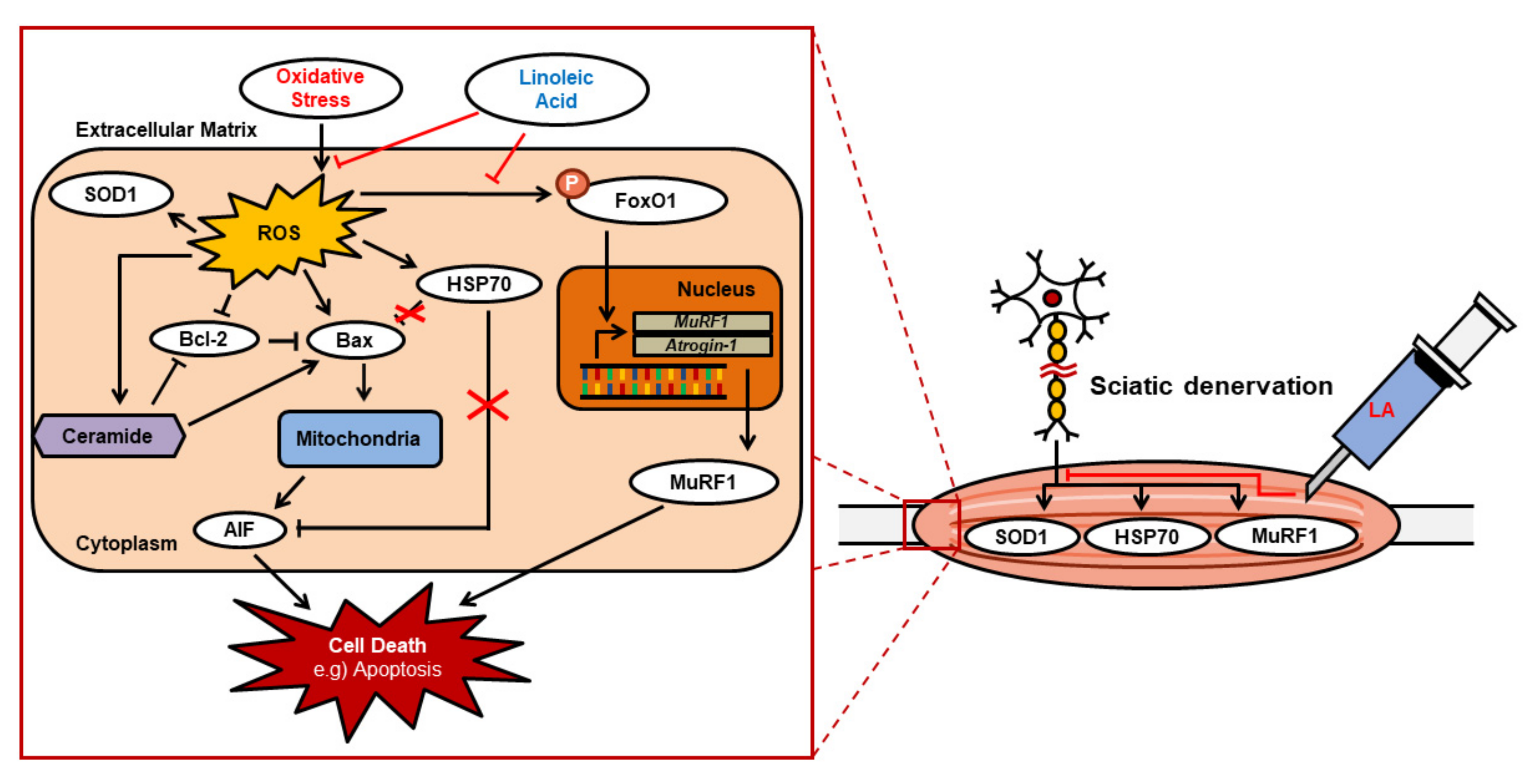
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Viability
4.4. Cytotoxicity
4.5. Evaluation of Apoptotic Cells
4.6. Quantitative Reverse-Transcriptase Polymerase Chain Reaction (qRT-PCR)
4.7. Detection of Intracellular ROS
4.8. Immunoblot Analysis
4.9. High-Performance Liquid Chromatography
4.10. Animal Experiment
4.11. Micro-Computed Tomography (micro-CT)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krebs, J.M.; Denney, R.M. Effect of atrophy and contractions on myogenin mRNA concentration in chick and rat myoblast omega muscle cells. In Vitro Cell. Dev. Biol. Anim. 1997, 33, 187–194. [Google Scholar] [CrossRef]
- Pompeani, N.; Rybalka, E.; Latchman, H.; Murphy, R.M.; Croft, K.; Hayes, A. Skeletal muscle atrophy in sedentary Zucker obese rats is not caused by calpain-mediated muscle damage or lipid peroxidation induced by oxidative stress. J. Negat. Results Biomed. 2014, 13, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervenakova, L.; Protas, I.I.; Hirano, A.; Votiakov, V.I.; Nedzved, M.K.; Kolomiets, N.D.; Taller, I.; Park, K.Y.; Sambuughin, N.; Gajdusek, D.C.; et al. Progressive muscular atrophy variant of familial amyotrophic lateral sclerosis (PMA/ALS). J. Neurol. Sci. 2000, 177, 124–130. [Google Scholar] [CrossRef]
- Gwag, T.; Lee, K.; Ju, H.; Shin, H.; Lee, J.W.; Choi, I. Stress and signaling responses of rat skeletal muscle to brief endurance exercise during hindlimb unloading: A catch-up process for atrophied muscle. Cell. Physiol. Biochem. 2009, 24, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Yeo, D.; Kang, C. Muscle disuse atrophy caused by discord of intracellular signaling. Antioxid. Redox Signal. 2020, 33, 727–744. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Li, Y.-P.; Chen, Y.; Li, A.S.; Reid, M.B. Hydrogen peroxide stimulates ubiquitin-conjugating activity and expression of genes for specific E2 and E3 proteins in skeletal muscle myotubes. Am. J. Physiol.-Cell Physiol. 2003, 285, C806–C812. [Google Scholar] [CrossRef] [Green Version]
- Powers, S.K.; Smuder, A.J.; Criswell, D.S. Mechanistic links between oxidative stress and disuse muscle atrophy. Antioxid. Redox Signal. 2011, 15, 2519–2528. [Google Scholar] [CrossRef] [Green Version]
- Smuder, A.J.; Kavazis, A.N.; Hudson, M.B.; Nelson, W.B.; Powers, S.K. Oxidation enhances myofibrillar protein degradation via calpain and caspase-3. Free Radic. Biol. Med. 2010, 49, 1152–1160. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Lewén, A.; Gasche, Y.; Yu, F.; Chan, P.H. Overexpression of SOD1 protects vulnerable motor neurons after spinal cord injury by attenuating mitochondrial cytochrome c release. FASEB J. 2002, 16, 1997–1999. [Google Scholar] [CrossRef]
- Tsang, C.K.; Liu, Y.; Thomas, J.; Zhang, Y.; Zheng, X.F. Superoxide dismutase 1 acts as a nuclear transcription factor to regulate oxidative stress resistance. Nat. Commun. 2014, 5, 3446–3456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, T.; Oishi, Y.; Higashida, K.; Higuchi, M.; Muraoka, I. Prolonged exercise training induces long-term enhancement of HSP70 expression in rat plantaris muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1557–R1563. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Kim, D.H.; Kim, Y.S.; Kim, T.J. Prevention of oxidative stress-induced apoptosis of C2C12 myoblasts by a Cichorium intybus root extract. Biosci. Biotechnol. Biochem. 2013, 77, 375–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanicolaou, K.N.; Izumiya, Y.; Walsh, K. Forkhead transcription factors and cardiovascular biology. Circ. Res. 2008, 102, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Schachter, T.N.; Shen, T.; Liu, Y.; Schneider, M.F. Kinetics of nuclear-cytoplasmic translocation of Foxo1 and Foxo3A in adult skeletal muscle fibers. Am. J. Physiol. Cell. Physiol. 2012, 303, C977–C990. [Google Scholar] [CrossRef] [Green Version]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Mobley, C.B.; Fox, C.D.; Ferguson, B.S.; Amin, R.H.; Dalbo, V.J.; Baier, S.; Rathmacher, J.A.; Wilson, J.M.; Roberts, M.D. L-leucine, beta-hydroxy-beta-methylbutyric acid (HMB) and creatine monohydrate prevent myostatin-induced Akirin-1/Mighty mRNA down-regulation and myotube atrophy. J. Int. Soc. Sports. Nutr. 2014, 11, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Frost, R.A.; Nystrom, G.J.; Jefferson, L.S.; Lang, C.H. Hormone, cytokine, and nutritional regulation of sepsis-induced increases in atrogin-1 and MuRF1 in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E501–E512. [Google Scholar] [CrossRef] [Green Version]
- Kritchevsky, D.; Tepper, S.A.; Story, J.A. Linoleic acid amides: Effect on cholesteremia and atherosclerosis. Lipids 1977, 12, 16–21. [Google Scholar] [CrossRef]
- Vasil’eva, E.M.; Ziabkina, A.G. Effect of linoleic acid in the diet on arterial pressure, ATPase activity and cyclic nucleotide level in the heart of rats with spontaneous hypertension. Vopr. Pitan. 1989, 3, 41–44. [Google Scholar]
- Hamilton, J.S.; Klett, E.L. Linoleic acid and the regulation of glucose homeostasis: A review of the evidence. Prostaglandins Leukot. Essent. Fat. Acids 2021, 175, 102366. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, H.; Meng, X.; Tong, P.; Liu, X. Biosynthesis of c9, t11-conjugated linoleic acid and the effect on characteristics in fermented soy milk. Food Chem. 2022, 368, 130866. [Google Scholar] [CrossRef] [PubMed]
- Pileggi, C.A.; Segovia, S.A.; Markworth, J.F.; Gray, C.; Zhang, X.D.; Milan, A.M.; Mitchell, C.J.; Barnett, M.P.G.; Roy, N.C.; Vickers, M.H. Maternal conjugated linoleic acid supplementation reverses high-fat diet-induced skeletal muscle atrophy and inflammation in adult male rat offspring. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2016, 310, R432–R439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poruk, K.E.; Davis, R.H.; Smart, A.L.; Chisum, B.S.; LaSalle, B.A.; Chan, G.M.; Gill, G.; Reyna, S.P.; Swoboda, K.J. Observational study of caloric and nutrient intake, bone density, and body composition in infants and children with spinal muscular atrophy type I. Neuromuscul. Disord. 2012, 22, 966–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moresi, V.; Williams, A.H.; Meadows, E.; Flynn, J.M.; Potthoff, M.J.; McAnally, J.; Shelton, J.M.; Backs, J.; Klein, W.H.; Richardson, J.A.; et al. Myogenin and class II HDACs control neurogenic muscle atrophy by inducing E3 ubiquitin ligases. Cell 2010, 143, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.J.; Lee, M.H.; Kim, W.J.; Chae, Y.; Iwasa, M.; Han, K.I.; Kim, W.J.; Kim, T.J. Effect of Heat-Killed Enterococcus faecalis, EF-2001 on C2C12 Myoblast Damage Induced by Oxidative Stress and Muscle Volume Decreased by Sciatic Denervation in C57BL/6 Mice. J. Life Sci. 2019, 29, 215–222. [Google Scholar]
- Singh, K.; Hood, D.A. Effect of denervation-induced muscle disuse on mitochondrial protein import. Am. J. Physiol. Cell Physiol. 2011, 300, C138–C145. [Google Scholar] [CrossRef] [Green Version]
- Haycock, J.W.; Mac Neil, S.; Mantle, D. Differential protein oxidation in Duchenne and Becker muscular dystrophy. Neuroreport 1998, 9, 2201–2207. [Google Scholar] [CrossRef]
- Yotani, K.; Nakamoto, H.; Ikudome, S.; Yuki, A. Muscle contraction and relaxation-response time in response to on or off status of visual stimulus. J. Physiol. Anthropol. 2014, 33, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.C. Tarnopolsky MA. Patients with dystrophinopathyshow evidence of increased oxidative stress. Free Radic. Biol. Med. 2003, 34, 1217–1220. [Google Scholar] [CrossRef]
- Haycock, J.W.; MacNeil, S.; Jones, P.; Harris, J.B.; Mantle, D. Oxidative damage to muscle protein in Duchenne muscular dystrophy. NeuroReport 1996, 8, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Petry, É.R.; Dresch, D.D.F.; Carvalho, C.; Medeiros, P.C.; Rosa, T.G.; Oliveira, C.M.D.; Martins, L.A.M.; Schemitt, E.; Bona, S.; Guma, F.C.R.; et al. Oral glutamine supplementation attenuates inflammation and oxidative stress-mediated skeletal muscle protein content degradation in immobilized rats: Role of 70 kDa heat shock protein. Free Radic. Biol. Med. 2019, 145, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Landolfo, S.; Politi, H.; Angelozzi, D.; Mannazzu, I. ROS accumulation and oxidative damage to cell structures in Saccharomyces cerevisiae wine strains during fermentation of high-sugar-containing medium. Biochim. Biophys. Acta 2008, 1780, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Andrey, J.S.; José, R.P.; Marko, D.P.; Gisela, A.; Andrea, V. Oxidative Stress in Muscle Diseases: Current and Future Therapy 2019. Oxid. Med. Cell. Longev. 2020, 2020, 6030417. [Google Scholar]
- Lee, Y.H.; Kim, W.J.; Lee, M.H.; Kim, S.Y.; Seo, D.H.; Kim, H.S.; Gelinsky, M.; Kim, T.J. Anti-skeletal muscle atrophy effect of Oenothera odorata root extract via reactive oxygen species-dependent signaling pathways in cellular and mouse model. Biosci. Biotechnol. Biochem. 2015, 80, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.M.; Decker, E.A. Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 1994, 34, 403–426. [Google Scholar] [CrossRef]
- Nishida, H.; Ichikawa, H.; Konishi, T. Shengmai-san enhancesantioxidant potential in C2C12 myoblasts through the inductionof intracellular glutathione peroxidase. J. Pharmacol. Sci. 2007, 105, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Joza, N.; Oudit, G.Y.; Brown, D.; Bénit, P.; Kassiri, Z.; Vahsen, N.; Benoit, L.; Patel, M.M.; Nowikovsky, K.; Vassault, A.; et al. Muscle-specific loss of apoptosis-inducing factor leads to mitochondrial dysfunction, skeletal muscle atrophy, and dilated cardiomyopathy. Mol. Cell. Biol. 2005, 25, 10261–10272. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, D.; Nojiri, H.; Saita, Y.; Kobayashi, K.; Watanabe, K.; Ozawa, Y.; Koike, M.; Asou, Y.; Takaku, T.; Kaneko, K.; et al. Cytoplasmic reactive oxygen species and SOD1 regulate bone mass during mechanical unloading. J. Bone Miner. Res. 2013, 28, 2368–2380. [Google Scholar] [CrossRef]
- Senf, S.M.; Dodd, S.L.; McClung, J.M.; Judge, A.R. Hsp70 overexpression inhibits NF-κB and Foxo3a transcriptional activities and prevents skeletal muscle atrophy. FASEB J. 2008, 22, 3836–3845. [Google Scholar] [CrossRef]
- Léger, B.; Cartoni, R.; Praz, M.; Lamon, S.; Dériaz, O.; Crettenand, A.; Gobelet, C.; Rohmer, P.; Konzelmann, M.; Luthi, F.; et al. Akt signalling through GSK-3beta, mTOR and Foxo1 is involved in human skeletal muscle hypertrophy and atrophy. J. Physiol. 2006, 576, 923–933. [Google Scholar] [CrossRef]
- Fjällström, A.K.; Evertsson, K.; Norrby, M.; Tågerud, S. Forkhead box O1 and muscle RING finger 1 protein expression in atrophic and hypertrophic denervated mouse skeletal muscle. J. Mol. Signal. 2014, 9, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Jiao, H.; Zhao, J.; Wang, X.; Lin, H. Glucocorticoids enhance muscle proteolysis through a myostatin-dependent pathway at the early stage. PLoS ONE 2016, 11, e0156225. [Google Scholar] [CrossRef] [PubMed]
- Tacchi, L.; Bickerdike, R.; Secombes, C.J.; Martin, S.A. Muscle-specific RING finger (MuRF) cDNAs in Atlantic salmon (Salmo salar) and their role as regulators of muscle protein degradation. Mar. Biotechnol. 2012, 14, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Natanek, S.A.; Riddoch-Contreras, J.; Marsh, G.S.; Hopkinson, N.S.; Moxham, J.; Man, W.D.; Kemp, P.R.; Polkey, M.I. MuRF-1 and atrogin-1 protein expression and quadriceps fiber size and muscle mass in stable patients with COPD. COPD 2013, 10, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Jang, M.; Park, R.; Jo, D.; Choi, I.; Choe, J.; Oh, W.K.; Park, J. Conessine Treatment Reduces Dexamethasone-Induced Muscle Atrophy by Regulating MuRF1 and Atrogin-1 Expression. J. Microbiol. Biotechnol. 2018, 28, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Geilen, C.C.; Wieder, T.; Orfanos, C.E. Ceramide signalling: Regulatory role in cell proliferation, differentiation and apoptosis in human epidermis. Arch. Dermatol. Res. 1997, 289, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Metkar, S.S.; Anand, M.; Manna, P.P.; Naresh, K.N.; Nadkarni, J.J. Ceramide-induced apoptosis in fas-resistant Hodgkin’s disease cell lines is caspase independent. Exp. Cell Res. 2000, 255, 18–29. [Google Scholar] [CrossRef]
- Mebarek, S.; Komati, H.; Naro, F.; Zeiller, C.; Alvisi, M.; Lagarde, M.; Prigent, A.F.; Némoz, G. Inhibition of de novoceramide synthesis upregulates phospholipase D and enhancesmyogenic differentiation. J. Cell Sci. 2007, 120, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Meadows, K.A.; Holly, J.M.; Stewart, C.E. Tumor necrosis factorα-induced apoptosis is associated with suppression of insulin-like growth factor binding protein-5 secretion in differentiating murine skeletal myoblasts. J. Cell. Physiol. 2000, 183, 330–337. [Google Scholar] [CrossRef]
- Strle, K.; Broussard, S.R.; McCusker, R.H.; Shen, W.H.; Johnson, R.W.; Freund, G.G.; Dantzer, R.; Kelley, K.W. Proinflammatory cytokine impairment of insulin-like growth factor I-induced protein synthesis in skeletal muscle myoblasts requires ceramide. Endocrinology 2004, 145, 4592–4602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berk, M.; Malhi, G.S.; Gray, L.J.; Dean, O.M. The promise of N-acetylcysteine in neuropsychiatry. Trends Pharmacol. Sci. 2013, 34, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Michelucci, A.; Paolini, C.; Canato, M.; Wei-Lapierre, L.; Pietrangelo, L.; De Marco, A.; Reggiani, C.; Dirksen, R.T.; Protasi, F. Antioxidants protect calsequestrin-1 knockout mice from halothane- and heat-induced sudden death. Anesthesiology 2015, 123, 603–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelucci, A.; De Marco, A.; Guarnier, F.A.; Protasi, F.; Boncompagni, S. Antioxidant Treatment Reduces Formation of Structural Cores and Improves Muscle Function in RYR1Y522S/WT Mice. Oxid. Med. Cell. Longev. 2017, 2017, 6792694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Miura, M.; Itokawa, Y. Antioxidant enzyme systems in skeletal muscle atrophied by immobilization. Pflüg. Arch. 1993, 422, 404–406. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Nakagaki, I.; Sasaki, S.; Hori, S.; Itokawa, Y. Mechanism of oxidative stress in skeletal muscle atrophied by immobilization. Am. J. Physiol. 1993, 265, E839–E844. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Seo, D.H.; Park, J.H.; Kabayama, K.; Opitz, J.; Lee, K.H.; Kim, H.S.; Kim, T.J. Effect of Oenothera odorata root extract on microgravity and disuse-induced muscle atrophy. Evid. Based Complement. Altern. Med. 2015, 2015, 130513. [Google Scholar] [CrossRef] [Green Version]
- Blumenthal, M. The ABC Clinical Guide to Herbs; American Botanical Council: Austin, TX, USA, 2003; pp. 123–134. [Google Scholar]
- Kim, H.Y.; Oh, H.; Li, X.; Cho, K.W.; Kang, D.G.; Lee, H.S. Ethanol extract of seeds of Oenothera odorata induces vasorelaxation via endothelium-dependent NO-cGMP signaling through activation of Akt-eNOS-sGC pathway. J. Ethnopharmacol. 2011, 133, 315–323. [Google Scholar] [CrossRef]
- Chen, W.; Jiang, T.; Wang, H.; Tao, S.; Lau, A.; Fang, D.; Zhang, D.D. Does Nrf2 contribute to p53-mediated control of cell survival and death? Antioxid. Redox Signal. 2012, 17, 1670–1675. [Google Scholar] [CrossRef] [Green Version]
- Bensaad, K.; Vousden, K.H. Savior and slayer: The two faces of p53. Nat. Med. 2005, 11, 1278–1279. [Google Scholar] [CrossRef]
- Faraonio, R.; Vergara, P.; Di Marzo, D.; Pierantoni, M.G.; Napolitano, M.; Russo, T.; Cimino, F. p53 suppresses the Nrf2-dependent transcription of antioxidant response genes. J. Biol. Chem. 2006, 281, 39776–39784. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-H.; Lee, J.-H.; Kim, W.-J.; Kim, S.H.; Kim, S.-Y.; Kim, H.S.; Kim, T.-J. Linoleic Acid Attenuates Denervation-Induced Skeletal Muscle Atrophy in Mice through Regulation of Reactive Oxygen Species-Dependent Signaling. Int. J. Mol. Sci. 2022, 23, 4778. https://doi.org/10.3390/ijms23094778
Lee M-H, Lee J-H, Kim W-J, Kim SH, Kim S-Y, Kim HS, Kim T-J. Linoleic Acid Attenuates Denervation-Induced Skeletal Muscle Atrophy in Mice through Regulation of Reactive Oxygen Species-Dependent Signaling. International Journal of Molecular Sciences. 2022; 23(9):4778. https://doi.org/10.3390/ijms23094778
Chicago/Turabian StyleLee, Myung-Hun, Jin-Ho Lee, Wan-Joong Kim, Seo Ho Kim, Sun-Young Kim, Han Sung Kim, and Tack-Joong Kim. 2022. "Linoleic Acid Attenuates Denervation-Induced Skeletal Muscle Atrophy in Mice through Regulation of Reactive Oxygen Species-Dependent Signaling" International Journal of Molecular Sciences 23, no. 9: 4778. https://doi.org/10.3390/ijms23094778