
Cytotoxicity of Newly Synthesized Quinazoline–Sulfonamide Derivatives in Human Leukemia Cell Lines and Their Effect on Hematopoiesis in Zebrafish Embryos

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of Synthesized Compounds on Leukemia Cell Viability
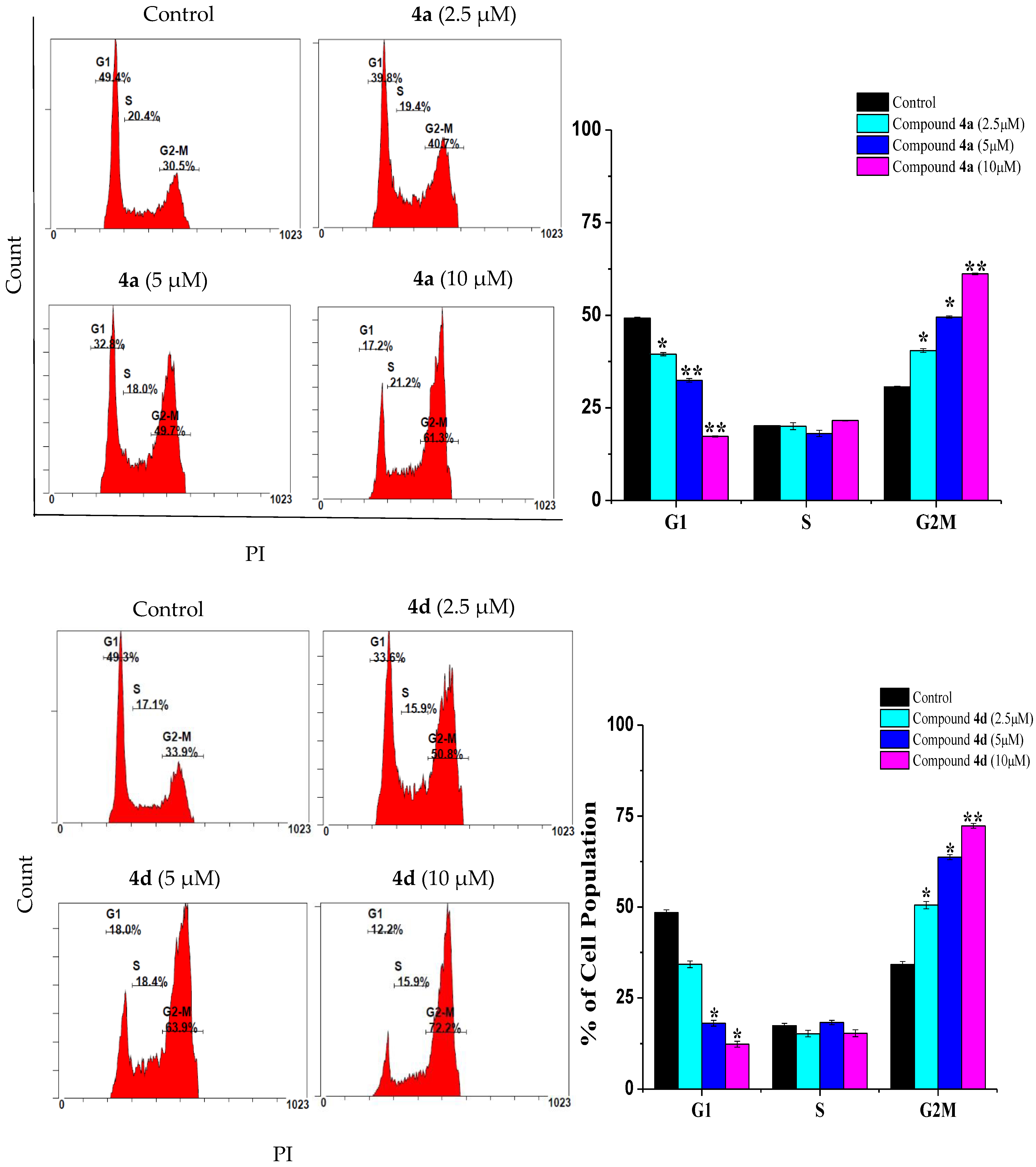
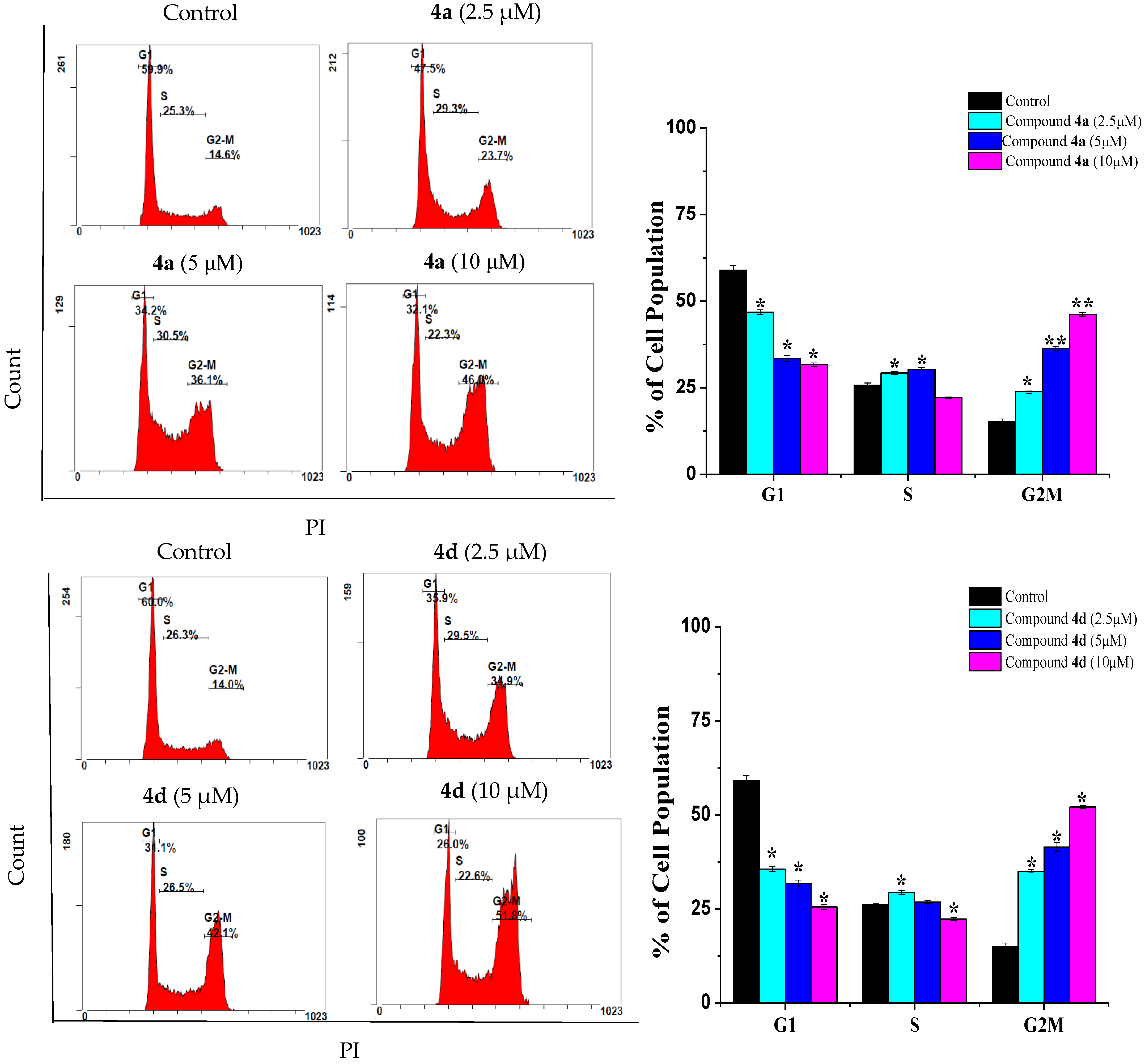
2.2. Compounds 4a and 4d Caused G2/M Cell Cycle Arrest
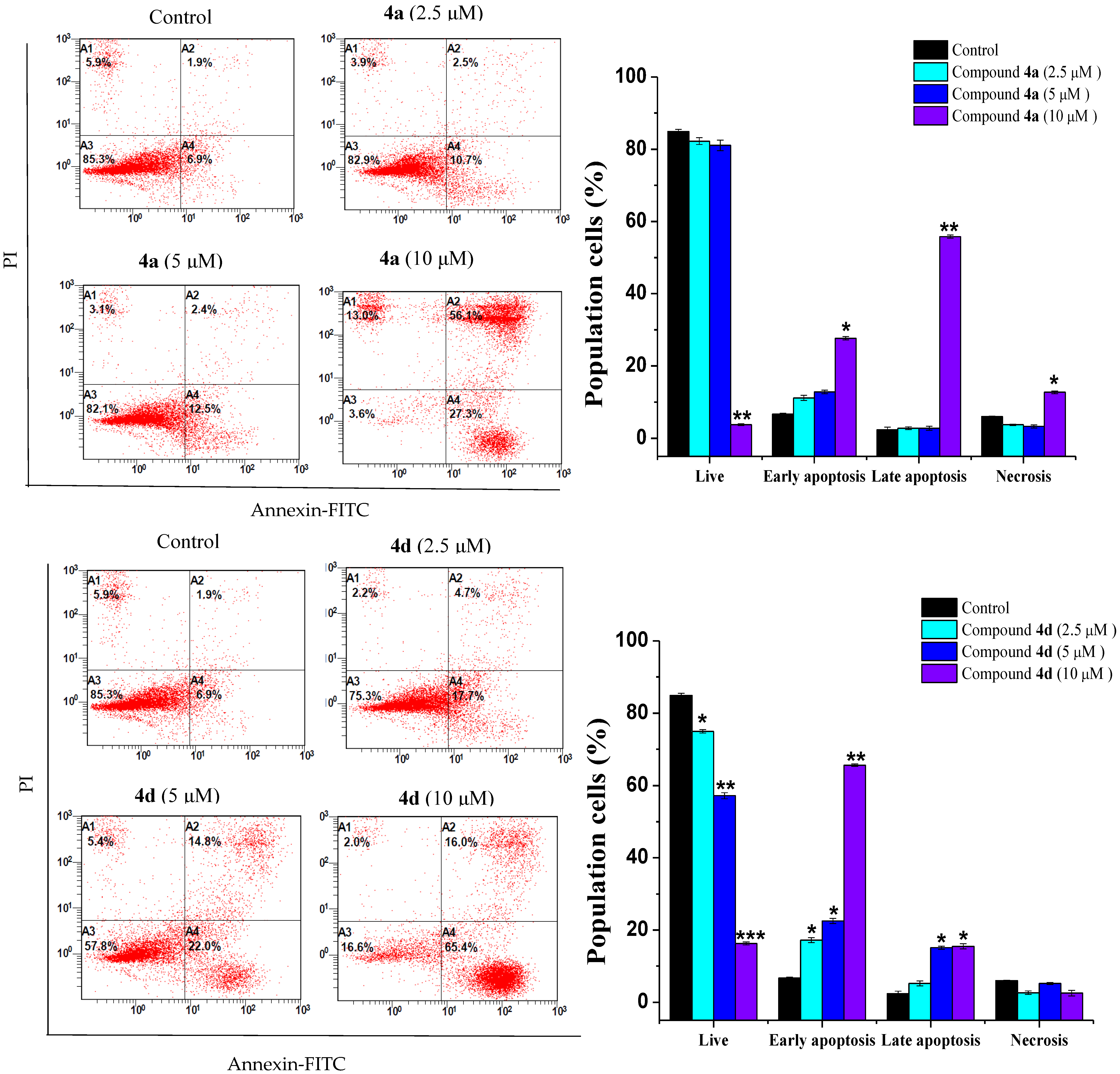
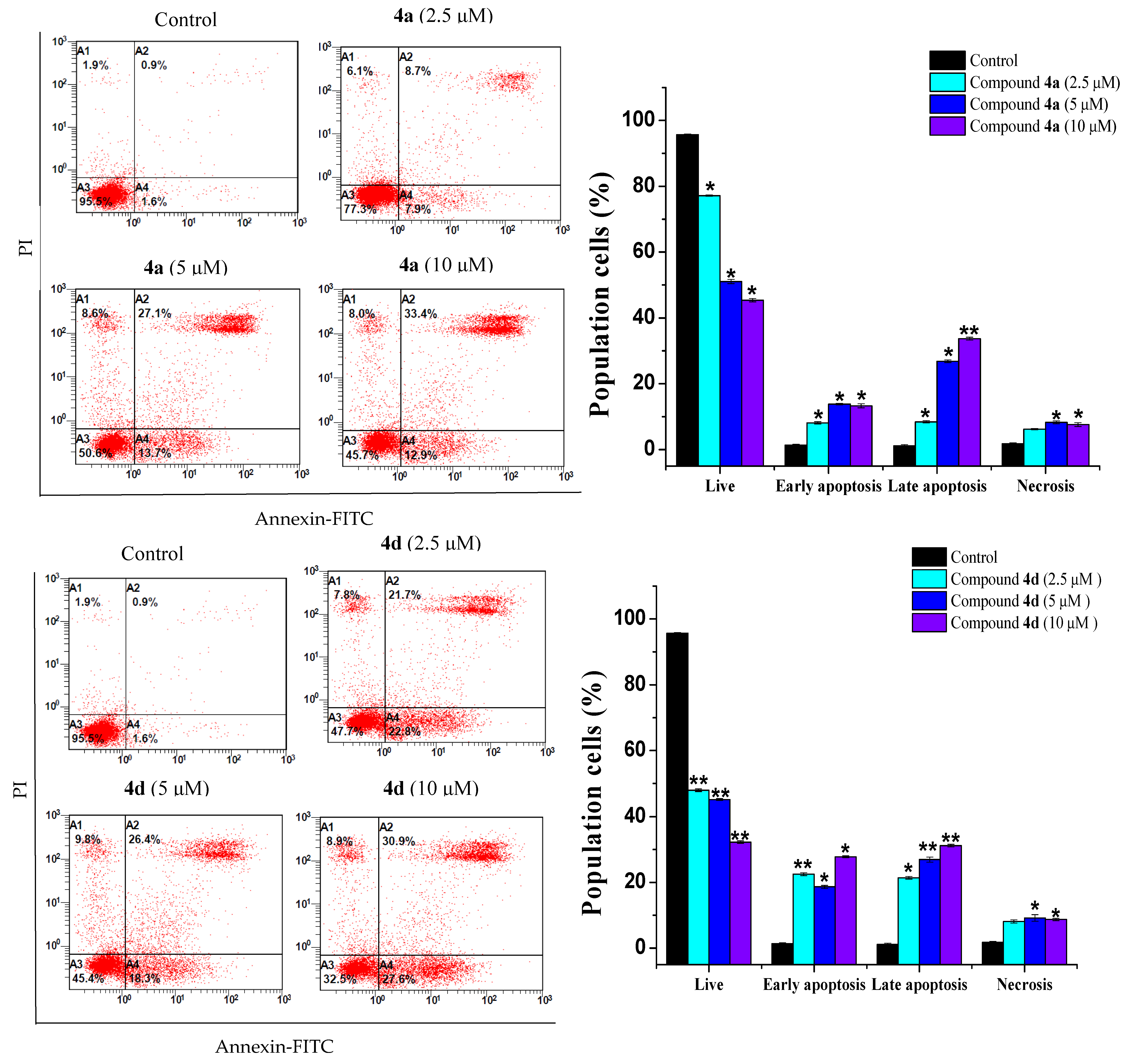
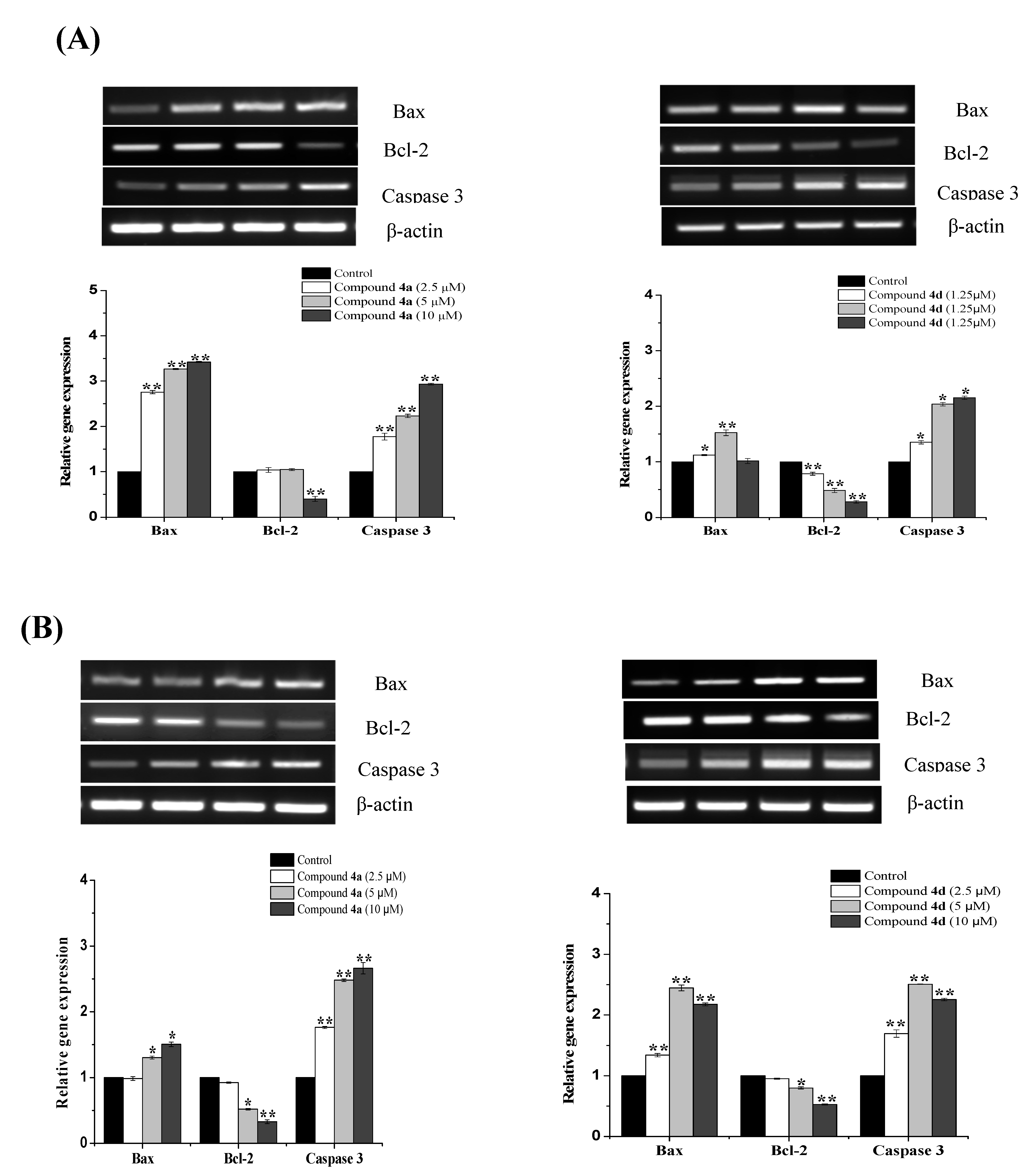
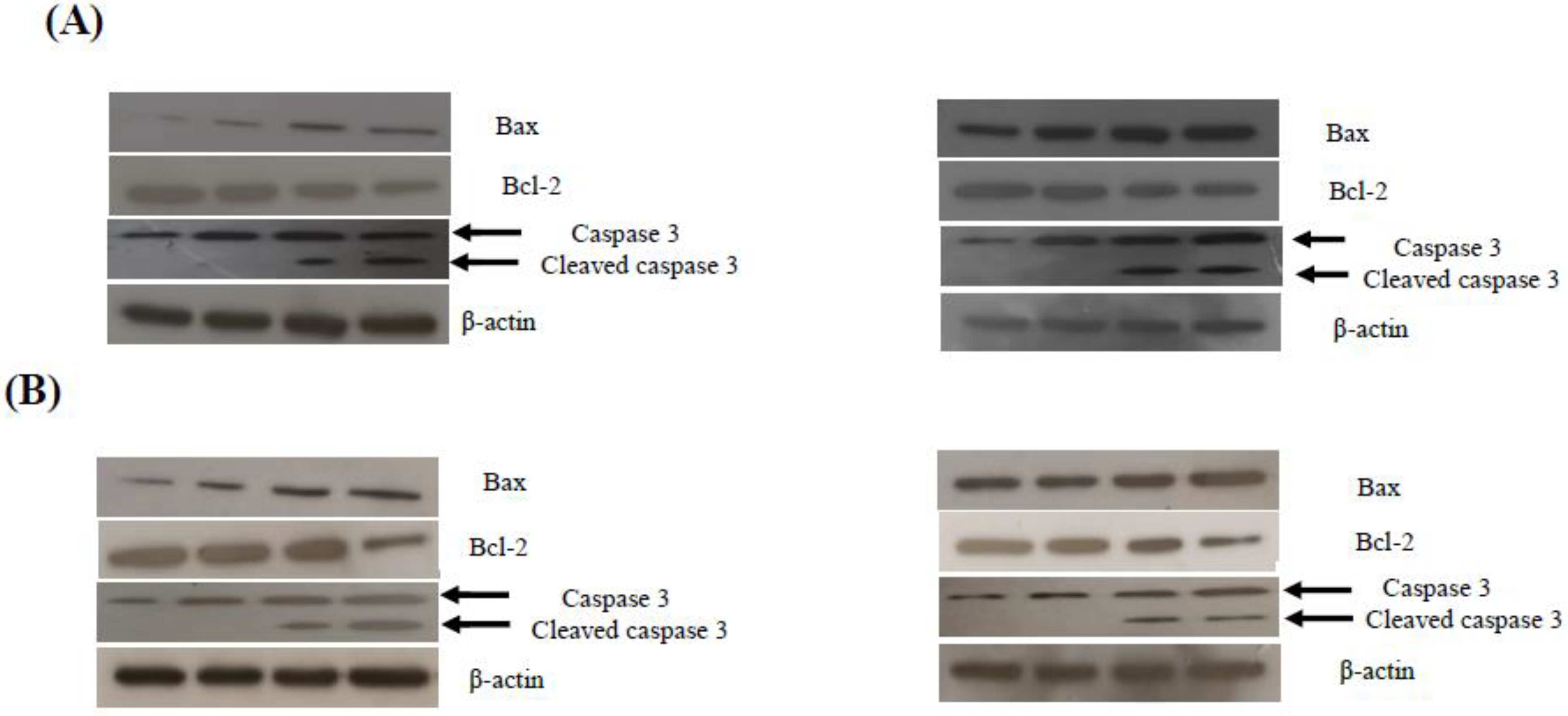
2.3. Compounds 4a and 4d Inhibited Leukemic Cancer Cell Proliferation through Apoptosis Induction
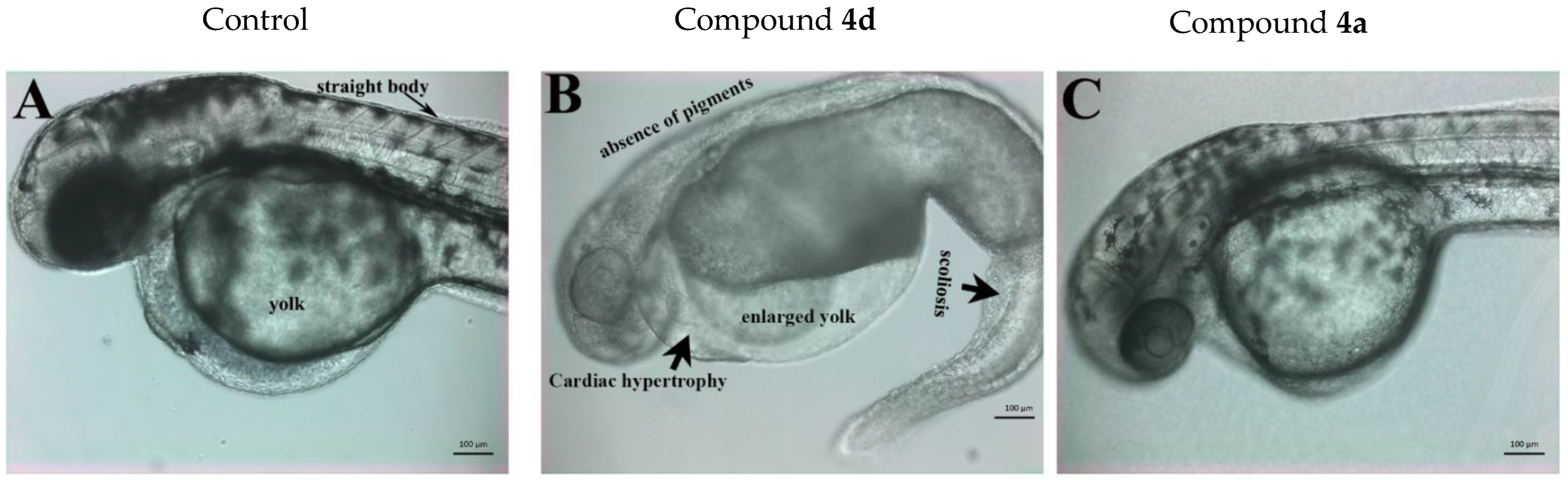
2.4. Determination of the In Vivo Toxicity of Compounds 4a and 4d in Zebrafish Embryos
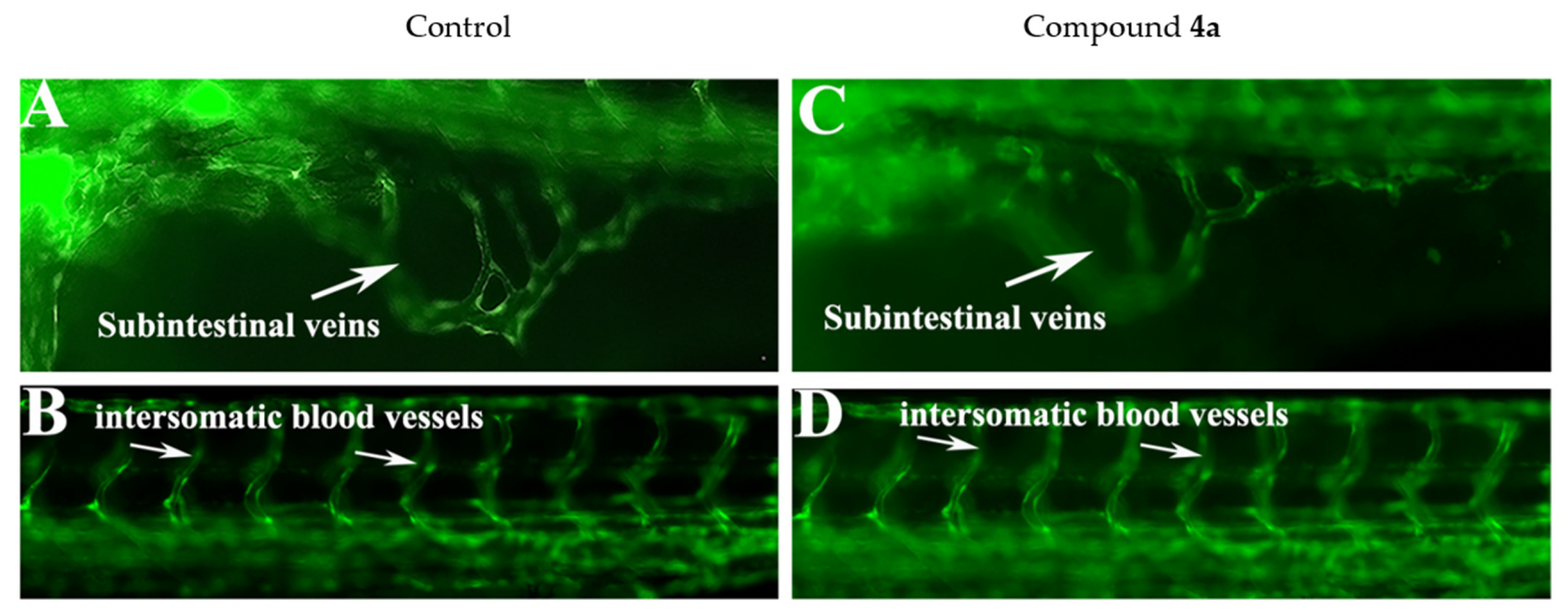
2.5. Compound 4a Affected the Haemopoiesis Process in Developing Zebrafish Embryos without Inducing Developmental Toxicity
3. Conclusions
4. Materials and Methods
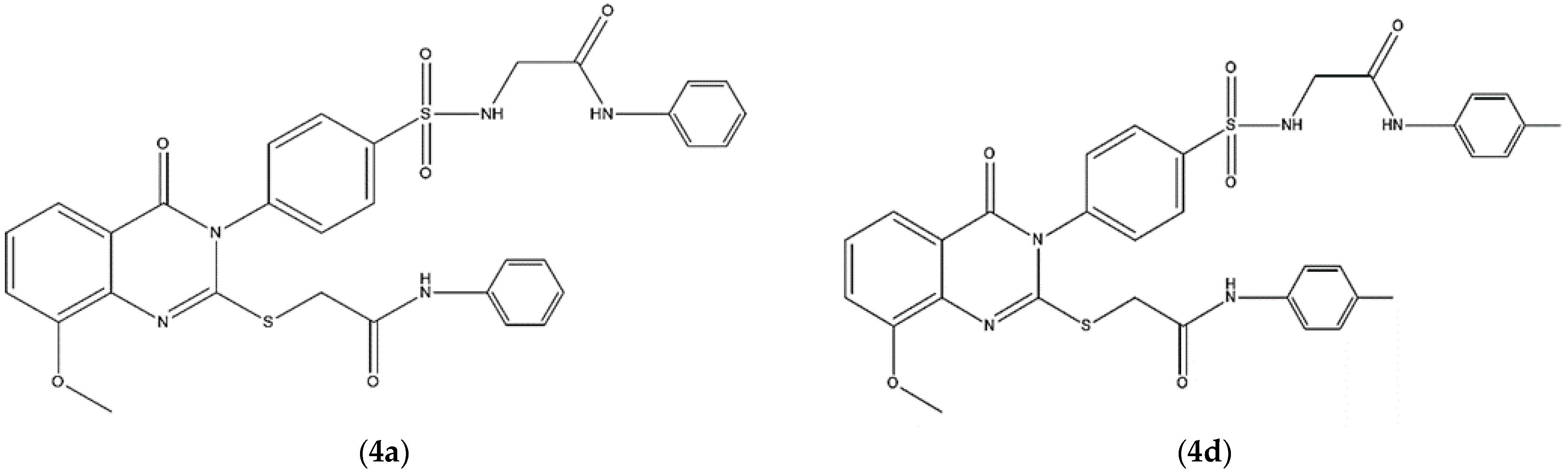
4.1. Compounds
4.2. Cell Lines and Cell Viability Assay
4.3. Cell Cycle Analysis
4.4. Detection of Apoptosis by Flow Cytometry
4.5. Extraction of RNA and Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
4.6. Western Blotting
4.7. In Vivo Zebrafish Study
4.7.1. Animal Treatment
4.7.2. Preparation of Compounds and Embryo Treatments
4.7.3. Hemoglobin Staining of Live Zebrafish Embryos
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Nemkov, T.; D’Alessandro, A.; Reisz, J.A. Metabolic underpinnings of leukemia pathology and treatment. Cancer Rep. 2019, 2, e1139. [Google Scholar] [CrossRef] [PubMed]
- Wendtner, C.M.; Gregor, M. Current perspectives on the role of chemotherapy in chronic lymphocytic leukemia. Leuk. Lymphoma 2018, 59, 300–310. [Google Scholar] [CrossRef]
- Maher, T.; Ahmad Raus, R.; Daddiouaissa, D.; Ahmad, F.; Adzhar, N.S.; Latif, E.S.; Abdulhafiz, F.; Mohammed, A. Medicinal Plants with Anti-Leukemic Effects: A Review. Molecules 2021, 26, 2741. [Google Scholar] [CrossRef]
- Alagarsamy, V.; Chitra, K.; Saravanan, G.; Solomon, V.R.; Sulthana, M.T.; Narendhar, B. An overview of quinazolines: Pharmacological significance and recent developments. Eur. J. Med. Chem. 2018, 151, 628–685. [Google Scholar] [CrossRef]
- Poudapally, S.; Battu, S.; Velatooru, L.R.; Bethu, M.S.; Janapala, V.R.; Sharma, S.; Sen, S.; Pottabathini, N.; Iska, V.B.R.; Katangoor, V. Synthesis and biological evaluation of novel quinazoline-sulfonamides as anti-cancer agents. Bioorg. Med. Chem. Lett. 2017, 27, 1923–1928. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Z.; Xia, S.; Liu, Q.; Gou, S. Design, synthesis and biological evaluation of sulfamoylphenyl-quinazoline derivatives as potential EGFR/CAIX dual inhibitors. Eur. J. Med. Chem. 2021, 216, 113300. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Sharma, V.; Alam, O.; Manaithiya, A.; Alam, P.; Kahksha, J.; Alam, M.T.; Imran, M. Novel quinazoline-based EGFR kinase inhibitors: A review focussing on SAR and molecular docking studies (2015–2019). Eur. J. Med. Chem. 2020, 204, 112640. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Malhotra, A. Therapeutic progression of quinazolines as targeted chemotherapeutic agents. Eur. J. Med. Chem. 2021, 211, 113016. [Google Scholar] [CrossRef] [PubMed]
- Mehndiratta, S.; Sapra, S.; Singh, G.; Singh, M.; Nepali, K. Quinazolines as Apoptosis Inducers and Inhibitors: A Review of Patent Literature. Recent. Pat. Anticancer Drug Discov. 2016, 11, 2–66. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, A.S.; Ghorab, M.M.; Nasr, F.A.; Ahmed, M.Z.; Al-Mishari, A.A.; Attia, S.M. The Antiproliferative and Apoptotic Effects of a Novel Quinazoline Carrying Substituted-Sulfonamides: In Vitro and Molecular Docking Study. Molecules 2022, 27, 981. [Google Scholar] [CrossRef] [PubMed]
- de Pater, E.; Trompouki, E. Bloody Zebrafish: Novel Methods in Normal and Malignant Hematopoiesis. Front. Cell Dev. Biol. 2018, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yu, S.H.; Zhen, W.X.; Cheng, T.; Wang, D.; Lin, J.B.; Wu, Y.H.; Wang, Y.F.; Chen, Y.; Shu, L.P.; et al. Tanshinone I, a new EZH2 inhibitor restricts normal and malignant hematopoiesis through upregulation of MMP9 and ABCG2. Theranostics 2021, 11, 6891–6904. [Google Scholar] [CrossRef]
- Jing, L.; Zon, L.I. Zebrafish as a model for normal and malignant hematopoiesis. Dis. Model. Mech. 2011, 4, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, G.I.; Harper, J.W. Anticancer drug targets: Cell cycle and checkpoint control. J. Clin. Investig. 1999, 104, 1645–1653. [Google Scholar] [CrossRef]
- Zuryń, A.; Litwiniec, A.; Gackowska, L.; Pawlik, A.; Grzanka, A.A.; Grzanka, A. Expression of cyclin A, B1 and D1 after induction of cell cycle arrest in the Jurkat cell line exposed to doxorubicin. Cell Biol. Int. 2012, 36, 1129–1135. [Google Scholar] [CrossRef]
- Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Doxorubicin induces apoptosis in Jurkat cells by mitochondria-dependent and mitochondria-independent mechanisms under normoxic and hypoxic conditions. Anticancer Drugs 2015, 26, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, H.; Jamshidi, A.; Ghatee, M.A.; Mazhab-Jafari, K.; Khorasani, M.; Rahmati, M.; Mohammadi, S. PPARγ activation by pioglitazone enhances the anti-proliferative effects of doxorubicin on pro-monocytic THP-1 leukemia cells via inducing apoptosis and G2/M cell cycle arrest. J. Recept. Signal. Transduct. Res. 2021, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shagufta; Ahmad, I. An insight into the therapeutic potential of quinazoline derivatives as anticancer agents. Medchemcomm 2017, 8, 871–885. [Google Scholar] [CrossRef]
- Campbell, K.J.; Tait, S.W.G. Targeting BCL-2 regulated apoptosis in cancer. Open Biol. 2018, 8, 180002. [Google Scholar] [CrossRef]
- Walsh, J.G.; Cullen, S.P.; Sheridan, C.; Lüthi, A.U.; Gerner, C.; Martin, S.J. Executioner caspase-3 and caspase-7 are functionally distinct proteases. Proc. Natl. Acad. Sci. USA 2008, 105, 12815–12819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, T.V.; Zon, L.I. Swimming into the future of drug discovery: In vivo chemical screens in zebrafish. ACS Chem. Biol. 2010, 5, 159–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.T.; Zon, L.I. Zebrafish blood stem cells. J. Cell Biochem. 2009, 108, 35–42. [Google Scholar] [CrossRef]
- Rasighaemi, P.; Basheer, F.; Liongue, C.; Ward, A.C. Zebrafish as a model for leukemia and other hematopoietic disorders. J. Hematol. Oncol. 2015, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Molina, B.; Chavez, J.; Grainger, S. Zebrafish models of acute leukemias: Current models and future directions. Wiley Interdiscip. Rev. Dev. Biol. 2021, 10, e400. [Google Scholar] [CrossRef] [PubMed]
- Mizgirev, I.V.; Revskoy, S. A new zebrafish model for experimental leukemia therapy. Cancer Biol. Ther. 2010, 9, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Ridges, S.; Heaton, W.L.; Joshi, D.; Choi, H.; Eiring, A.; Batchelor, L.; Choudhry, P.; Manos, E.J.; Sofla, H.; Sanati, A. Zebrafish screen identifies novel compound with selective toxicity against leukemia. Blood J. Am. Soc. Hematol. 2012, 119, 5621–5631. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.; Pan, L.; Groen, R.W.; Baleydier, F.; Kentsis, A.; Marineau, J.; Grebliunaite, R.; Kozakewich, E.; Reed, C.; Pflumio, F. Phenothiazines induce PP2A-mediated apoptosis in T cell acute lymphoblastic leukemia. J. Clin. Investig. 2014, 124, 644–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Xiong, J.W. Molecular and developmental biology of the hemangioblast. Dev. Dyn. 2008, 237, 1218–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.Z.; Nasr, F.A.; Qamar, W.; Noman, O.M.; Khan, J.M.; Al-Mishari, A.A.; Alqahtani, A.S. Janerin Induces Cell Cycle Arrest at the G2/M Phase and Promotes Apoptosis Involving the MAPK Pathway in THP-1, Leukemic Cell Line. Molecules 2021, 26, 7555. [Google Scholar] [CrossRef] [PubMed]
- Isogai, S.; Horiguchi, M.; Weinstein, B.M. The vascular anatomy of the developing zebrafish: An atlas of embryonic and early larval development. Dev. Biol. 2001, 230, 278–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio); University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Schoofs, G.M.; Willhite, C.C. A probit analysis program for the personal computer. J. Appl. Toxicol. 1984, 4, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Strahle, U.; Scholz, S.; Geisler, R.; Greiner, P.; Hollert, H.; Rastegar, S.; Schumacher, A.; Selderslaghs, I.; Weiss, C.; Witters, H.; et al. Zebrafish embryos as an alternative to animal experiments—A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 2012, 33, 128–132. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No. | Cell Lines and IC50 (µM) | |
|---|---|---|
| Jurkat | THP-1 | |
| 4a | 6.1 ± 0.19 | 6.5 ± 0.18 |
| 4b | 17.4 ± 0.28 | 16.7 ± 0.61 |
| 4c | - | - |
| 4d | 4.4 ± 0.2 | 4.2 ± 0.2 |
| 4e | - | - |
| 4f | 12.55 ± 0.63 | 11.9 ± 0.3 |
| 4g | - | - |
| 4h | 13.4 ± 0.15 | 9.75 ± 0.35 |
| 4i | 33.8 ± 1.5 | 8.37 ± 0.1 |
| 4j | - | - |
| 4k | - | - |
| 4l | - | - |
| 4m | - | - |
| 4n | - | - |
| Doxorubicin | 1.3 ± 0.01 | 1.1 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqahtani, A.S.; Ghorab, M.M.; Nasr, F.A.; Ahmed, M.Z.; Al Mishari, A.A.; Attia, S.M.; Farooq Khan, M. Cytotoxicity of Newly Synthesized Quinazoline–Sulfonamide Derivatives in Human Leukemia Cell Lines and Their Effect on Hematopoiesis in Zebrafish Embryos. Int. J. Mol. Sci. 2022, 23, 4720. https://doi.org/10.3390/ijms23094720
Alqahtani AS, Ghorab MM, Nasr FA, Ahmed MZ, Al Mishari AA, Attia SM, Farooq Khan M. Cytotoxicity of Newly Synthesized Quinazoline–Sulfonamide Derivatives in Human Leukemia Cell Lines and Their Effect on Hematopoiesis in Zebrafish Embryos. International Journal of Molecular Sciences. 2022; 23(9):4720. https://doi.org/10.3390/ijms23094720
Chicago/Turabian StyleAlqahtani, Ali S., Mostafa M. Ghorab, Fahd A. Nasr, Mohammad Z. Ahmed, Abdullah A. Al Mishari, Sabry M. Attia, and Muhammad Farooq Khan. 2022. "Cytotoxicity of Newly Synthesized Quinazoline–Sulfonamide Derivatives in Human Leukemia Cell Lines and Their Effect on Hematopoiesis in Zebrafish Embryos" International Journal of Molecular Sciences 23, no. 9: 4720. https://doi.org/10.3390/ijms23094720