
Role of Polyamine-Induced Dimerization of Antizyme in Its Cellular Functions

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
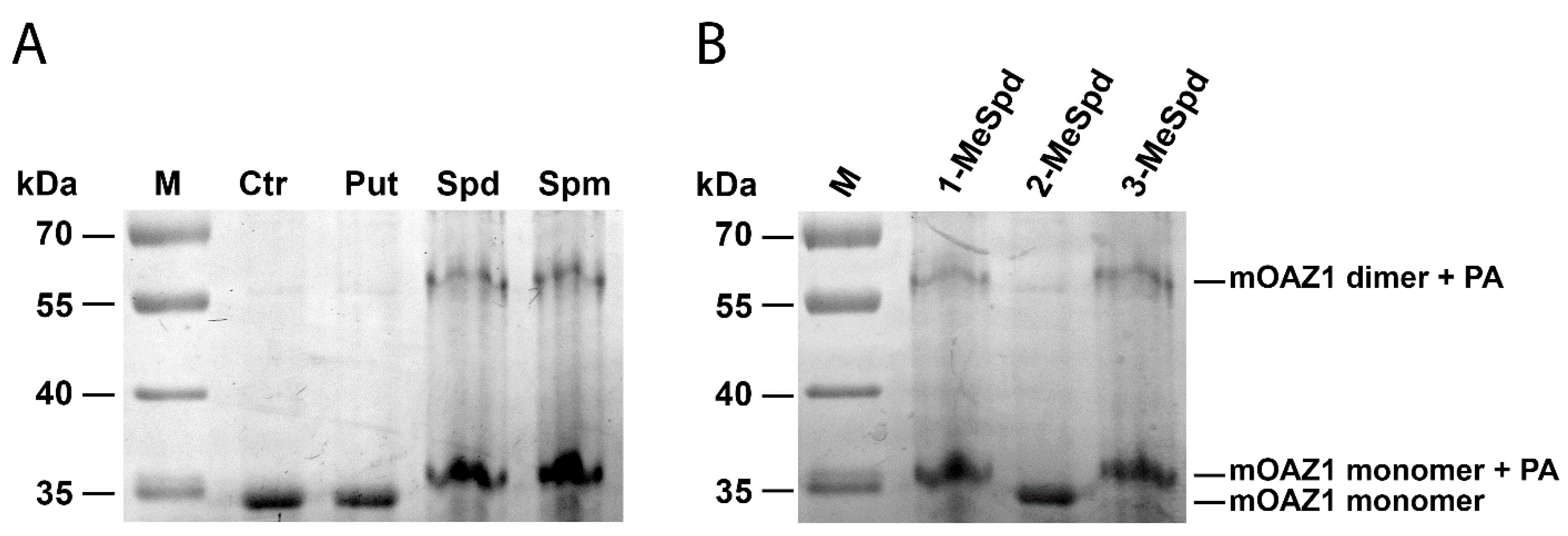
2.1. Polyamines Induce mOAZ1 Dimerization In Vitro
2.2. Modulation of (mOAZ1)2-Polyamine Complex Formation with MeSpds
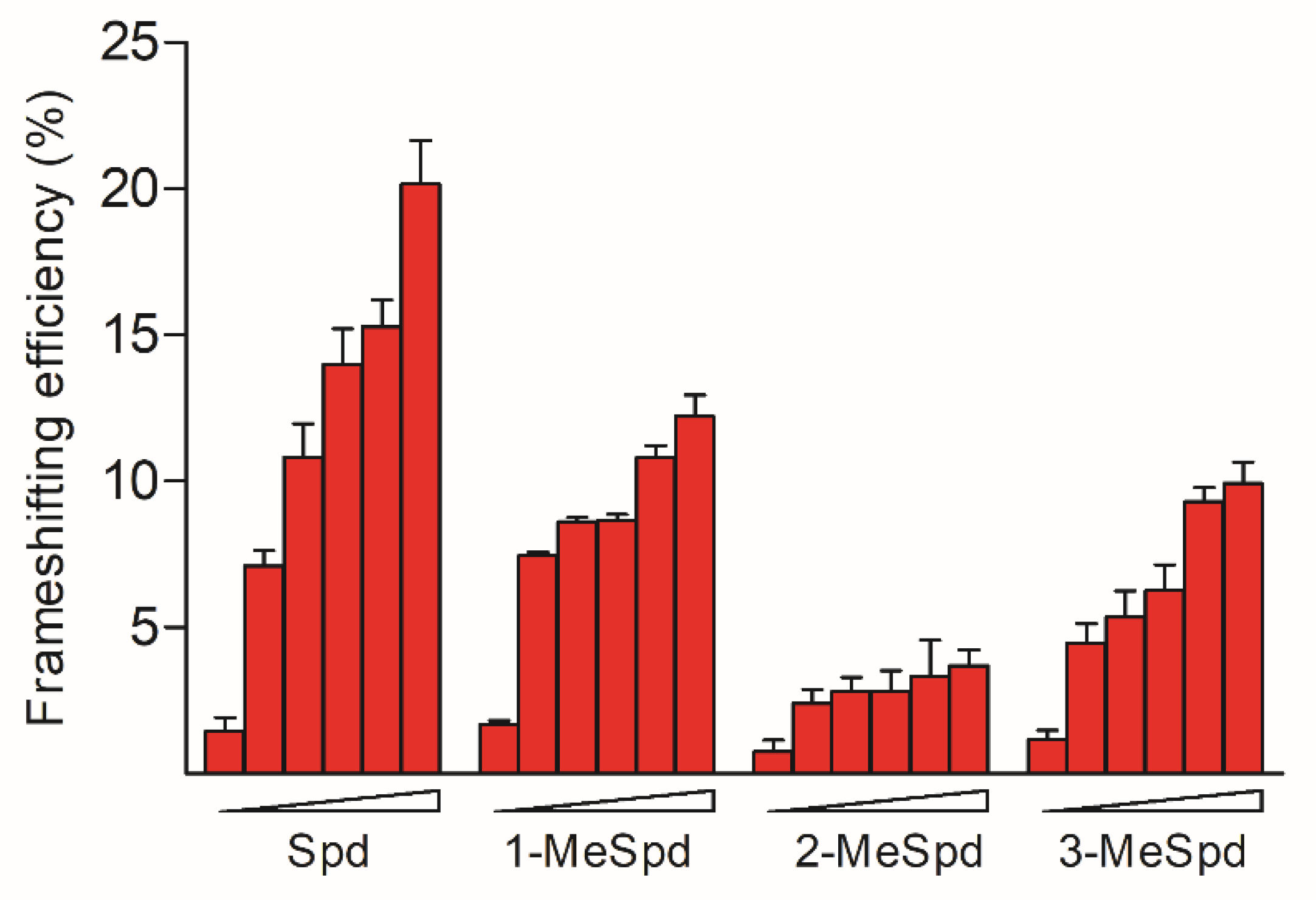
2.3. MeSpds Differently Induce +1 Frameshifting of OAZ1 mRNA
2.4. MeSpds Differently Stabilize the Stem–Loop Region of mOAZ1 mRNA
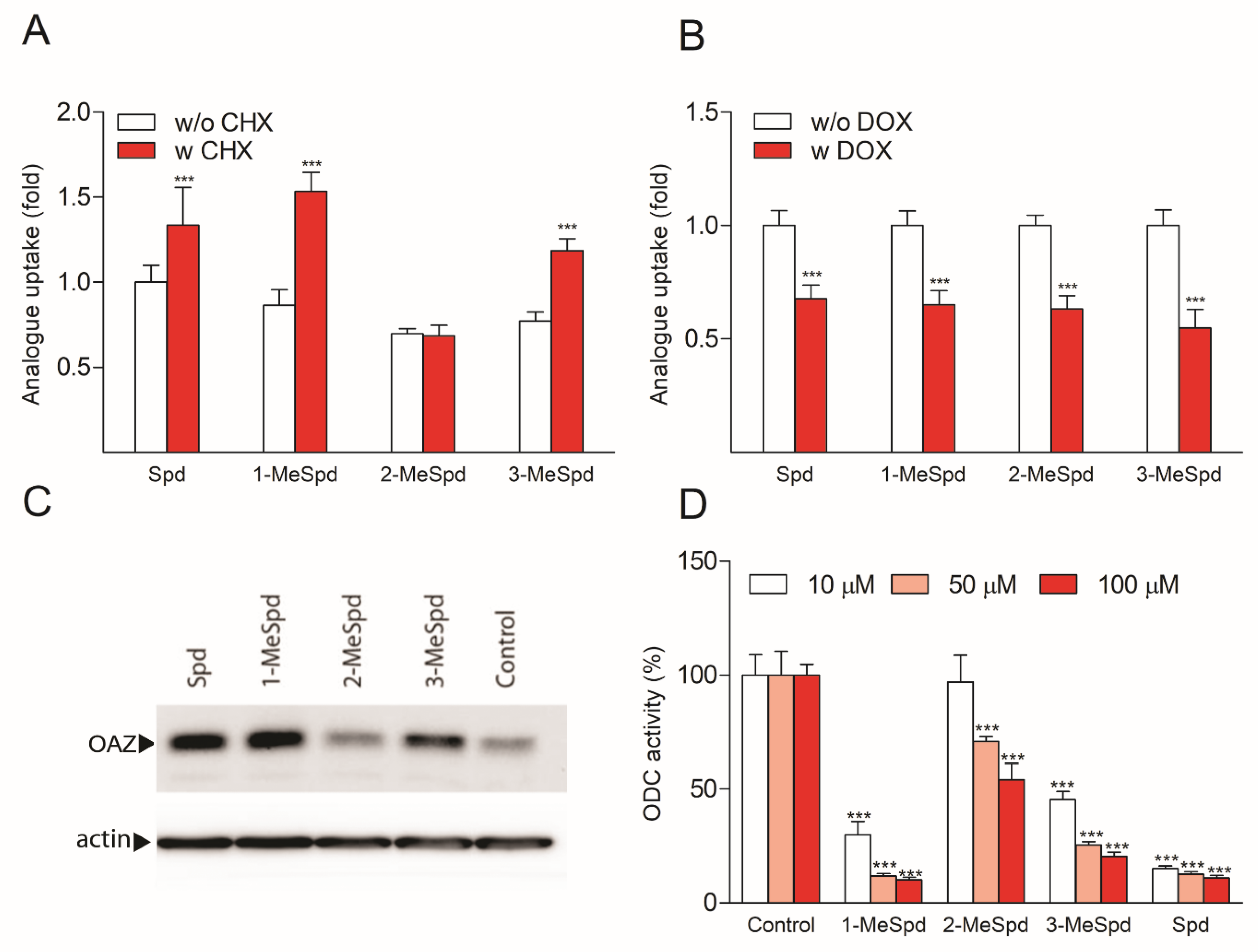
2.5. MeSpds Differently Affect OAZ1 Synthesis and ODC Activity in DU145 Cells
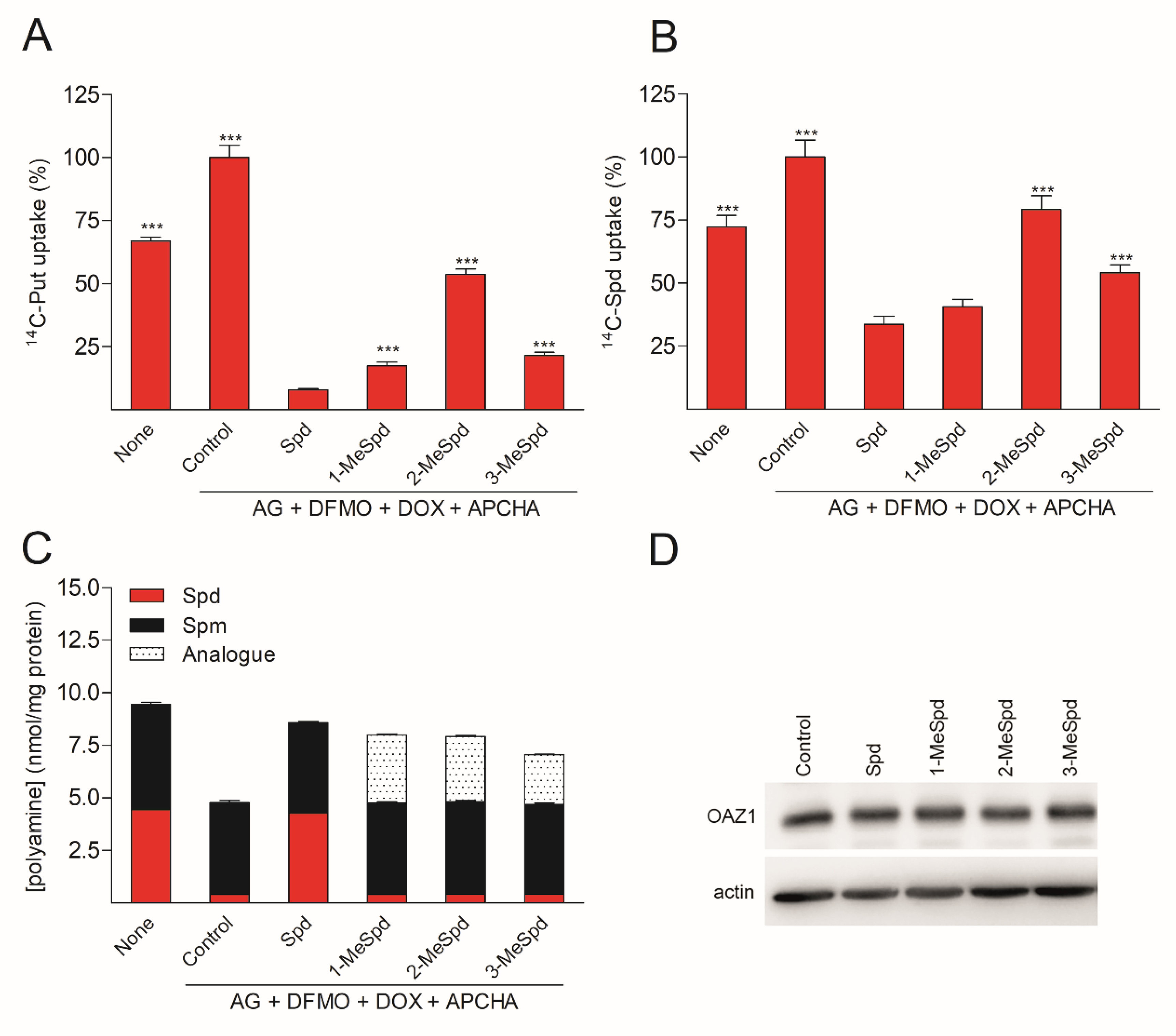
2.6. (OAZ1)2-Polyamine Complex Downregulates Polyamine Uptake in DU145 Cells
2.7. Polyamines Do Not Stabilize OAZ1 Protein in DU145 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plasmid Construction
4.3. The Expression and Purification of mOAZ1 Protein
4.4. Isothermal Titration Calorimetry (ITC)
4.5. Electrophoresis of mOAZ1-Polyamine Complex
4.6. Cell Culture
4.7. [14C]-Put and -Spd Uptake Experiments
4.8. OAZ1 Half-Life in DU145 Cells
4.9. In Vitro Effect of Spd and Its Analogues on ODC Activity
4.10. RNA Melting Point Experiments
4.11. Determination of the Efficiency of Ribosomal Frameshifting Using β-Galactosidase and Firefly Luciferase Reporter System
4.12. Analytical Methods
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, N.M.; Ju, S.; Verbitsky, M.; Ross, B.; Geddie, M.L.; Rockenstein, E.; Adame, A.; Muhammad, A.; Vonsattel, J.P.; Ringe, D.; et al. Polyamine pathway contributes to the pathogenesis of Parkinson disease. Proc. Natl. Acad. Sci. USA 2010, 107, 16970–16975. [Google Scholar] [CrossRef] [Green Version]
- Alhonen, L.; Parkkinen, J.J.; Keinänen, T.; Sinervirta, R.; Herzig, K.H.; Jänne, J. Activation of polyamine catabolism in transgenic rats induces acute pancreatitis. Proc. Natl. Acad. Sci. USA 2000, 97, 8290–8295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casero, R.A., Jr.; Murray Stewart, T.; Pegg, A.E. Polyamine metabolism and cancer: Treatments, challenges and opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Murray-Stewart, T.; Dunworth, M.; Foley, J.R.; Schwartz, C.E.; Casero, R.A., Jr. Polyamine Homeostasis in Snyder-Robinson Syndrome. Med. Sci. 2018, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Damiani, E.; Wallace, H.M. Polyamines and Cancer. Methods Mol. Biol. 2018, 1694, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Guerra, G.P.; Rubin, M.A.; Mello, C.F. Modulation of learning and memory by natural polyamines. Pharmacol. Res. 2016, 112, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yu, C.H.; Yu, H.; Borstnar, R.; Kamerlin, S.C.; Graslund, A.; Abrahams, J.P.; Warmlander, S.K. Cellular polyamines promote amyloid-beta (Aβ) peptide fibrillation and modulate the aggregation pathways. ACS Chem. Neurosci. 2013, 4, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Kahana, C. The antizyme family for regulating polyamines. J. Biol. Chem. 2018, 293, 18730–18735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, Y.; Matsufuji, S.; Kameji, T.; Hayashi, S.; Igarashi, K.; Tamura, T.; Tanaka, K.; Ichihara, A. Ornithine decarboxylase is degraded by the 26S proteasome without ubiquitination. Nature 1992, 360, 597–599. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; He, Y.; Kashiwagi, K.; Murakami, Y.; Hayashi, S.; Igarashi, K. Antizyme protects against abnormal accumulation and toxicity of polyamines in ornithine decarboxylase- overproducing cells. Proc. Natl. Acad. Sci. USA 1994, 91, 8930–8934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.K.; Gopalan, G. Antizyme1 mediates AURKAIP1-dependent degradation of Aurora-A. Oncogene 2007, 26, 6593–6603. [Google Scholar] [CrossRef] [Green Version]
- Gruendler, C.; Lin, Y.; Farley, J.; Wang, T. Proteasomal degradation of Smad1 induced by bone morphogenetic proteins. J. Biol. Chem. 2001, 276, 46533–46543. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Lee, C.Y.; Lin, C.L.; Chen, H.Y.; Liu, G.Y.; Hung, H.C. Multifaceted interactions and regulation between antizyme and its interacting proteins cyclin D1, ornithine decarboxylase and antizyme inhibitor. Oncotarget 2015, 6, 23917–23929. [Google Scholar] [CrossRef] [Green Version]
- Newman, R.M.; Mobascher, A.; Mangold, U.; Koike, C.; Diah, S.; Schmidt, M.; Finley, D.; Zetter, B.R. Antizyme targets cyclin D1 for degradation. A novel mechanism for cell growth repression. J. Biol. Chem. 2004, 279, 41504–41511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bercovich, Z.; Snapir, Z.; Keren-Paz, A.; Kahana, C. Antizyme affects cell proliferation and viability solely through regulating cellular polyamines. J. Biol. Chem. 2011, 286, 33778–33783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajima, A.; Murai, N.; Murakami, Y.; Iwamoto, T.; Migita, T.; Matsufuji, S. Polyamine regulating protein antizyme binds to ATP citrate lyase to accelerate acetyl-CoA production in cancer cells. Biochem. Biophys. Res. Commun. 2016, 471, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Matsufuji, S.; Matsufuji, T.; Miyazaki, Y.; Murakami, Y.; Atkins, J.F.; Gesteland, R.F.; Hayashi, S.-I. Autoregulatory frameshifting in decoding mammalian ornithine decarboxylase antizyme. Cell 1995, 80, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Rom, E.; Kahana, C. Polyamines regulate the expression of ornithine decarboxylase antizyme in vitro by inducing ribosomal frame-shifting. Proc. Natl. Acad. Sci. USA 1994, 91, 3959–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, I.P.; Atkins, J.F. Ribosomal frameshifting in decoding antizyme mRNAs from yeast and protists to humans: Close to 300 cases reveal remarkable diversity despite underlying conservation. Nucleic Acids Res. 2007, 35, 1842–1858. [Google Scholar] [CrossRef] [PubMed]
- Petros, L.M.; Howard, M.T.; Gesteland, R.F.; Atkins, J.F. Polyamine sensing during antizyme mRNA programmed frameshifting. Biochem. Biophys. Res. Commun. 2005, 338, 1478–1489. [Google Scholar] [CrossRef]
- Hayashi, S.; Murakami, Y.; Matsufuji, S. Ornithine decarboxylase antizyme: A novel type of regulatory protein. Trends Biochem. Sci. 1996, 21, 27–30. [Google Scholar] [CrossRef]
- Petros, L.M.; Graminski, G.F.; Robinson, S.; Burns, M.R.; Kisiel, N.; Gesteland, R.F.; Atkins, J.F.; Kramer, D.L.; Howard, M.T.; Weeks, R.S. Polyamine Analogs with Xylene Rings Induce Antizyme Frameshifting, Reduce ODC Activity, and Deplete Cellular Polyamines. J. Biochem. 2006, 140, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Keinanen, T.A.; Hyvonen, M.T.; Alhonen, L.; Vepsalainen, J.; Khomutov, A.R. Selective regulation of polyamine metabolism with methylated polyamine analogues. Amino Acids 2014, 46, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Hyvönen, M.T.; Keinänen, T.A.; Khomutov, M.; Simonian, A.; Weisell, J.; Kochetkov, S.N.; Vepsäläinen, J.; Alhonen, L.; Khomutov, A.R. The use of novel C-methylated spermidine derivatives to investigate the regulation of polyamine metabolism. J. Med. Chem. 2011, 54, 4611–4618. [Google Scholar] [CrossRef]
- Khomutov, M.; Hyvönen, M.T.; Simonian, A.; Formanovsky, A.A.; Mikhura, I.V.; Chizhov, A.O.; Kochetkov, S.N.; Alhonen, L.; Vepsäläinen, J.; Keinänen, T.A.; et al. Unforeseen Possibilities to Investigate the Regulation of Polyamine Metabolism Revealed by Novel C-Methylated Spermine Derivatives. J. Med. Chem. 2019, 62, 11335–11347. [Google Scholar] [CrossRef]
- Atkins, J.F.; Loughran, G.; Bhatt, P.R.; Firth, A.E.; Baranov, P.V. Ribosomal frameshifting and transcriptional slippage: From genetic steganography and cryptography to adventitious use. Nucleic Acids Res. 2016, 44, 7007–7078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rato, C.; Amirova, S.R.; Bates, D.G.; Stansfield, I.; Wallace, H.M. Translational recoding as a feedback controller: Systems approaches reveal polyamine-specific effects on the antizyme ribosomal frameshift. Nucleic Acids Res. 2011, 39, 4587–4597. [Google Scholar] [CrossRef] [Green Version]
- Palanimurugan, R.; Scheel, H.; Hofmann, K.; Dohmen, R.J. Polyamines regulate their synthesis by inducing expression and blocking degradation of ODC antizyme. EMBO J. 2004, 23, 4857–4867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef]
- Erales, J.; Coffino, P. Ubiquitin-independent proteasomal degradation. Biochim. Biophys. Acta 2014, 1843, 216–221. [Google Scholar] [CrossRef] [Green Version]
- Matsufuji, S.; Kanamoto, R.; Murakami, Y.; Hayashi, S. Monoclonal antibody studies on the properties and regulation of murine ornithine decarboxylase antizymes. J. Biochem. 1990, 107, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.-i.; Kusama-Eguchi, K.; Kobayashi, H.; Igarashi, K. Estimation of polyamine binding to macromolecules and ATP in bovine lymphocytes and rat liver. J. Biol. Chem. 1991, 266, 20803–20809. [Google Scholar] [CrossRef]
- Schuster, I.; Bernhardt, R. Interactions of natural polyamines with mammalian proteins. Biomol. Concepts 2011, 2, 79–94. [Google Scholar] [CrossRef]
- Savarin, P.; Barbet, A.; Delga, S.; Joshi, V.; Hamon, L.; Lefevre, J.; Nakib, S.; De Bandt, J.P.; Moinard, C.; Curmi, P.A.; et al. A central role for polyamines in microtubule assembly in cells. Biochem. J. 2010, 430, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Leal, O.; Barrero, C.A.; Clarkson, A.B.; Casero, R.A., Jr.; Merali, S. Polyamine-regulated translation of spermidine/spermine-N1-acetyltransferase. Mol. Cell. Biol. 2012, 32, 1453–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, D.W.; Carroll, D.; Martinez, N.; Hackert, M.L. Solution structure of a conserved domain of antizyme: A protein regulator of polyamines. Biochemistry 2005, 44, 11777–11785. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Chen, S.F.; Hsieh, J.Y.; Chou, F.; Wang, Y.H.; Lin, W.T.; Lee, P.Y.; Yu, Y.J.; Lin, L.Y.; Lin, T.S.; et al. Structural basis of antizyme-mediated regulation of polyamine homeostasis. Proc. Natl. Acad. Sci. USA 2015, 112, 11229–11234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurian, L.; Palanimurugan, R.; Godderz, D.; Dohmen, R.J. Polyamine sensing by nascent ornithine decarboxylase antizyme stimulates decoding of its mRNA. Nature 2011, 477, 490–494. [Google Scholar] [CrossRef]
- Romani, A.M. Cellular magnesium homeostasis. Arch. Biochem. Biophys. 2011, 512, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Cowan, J.A. Structural and catalytic chemistry of magnesium-dependent enzymes. Biometals 2002, 15, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Hyvönen, M.T.; Keinänen, T.A.; Khomutov, M.; Simonian, A.; Vepsäläinen, J.; Park, J.H.; Khomutov, A.R.; Alhonen, L.; Park, M.H. Effects of novel C-methylated spermidine analogs on cell growth via hypusination of eukaryotic translation initiation factor 5A. Amino Acids 2012, 42, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Vuohelainen, S.; Pirinen, E.; Cerrada-Gimenez, M.; Keinänen, T.A.; Uimari, A.; Pietilä, M.; Khomutov, A.R.; Jänne, J.; Alhonen, L. Spermidine is indispensable in differentiation of 3T3-L1 fibroblasts to adipocytes. J. Cell. Mol. Med. 2010, 14, 1683–1692. [Google Scholar] [CrossRef] [Green Version]
- Hyvönen, M.T.; Koponen, T.; Weisell, J.; Pietilä, M.; Khomutov, A.R.; Vepsäläinen, J.; Alhonen, L.; Keinänen, T.A. Spermidine promotes adipogenesis of 3T3-L1 cells by preventing interaction of ANP32 with HuR and PP2A. Biochem. J. 2013, 453, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Hobley, L.; Li, B.; Wood, J.L.; Kim, S.H.; Naidoo, J.; Ferreira, A.S.; Khomutov, M.; Khomutov, A.; Stanley-Wall, N.R.; Michael, A.J. Spermidine promotes Bacillus subtilis biofilm formation by activating expression of the matrix regulator slrR. J. Biol. Chem. 2017, 292, 12041–12053. [Google Scholar] [CrossRef] [Green Version]
- Räsänen, T.L.; Alhonen, L.; Sinervirta, R.; Keinänen, T.; Herzig, K.H.; Suppola, S.; Khomutov, A.R.; Vepsäläinen, J.; Jänne, J. A polyamine analogue prevents acute pancreatitis and restores early liver regeneration in transgenic rats with activated polyamine catabolism. J. Biol. Chem. 2002, 277, 39867–39872. [Google Scholar] [CrossRef] [Green Version]
- Hyvönen, M.T.; Sinervirta, R.; Grigorenko, N.; Khomutov, A.R.; Vepsäläinen, J.; Keinänen, T.A.; Alhonen, L. alpha-Methylspermidine protects against carbon tetrachloride-induced hepatic and pancreatic damage. Amino Acids 2010, 38, 575–581. [Google Scholar] [CrossRef]
- Hyvönen, M.T.; Sinervirta, R.; Keinänen, T.A.; Fashe, T.; Grigorenko, N.; Khomutov, A.R.; Vepsäläinen, J.; Alhonen, L. Acute pancreatitis induced by activated polyamine catabolism is associated with coagulopathy: Effects of alpha-methylated polyamine analogs on hemostasis. Pancreatology 2010, 10, 208–221. [Google Scholar] [CrossRef]
- Fashe, T.M.; Keinänen, T.A.; Grigorenko, N.A.; Khomutov, A.R.; Jänne, J.; Alhonen, L.; Pietilä, M. Cutaneous application of alpha-methylspermidine activates the growth of resting hair follicles in mice. Amino Acids 2010, 38, 583–590. [Google Scholar] [CrossRef]
- Marzani, B.; Pinto, D.; Sorbellini, E.; Rinaldi, F. New multi-targeting strategy in hair growth promotion: In vitro and in vivo studies. G Ital. Dermatol. Venereol. 2018, 153, 338–343. [Google Scholar] [CrossRef]
- Beenukumar, R.R.; Godderz, D.; Palanimurugan, R.; Dohmen, R.J. Polyamines directly promote antizyme-mediated degradation of ornithine decarboxylase by the proteasome. Microb. Cell 2015, 2, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, J.L.; Leyser, A.; Holtorff, M.S.; Bates, J.S.; Frydman, B.; Valasinas, A.L.; Reddy, V.K.; Marton, L.J. Antizyme induction by polyamine analogues as a factor of cell growth inhibition. Biochem. J. 2002, 366, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Coffino, P. Regulation of cellular polyamines by antizyme. Nat. Rev. Mol. Cell Biol. 2001, 2, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, A.J.; Cerrada-Gimenez, M.; Grigorenko, N.A.; Khomutov, A.R.; Vepsäläinen, J.J.; Sinervirta, R.M.; Keinänen, T.A.; Alhonen, L.I.; Jänne, J. Alpha-methyl polyamines: Efficient synthesis and tolerance studies in vivo and in vitro. First evidence for dormant stereospecificity of polyamine oxidase. J. Med. Chem. 2006, 49, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Rehse, K.; Puchert, E.; Leissring, S. Alkyl- und Arylalkyderivate von Putrescin, Spermidin und Spermin. Arch. Pharm. 1990, 323, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Shirahata, A.; Morohohi, T.; Fukai, M.; Akatsu, S.; Samajima, K. Putrescine or spermidine binding site of aminopropyltransferases and competitive inhibitors. Biochem. Pharmacol. 1991, 41, 205–212. [Google Scholar] [CrossRef]
- Kankare, K.; Uusi-Oukari, M.; Janne, O.A. Structure, organization and expression of the mouse ornithine decarboxylase antizyme gene. Biochem. J. 1997, 324, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Korovina, A.N.; Tunitskaya, V.L.; Kostyuk, D.A.; Rechinsky, V.O.; Kukhanova, M.K.; Kochetkov, S.N. Development of the system ensuring a high-level expression of hepatitis C virus nonstructural NS5B and NS5A proteins. Protein Expr. Purif. 2006, 48, 14–23. [Google Scholar] [CrossRef]
- Petrushanko, I.Y.; Mitkevich, V.A.; Anashkina, A.A.; Klimanova, E.A.; Dergousova, E.A.; Lopina, O.D.; Makarov, A.A. Critical role of gamma-phosphate in structural transition of Na,K-ATPase upon ATP binding. Sci. Rep. 2014, 4, 5165. [Google Scholar] [CrossRef] [Green Version]
- Weisell, J.; Hyvönen, M.T.; Häkkinen, M.R.; Grigorenko, N.A.; Pietilä, M.; Lampinen, A.; Kochetkov, S.N.; Alhonen, L.; Vepsäläinen, J.; Keinänen, T.A.; et al. Synthesis and biological characterization of novel charge-deficient spermine analogues. J. Med. Chem. 2010, 53, 5738–5748. [Google Scholar] [CrossRef]
- Hou, M.H.; Lin, S.B.; Yuann, J.M.; Lin, W.C.; Wang, A.H.; Kan Ls, L. Effects of polyamines on the thermal stability and formation kinetics of DNA duplexes with abnormal structure. Nucleic Acids Res. 2001, 29, 5121–5128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osipov, S.T.; Preobrazhenskaya, O.V.; Karpov, V. A rapid method for liquid β-galactosidase reporter assay in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2011, 27, 1255–1259. [Google Scholar] [CrossRef]
- Hyvönen, T.; Keinänen, T.A.; Khomutov, A.R.; Khomutov, R.M.; Eloranta, T.O. Monitoring of the uptake and metabolism of aminooxy analogues of polyamines in cultured cells by high-performance liquid chromatography. J. Chromatogr. 1992, 574, 17–21. [Google Scholar] [CrossRef]
- Jänne, J.; Williams-Ashman, H.G. On the purification of L-ornithine decarboxylase from rat prostate and effects of thiol compounds on the enzyme. J. Biol. Chem. 1971, 246, 1725–1732. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyamine | Stoichiometry mOAZ1/Polyamine | Ka b, M−1 | ΔH d, kcal/mole | TΔS e, kcal/mole | Kd c, µM |
|---|---|---|---|---|---|
 Spd | 2:1 | 2.8 × 105 | −3.3 | 4.2 | 3.6 |
 Spm | 2:1 | 9.2 × 105 | −3.7 | 4.5 | 1.1 |
 DENSpm | 2:1 | 7.5 × 105 | −5.1 | 3.0 | 1.3 |
 Put | No binding f | ||||
 1-MeSpd | 2:1 | 5.4 × 105 | −2.2 | 5.8 | 1.9 |
 2-MeSpd | No binding f | ||||
 3-MeSpd | 2:1 | 4.0 × 105 | −1.2 | 6.6 | 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyvönen, M.T.; Smirnova, O.A.; Mitkevich, V.A.; Tunitskaya, V.L.; Khomutov, M.; Karpov, D.S.; Korolev, S.P.; Häkkinen, M.R.; Pietilä, M.; Gottikh, M.B.; et al. Role of Polyamine-Induced Dimerization of Antizyme in Its Cellular Functions. Int. J. Mol. Sci. 2022, 23, 4614. https://doi.org/10.3390/ijms23094614
Hyvönen MT, Smirnova OA, Mitkevich VA, Tunitskaya VL, Khomutov M, Karpov DS, Korolev SP, Häkkinen MR, Pietilä M, Gottikh MB, et al. Role of Polyamine-Induced Dimerization of Antizyme in Its Cellular Functions. International Journal of Molecular Sciences. 2022; 23(9):4614. https://doi.org/10.3390/ijms23094614
Chicago/Turabian StyleHyvönen, Mervi T., Olga A. Smirnova, Vladimir A. Mitkevich, Vera L. Tunitskaya, Maxim Khomutov, Dmitry S. Karpov, Sergey P. Korolev, Merja R. Häkkinen, Marko Pietilä, Marina B. Gottikh, and et al. 2022. "Role of Polyamine-Induced Dimerization of Antizyme in Its Cellular Functions" International Journal of Molecular Sciences 23, no. 9: 4614. https://doi.org/10.3390/ijms23094614