
TaNBR1, a Novel Wheat NBR1-like Domain Gene Negatively Regulates Drought Stress Tolerance in Transgenic Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Molecular Characterization of TaNBR1
2.2. TaNBR1 Expression Profiles
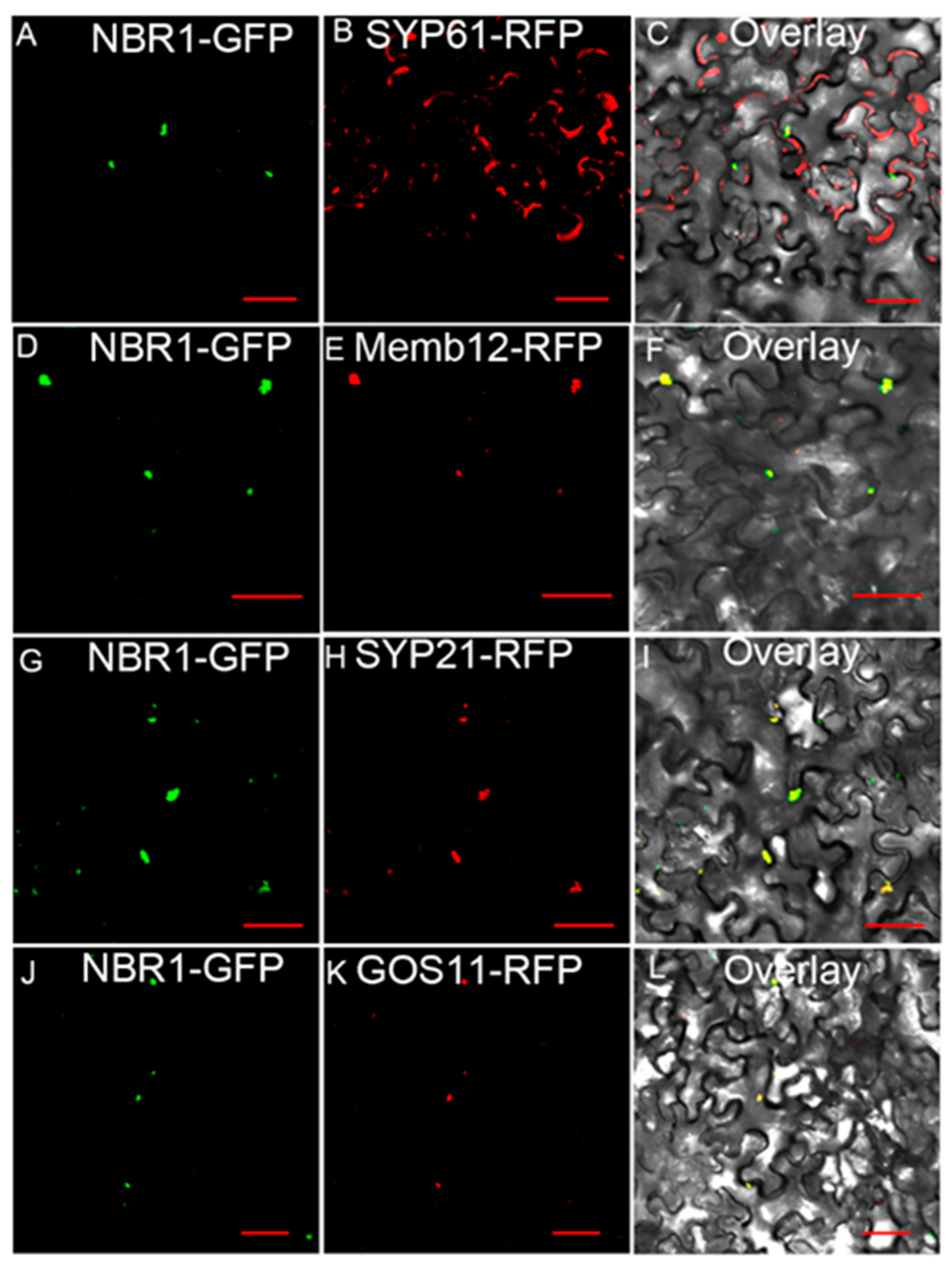
2.3. TaNBR1 Localizes in the Golgi Apparatus and Autophagosomes
2.4. The Characteristics of TaNBR1 Overexpressing Transgenic Arabidopsis
2.5. The Effects on Seed Germination and Seedling Growth in TaNBR1 Overexpression Transgenic Plants under Drought Stress
2.6. The Overexpression of TaNBR1 in Plants Sensitive to ABA
2.7. The Effects of TaNBR1 on the Expression of Stress-Responsive Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. The Isolation of TaNBR1 and Sequence Analysis
4.3. RNA Isolation and Quantitative real-Time PCR (qRT-PCR)
4.4. The Subcellular Localization of TaNBR1t
4.5. The Genetic Trnasformation of Arabidopsis thaliana
4.6. The Physiological Characteristics of the Transgenic Lines
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought re-sponses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [PubMed]
- Jiang, Y.; Liang, G.; Yang, S.; Yu, D. Arabidopsis WRKY57 functions as a node of convergence for jasmonic acid and auxinmediated signaling in jasmonic acid-induced leaf senescence. Plant Cell 2014, 26, 230–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.W.; Youn, J.H.; Park, T.K.; Kim, E.J.; Park, C.H.; Wang, Z.Y.; Kim, S.K.; Kim, T.W. OST1 activation by the brassinosteroid-regulated kinase CDG1-LIKE1 in stomatal closure. Plant Cell 2018, 30, 1848–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Jing, R.; Mao, X.; Jia, X.; Chang, X. A wheat (Triticum aestivum) protein phosphatase 2A catalytic subunit gene provides enhanced drought tolerance in tobacco. Ann. Bot. 2007, 99, 439–450. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Li, L.; Wang, P.; Li, D.; Zhuge, Q. Overexpression of PtHMGR enhances drought and salt tolerance of poplar. Ann. Bot. 2020, 125, 785–803. [Google Scholar] [CrossRef]
- Begcy, K.; Mariano, E.D.; Lembke, C.G.; Zingaretti, S.M.; Souza, G.M.; Araujo, P.; Menossi, M. Overexpression of an evolutionarily conserved drought-responsive sugarcane gene enhances salinity and drought resilience. Ann. Bot. 2019, 124, 691–700. [Google Scholar] [CrossRef]
- Zheng, Y.; Huang, Y.; Xian, W.; Wang, J.; Liao, H. Identification and expression analysis of the Glycine max CYP707A gene family in response to drought and salt stresses. Ann. Bot. 2012, 110, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Klionsky, D.J. Autophagosome formation, core machinery and adaptations. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef]
- Yoshimori, T. Autophagy: A regulated bulk degradation process inside cells. Biochem. Bioph. Res. Commun. 2004, 313, 453–458. [Google Scholar] [CrossRef]
- Bjorkoy, G.; Lamark, T.; Brech, A.; Outzen, H.; Perander, M.; Overvatn, A.; Stenmark, H.; Johansen, T. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J. Cell Biol. 2005, 171, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Kirkin, V.; Lamark, T.; Sou, Y.S.; Bjorkoy, G.; Nunn, J.L.; Bruun, J.A.; Shvets, E.; McEwan, D.G.; Clausen, T.H.; Wild, P.; et al. A role for NBR1 in autophagosomal degradation of ubiquitinated substrates. Mol. Cell. 2009, 33, 505–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurston, T.L.; Ryzhakov, G.; Bloor, S.; von Muhlinen, N.; Randow, F. The TBK1 adaptor and autophagy receptor NDP52 restricts the proliferation of ubiquitin-coated bacteria. Nat. Immunol. 2009, 10, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.L.; Li, S.; Zhao, Y.; Ma, X.F.; Zhang, K.; He, X.L.; Wang, Z. Interaction domains of p62: A bridge between p62 and selective autophagy. DNA Cell Biol. 2013, 32, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Shin, J. P62 and the sequestosome, a novel mechanism for protein metabolism. Arch. Pharm. Res. 1998, 21, 629–633. [Google Scholar] [CrossRef]
- Myeku, N.; Figueiredo-Pereira, M.E. Dynamics of the degradation of ubiquitinated proteins by proteasomes and au-tophagy, association with sequestosome 1/p62. J. Biol. Chem. 2011, 286, 22426–22440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkin, V.; McEwan, D.G.; Novak, I.; Dikic, I. A role for ubiquitin in selective autophagy. Mol. Cell 2009, 34, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Moscat, J.; Diaz-Meco, M.T.; Wooten, M.W. Signal integration and diversification through the p62 scaffold protein. Trends Biochem. Sci. 2007, 32, 95–100. [Google Scholar] [CrossRef]
- Lange, S.; Xiang, F.; Yakovenko, A.; Vihola, A.; Hackman, P.; Rostkova, E.; Kristensen, J.; Brandmeier, B.; Franzen, G.; Hedberg, B.; et al. The kinase domain of titin controls muscle gene expression and protein turnover. Science 2005, 308, 1599–1603. [Google Scholar] [CrossRef] [Green Version]
- Hafren, A.; Macia, J.L.; Love, A.J.; Milner, J.J.; Drucker, M.; Hofius, D. Selective autophagy limits cauliflower mosaic virus infection by NBR1-mediated targeting of viral capsid protein and particles. Proc. Natl. Acad. Sci. USA 2017, 114, E2026–E2035. [Google Scholar] [CrossRef] [Green Version]
- Dagdas, Y.F.; Pandey, P.; Tumtas, Y.; Sanguankiattichai, N.; Belhaj, K.; Duggan, C.; Leary, A.Y.; Segretin, M.E.; Contreras, M.P.; Savage, Z.; et al. Host autophagy machinery is diverted to the pathogen interface to mediate focal defense responses against the Irish potato famine pathogen. eLife 2018, 7, e37476. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Cheng, Y.; Chi, Y.J.; Fan, B.; Yu, J.Q.; Chen, Z. NBR1-mediated selective autophagy targets insoluble ubiquitinated protein aggregates in plant stress responses. PLoS Genet. 2013, 9, e1003196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.Y.; Zhou, J.; Guo, R.F.; Lin, Y.S.; Kung, C.H.; Hu, S.; Ng, W.Y.; Zhuang, X.H.; Jiang, L.W. AtNBR1 is a selective autophagic receptor for AtExo70E2 in Arabidopsis. Plant Physiol. 2020, 184, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Ustun, S.; Hafren, A.; Liu, Q.; Marshall, R.S.; Minina, E.A.; Bozhkov, P.V.; Vierstra, R.D.; Hofius, D. Bacteria exploit autophagy for proteasome degradation and enhanced virulence in plants. Plant Cell 2018, 30, 668–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafren, A.; Ustun, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip mosaic virus counteracts selective autophagy of the viral silencing suppressor HCpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Lee, H.N.; Marshall, R.S.; Lomax, A.W.; Yoon, M.J.; Kim, J.; Kim, J.H.; Vierstra, R.D.; Chung, T. Arabidopsis cargo receptor NBR1 mediates selective autophagy of defective proteins. J. Exp. Bot. 2020, 71, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Dagdas, Y.F.; Belhaj, K.; Maqbool, A.; Chaparro-Garcia, A.; Pandey, P.; Petre, B.; Tabassum, N.; Cruz-Mireles, N.; Hughes, R.K.; Sklenar, J.; et al. An effector of the Irish potato famine pathogen antagonizes a host autophagy cargo receptor. eLife 2016, 5, e10856. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Gorka, M.; Schulz, K.; Masclaux-Daubresse, C.; Sampathkumar, A.; Skirycz, A.; Vierstra, R.D.; Balazadeh, S. Selective autophagy regulates heat stress memory in Arabidopsis by NBR1-mediated targeting of HSP90 and ROF1. Autophagy 2021, 17, 2184–2199. [Google Scholar] [CrossRef]
- Lv, Q.; Zhang, L.; Zan, T.; Li, L.; Li, X. Wheat RING E3 ubiquitin ligase TaDIS1 degrade TaSTP via the 26S proteasome pathway. Plant Sci. 2020, 296, 110494. [Google Scholar] [CrossRef]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar]
- Liu, Y.M.; Xiong, Y.; Bassham, D.C. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy. 2009, 5, 954–963. [Google Scholar] [CrossRef] [Green Version]
- Slavikova, S.; Ufaz, S.; Avin-Wittenberg, T.; Levanony, H.; Galili, G. An autophagy-associated Atg8 protein is involved in the responses of Arabidopsis seedlings to hormonal controls and abiotic stresses. J. Exp. Bot. 2008, 59, 4029–4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, T.M.; Brennan, B.; Yang, M.R.; Chen, J.N.; Zhang, M.C.; Li, Z.H.; Wang, X.L.; Bassham, D.C.; Walley, J.; Yin, Y.H. Selective Autophagy of BES1 Mediated by DSK2 Balances Plant Growth and Survival. Dev. Cell 2017, 41, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Qin, H.; Zhou, S.R.; Wei, P.C.; Zhang, H.W.; Zhou, Y.; Miao, Y.C.; Huang, R.F. The Ubiquitin-Binding Protein OsDSK2a Mediates Seedling Growth and Salt Responses by Regulating Gibberellin Metabolism in Rice. Plant Cell 2020, 32, 414–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, Z.; Wang, X.T.; Li, X.F.; Zhang, Z.C.; Fan, B.F.; Zhu, C.; Chen, Z.X. Dicot-specific ATG8-interacting ATI3 proteins interact with conserved UBAC2 proteins and play critical roles in plant stress responses. Autophagy 2018, 14, 487–504. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, J.; Yu, J.Q.; Chen, Z. Role and regulation of autophagy in heat stress responses of tomato plants. Front. Plant Sci. 2014, 5, 174. [Google Scholar] [CrossRef] [Green Version]
- Werner, A.; Herzog, B.; Voigt, O.; Valerius, O.; Braus, G.H.; Poggeler, S. NBR1 is involved in selective pexophagy in filamentous ascomycetes and can be functionally replaced by a tagged version of its human homolog. Autophagy 2019, 15, 78–97. [Google Scholar] [CrossRef] [Green Version]
- Su, W.L.; Bao, Y.; Lu, Y.Y.; He, F.; Wang, S.; Wang, D.L.; Yu, X.Q.; Yin, W.L.; Xia, X.L.; Liu, C. Poplar autophagy receptor nbr1 enhances salt stress tolerance by regulating selective autophagy and antioxidant system. Front. Plant Sci. 2021, 11, 568411. [Google Scholar] [CrossRef]
- Mao, H.D.; Li, S.M.; Wang, Z.X.; Cheng, X.X.; Li, F.F.; Mei, F.M.; Chen, N.; Kang, Z.S. Regulatory changes in TaSNAC8-6A are associated with drought tolerance in wheat seedlings. Plant Biotechnol. J. 2020, 18, 1078–1092. [Google Scholar] [CrossRef] [Green Version]
- Asaoka, R.; Uemura, T.; Ito, J.; Fujimoto, M.; Ito, E.; Ueda, T.; Nakano, A. Arabidopsis RABA1 GTPases are involved in transport between the trans-Golgi network and the plasma membrane, and are required for salinity stress tolerance. Plant J. 2013, 73, 240–249. [Google Scholar] [CrossRef]
- Yun, H.R.; Rim, Y.G.; Heo, J.B. Rice small GTPase Rab11 is required for intracellular trafficking from the trans-Golgi-network to the plasma membrane and/or prevacuolar compartments. Appl. Biol. Chem. 2016, 59, 163–171. [Google Scholar] [CrossRef]
- Fang, Y.J.; Xiong, L.Z. General mechanisms of drought response and their application in drought resistance im-provement in plants. Cell Mol. Life. Sci. 2015, 72, 673–689. [Google Scholar]
- He, Z.; Wu, J.; Sun, X.; Dai, M. The maize clade a PP2C phosphatases play critical roles in multiple abiotic stress re-sponses. Int. J. Mol. Sci. 2019, 20, 3573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, Y.; Yin, Z.; Jiang, J.; Zhang, M.; Guo, X.; Ye, Z.; Zhao, Y.; Xiong, H.; Zhang, Z.; et al. OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2O2 signalling in rice. Plant Biotechnol. J. 2017, 15, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.; Dubey, R.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 9, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Li, L.; Zhou, K.H.; Zhang, Y.H.; Han, X.; Din, Y.P.; Ge, X.Y.; Qin, W.Q.; Wang, P.; Li, F.G.; et al. GhWRKY6 acts as a negative regulator in both transgenic Arabidopsis and cotton during drought and salt stress. Front. Genet. 2019, 10, 392. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, L.; Zhang, L.; Lv, Q.; Zhao, Y.; Li, X. Isolation and identification of wheat gene TaDIS1 encoding a RING finger domain protein, which negatively regulates drought stress tolerance in transgenic Arabidopsis. Plant Sci. 2018, 275, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.X.; Li, Y.; Li, D.D.; Xu, W.L.; Zheng, Y.; Li, X.B. Arabidopsis drought-induced protein Di19-3 participates in plant response to drought and high salinity stresses. Plant Mol. Biol. 2014, 86, 609–625. [Google Scholar] [CrossRef]
- Ma, Q.; Xia, Z.; Cai, Z.; Li, L.; Cheng, Y.; Liu, J.; Nian, H. GmWRKY16 enhances drought and salt tolerance through an ABA-mediated pathway in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1979. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant. 2016, 9, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dor-mancy and germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar]
- Hirayama, T.; Shinozaki, K. Perception and transduction of abscisic acid signals: Keys to the function of the versatile plant hormone ABA. Trends Plant Sci. 2007, 12, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Tarnowski, L.; Rodriguez, M.C.; Brzywczy, J.; Piecho-Kabacik, M.; Krckova, Z.; Martinec, J.; Wawrzynska, A.; Sirko, A. A selective autophagy cargo receptor NBR1 modulates abscisic acid signalling in Arabidopsis thaliana. Sci. Rep. 2020, 10, 7778. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Zhang, S.C.; Zhang, N.; Zhang, W.Y.; Li, M.J.; Liu, B.H.; Shi, Z.L. MYB-CC transcription factor, TaMYBsm3, cloned from wheat is involved in drought tolerance. BMC Plant Biol. 2019, 19, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhao, H.X.; Wang, X.L.; Kang, J.Y.; Lv, B.B.; Dong, Q.X.; Li, C.L.; Chen, H.; Wu, Q. Tartary buckwheat transcription factor FtbZIP5, regulated by FtSnRK2.6, can improve salt/drought resistance in transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svenning, S.; Lamark, T.; Krause, K.; Johansen, T. Plant NBR1 is a selective autophagy substrate and a functional hybrid of the mammalian autophagic adapters NBR1 and p62/SQSTM1. Autophagy 2011, 7, 993–1010. [Google Scholar] [CrossRef]
- Murray, M.; Thompson, W. Rapid isolation of high molecular weight plant ONA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Panda, B.B.; Badoghar, A.K.; Sekhar, S.; Shaw, B.P.; Mohapatra, P.K. 1-MCP treatment enhanced expression of genes controlling endosperm cell division and starch biosynthesis for improvement of grain filling in a dense-panicle rice cultivar. Plant Sci. 2016, 246, 11–25. [Google Scholar] [CrossRef]
- Che, R.; Tong, H.; Shi, B.; Liu, Y.; Fang, S.; Liu, D.; Xiao, Y.; Hu, B.; Liu, L.; Wang, H.; et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants. 2015, 2, 15195. [Google Scholar] [CrossRef]
- Moreau, P.; Brandizzi, F.; Hanton, S.; Chatre, L.; Melser, S.; Hawes, C.; Satiat-Jeunemaitre, B. The plant ER-Golgi in-terface, a highly structured and dynamic membrane complex. J. Exp. Bot. 2007, 58, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.R.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef]
- Tian, M.; Lou, L.; Liu, L.; Yu, F.; Zhao, Q.; Zhang, H.; Wu, Y.; Tang, S.; Xia, R.; Zhu, B.; et al. The RING finger E3 ligase STRF1 is involved in membrane trafficking and modulates salt-stress response in Arabidopsis thaliana. Plant J. 2015, 82, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Ferroni, L.; Zivcak, M.; Sytar, O.; Kovar, M.; Watanabe, N.; Pancaldi, S.; Baldisserotto, C.; Brestic, M. Chloro-phyll-depleted wheat mutants are disturbed in photosynthetic electron flow regulation but can retain an acclimation ability to a fluctuating light regime. Environ. Exp. Bot. 2020, 178, 104156. [Google Scholar] [CrossRef]
- Lou, D.; Wang, H.; Yu, D. The sucrose non-fermenting-1-related protein kinases SAPK1 and SAPK2 function collabo-ratively as positive regulators of salt stress tolerance in rice. BMC Plant Biol. 2018, 18, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Lv, Q.; Yang, W.; Yang, H.; Chen, Q.; Wang, B.; Lei, Y.; Xie, Y. TaNBR1, a Novel Wheat NBR1-like Domain Gene Negatively Regulates Drought Stress Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 4519. https://doi.org/10.3390/ijms23094519
Chen L, Lv Q, Yang W, Yang H, Chen Q, Wang B, Lei Y, Xie Y. TaNBR1, a Novel Wheat NBR1-like Domain Gene Negatively Regulates Drought Stress Tolerance in Transgenic Arabidopsis. International Journal of Molecular Sciences. 2022; 23(9):4519. https://doi.org/10.3390/ijms23094519
Chicago/Turabian StyleChen, Liuping, Qian Lv, Weibing Yang, Hui Yang, Qiaoyan Chen, Bingxin Wang, Yanhong Lei, and Yanzhou Xie. 2022. "TaNBR1, a Novel Wheat NBR1-like Domain Gene Negatively Regulates Drought Stress Tolerance in Transgenic Arabidopsis" International Journal of Molecular Sciences 23, no. 9: 4519. https://doi.org/10.3390/ijms23094519