
Small RNAs beyond Model Organisms: Have We Only Scratched the Surface?



Abstract
:1. Introduction
2. Prevalence of sRNAs in Bacteria
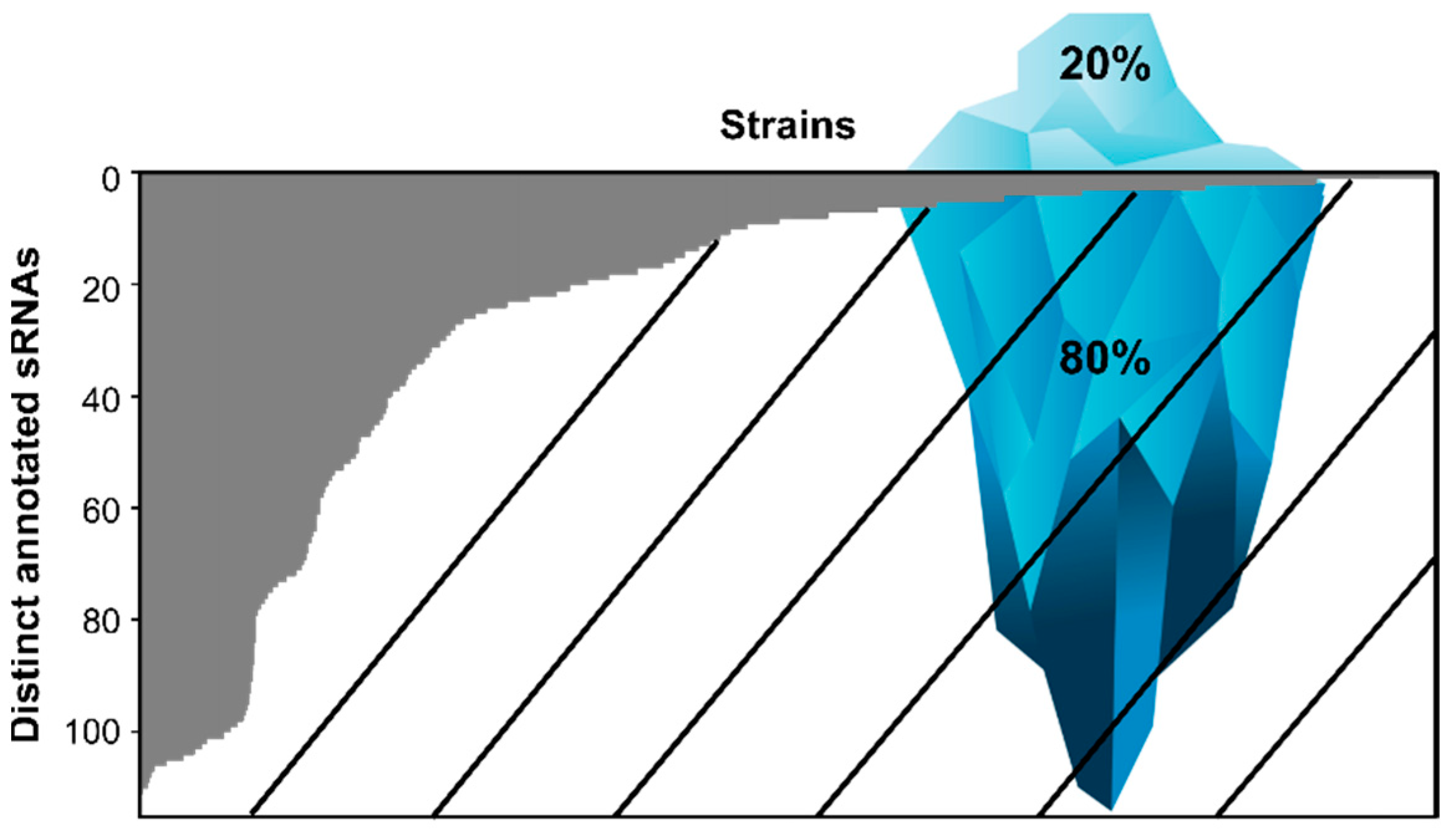
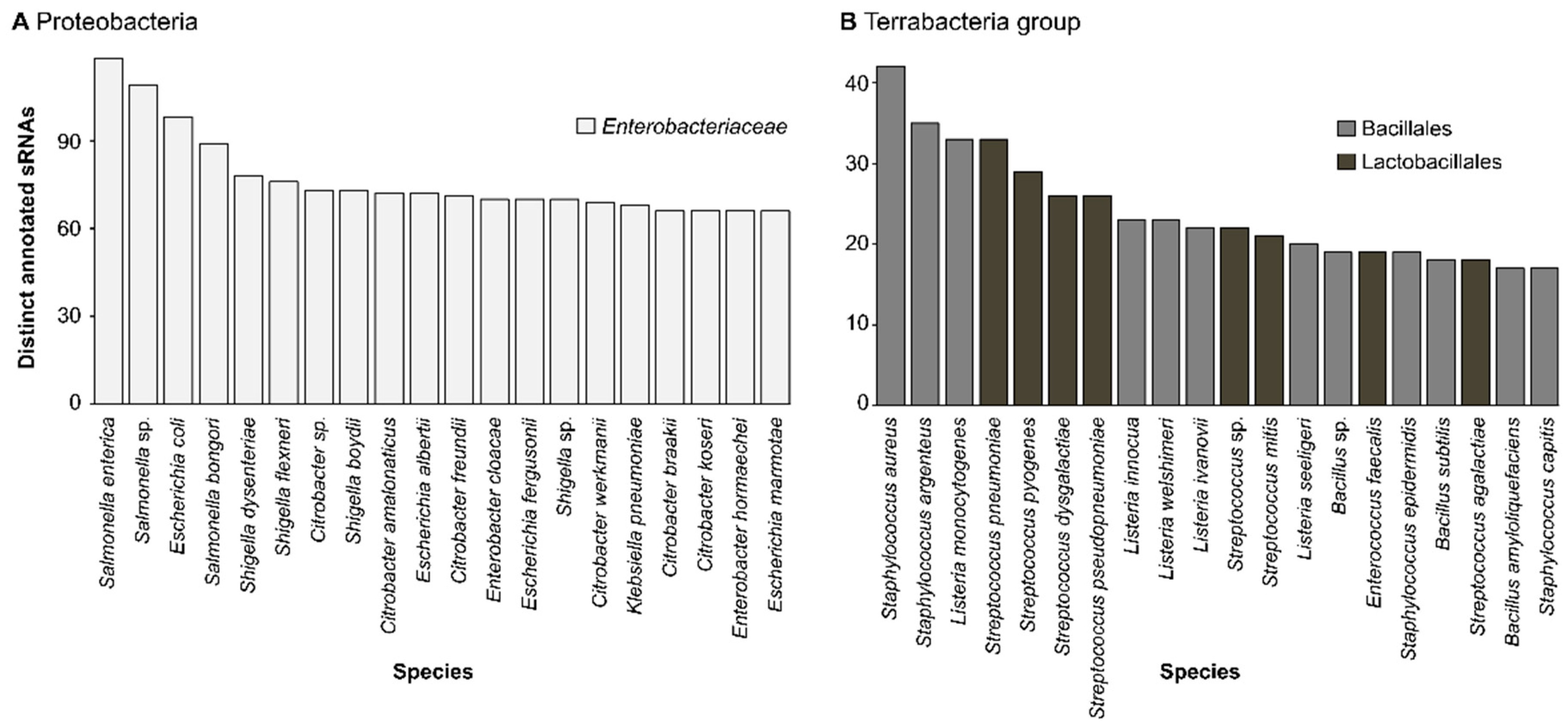
2.1. Species Encoding for sRNAs
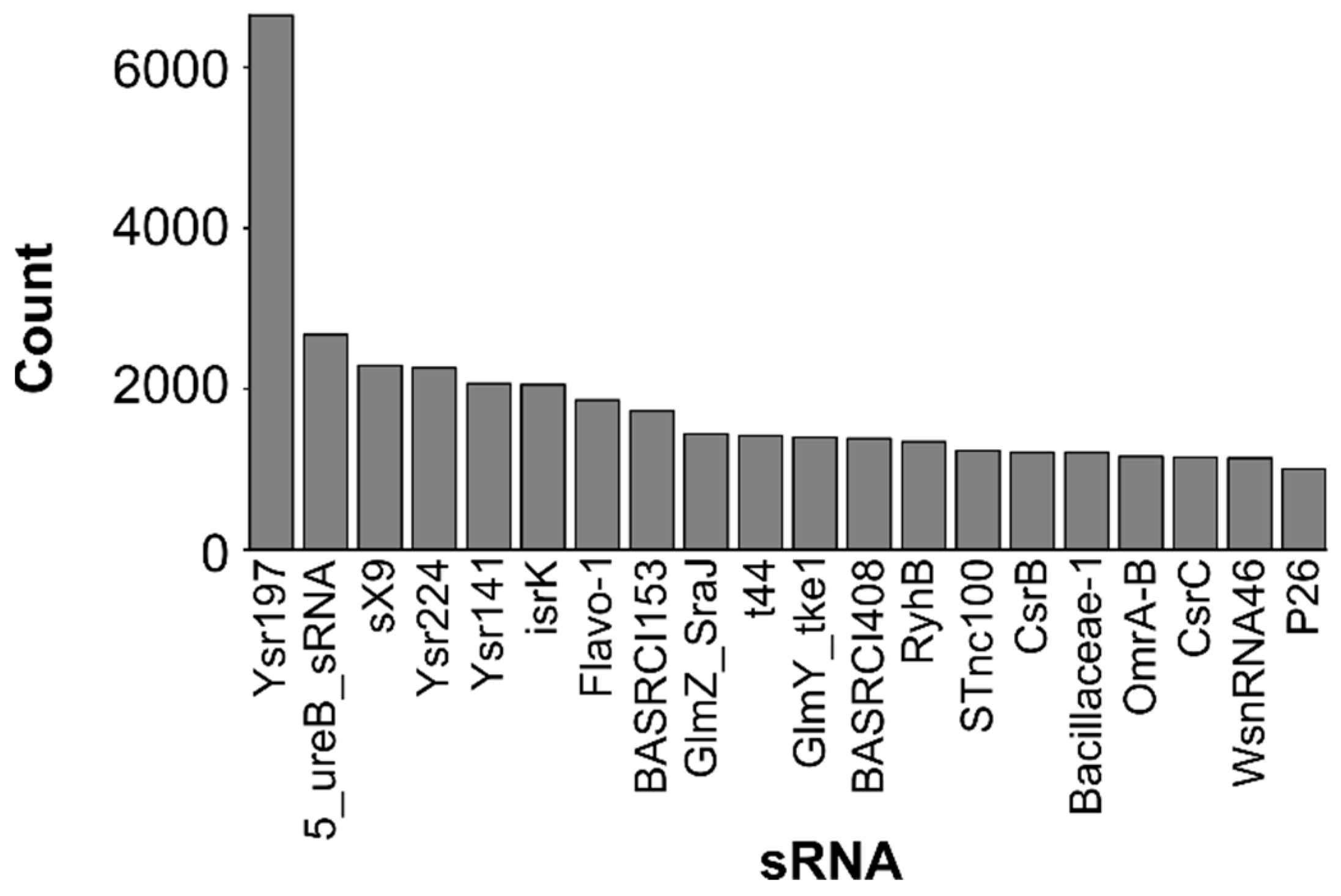
2.2. Most Abundant Small RNAs
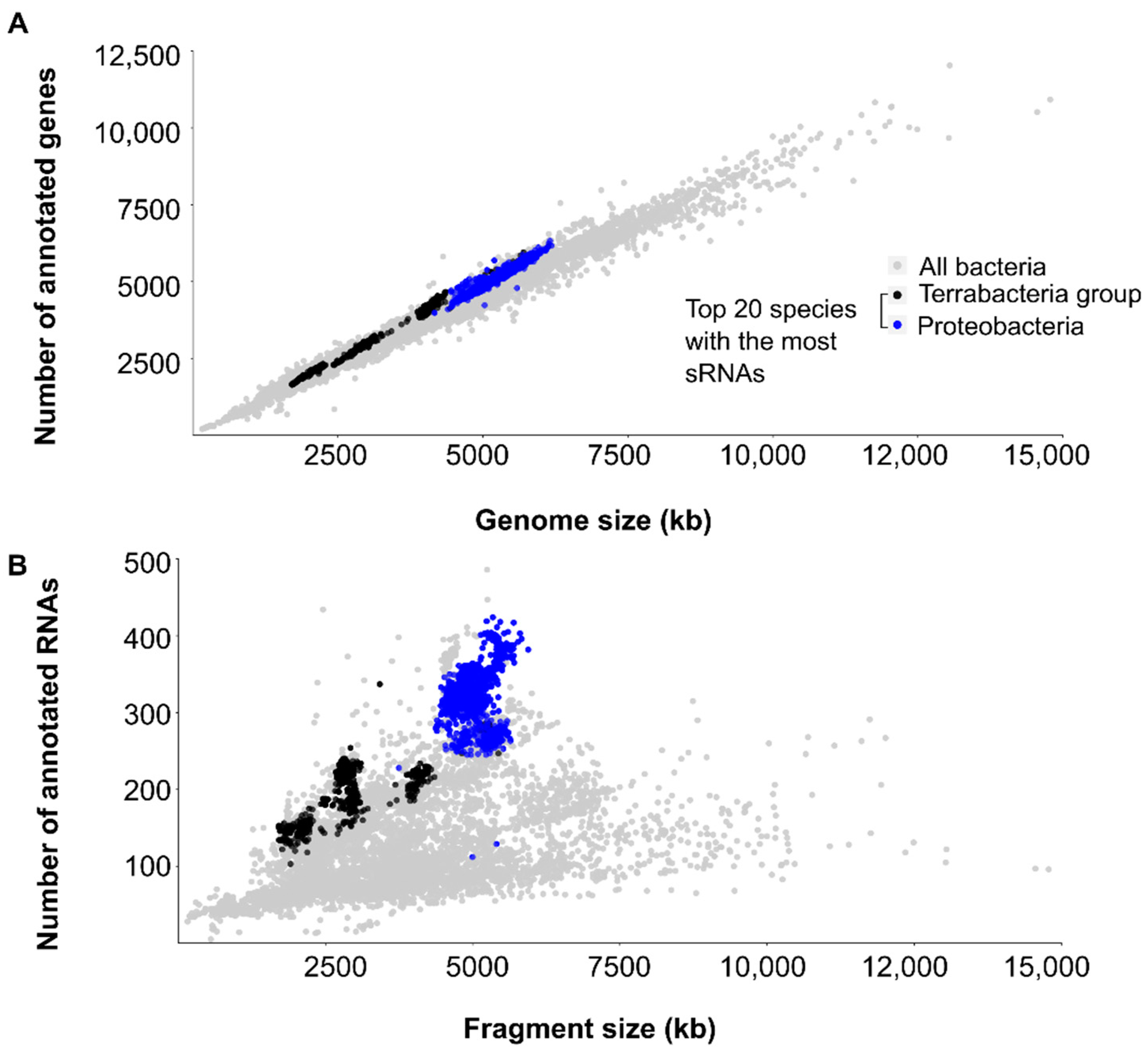
3. Biases towards Model Organisms and Pathogens
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jørgensen, M.G.; Pettersen, J.S.; Kallipolitis, B.H. sRNA-mediated control in bacteria: An increasing diversity of regulatory mechanisms. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194504. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.S.; Storz, G. Regulatory RNAs in Bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by Small RNAs in Bacteria: Expanding Frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.P.; Storz, G. Prevalence of small base-pairing RNAs derived from diverse genomic loci. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194524. [Google Scholar] [CrossRef]
- Heidrich, N.; Moll, I.; Brantl, S. In vitro analysis of the interaction between the small RNA SR1 and its primary target ahrC mRNA. Nucleic Acids Res. 2007, 35, 4331–4346. [Google Scholar] [CrossRef] [PubMed]
- Majdalani, N.; Chen, S.; Murrow, J.; St John, K.; Gottesman, S. Regulation of RpoS by a novel small RNA: The characterization of RprA. Mol. Microbiol. 2001, 39, 1382–1394. [Google Scholar] [CrossRef]
- Morita, T.; Maki, K.; Aiba, H. RNase E-based ribonucleoprotein complexes: Mechanical basis of mRNA destabilization mediated by bacterial noncoding RNAs. Genes Dev. 2005, 19, 2176–2186. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, V.; Papenfort, K.; Lucchini, S.; Hinton, J.; Vogel, J. Coding sequence targeting by MicC RNA reveals bacterial mRNA silencing downstream of translational initiation. Nat. Struct. Mol. Biol. 2009, 16, 840–846. [Google Scholar] [CrossRef]
- Sun, X.; Zhulin, I.; Wartell, R.M. Predicted structure and phyletic distribution of the RNA-binding protein Hfq. Nucleic Acids Res. 2002, 30, 3662–3671. [Google Scholar] [CrossRef] [Green Version]
- Christopoulou, N.; Granneman, S. The role of RNA-binding proteins in mediating adaptive responses in Gram-positive bacteria. FEBS J. 2021, 289, 1746–1764. [Google Scholar] [CrossRef]
- Müller, P.; Gimpel, M.; Wildenhain, T.; Brantl, S. A new role for CsrA: Promotion of complex formation between an sRNA and its mRNA target in Bacillus subtilis. RNA Biol. 2019, 16, 972–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.; A Forst, S.; Zhao, K.; Inouye, M.; Delihas, N. The function of micF RNA. micF RNA is a major factor in the thermal regulation of OmpF protein in Escherichia coli. J. Biol. Chem. 1989, 264, 17961–17970. [Google Scholar] [CrossRef]
- Cohen, S.P.; McMurry, L.M.; Levy, S.B. marA locus causes decreased expression of OmpF porin in multiple-antibiotic-resistant (Mar) mutants of Escherichia coli. J. Bacteriol. 1988, 170, 5416–5422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, T.; Chou, M.Y.; Inouye, M. A unique mechanism regulating gene expression: Translational inhibition by a complementary RNA transcript (micRNA). Proc. Natl. Acad. Sci. USA 1984, 81, 1966–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naghdi, M.R.; Smail, K.; Wang, J.X.; Wade, F.; Breaker, R.R.; Perreault, J. Search for 5’-leader regulatory RNA structures based on gene annotation aided by the RiboGap database. Methods 2017, 117, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalvari, I.; Nawrocki, E.P.; Ontiveros-Palacios, N.; Argasinska, J.; Lamkiewicz, K.; Marz, M.; Griffiths-Jones, S.; Toffano-Nioche, C.; Gautheret, D.; Weinberg, Z.; et al. Rfam 14: Expanded coverage of metagenomic, viral and microRNA families. Nucleic Acids Res. 2020, 49, D192–D200. [Google Scholar] [CrossRef]
- Sayers, E.W.; Barrett, T.; Benson, D.A.; Bolton, E.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Federhen, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2010, 39, D38–D51. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-Y.; Chang, H.-Y.; Chou, C.-H.; Tseng, C.-P.; Ho, S.-Y.; Yang, C.-D.; Ju, Y.-W.; Huang, H.-D. sRNAMap: Genomic maps for small non-coding RNAs, their regulators and their targets in microbial genomes. Nucleic Acids Res. 2008, 37, D150–D154. [Google Scholar] [CrossRef]
- Pischimarov, J.; Kuenne, C.; Billion, A.; Hemberger, J.; Cemič, F.; Chakraborty, T.; Hain, T. sRNAdb: A small non-coding RNA database for gram-positive bacteria. BMC Genom. 2012, 13, 384. [Google Scholar] [CrossRef] [Green Version]
- Santos-Zavaleta, A.; Salgado, H.; Gama-Castro, S.; Sánchez-Pérez, M.; Gómez-Romero, L.; Ledezma-Tejeida, D.; García-Sotelo, J.S.; Alquicira-Hernández, K.; Muñiz-Rascado, L.J.; Peña-Loredo, P.; et al. RegulonDB v 10.5: Tackling challenges to unify classic and high throughput knowledge of gene regulation in E. coli K-12. Nucleic Acids Res. 2019, 47, D212–D220. [Google Scholar] [CrossRef] [Green Version]
- Keseler, I.M.; Bonavides-Martinez, C.; Collado-Vides, J.; Gama-Castro, S.; Gunsalus, R.P.; Johnson, D.A.; Krummenacker, M.; Nolan, L.; Paley, S.; Paulsen, I.; et al. EcoCyc: A comprehensive view of Escherichia coli biology. Nucleic Acids Res. 2008, 37, D464–D470. [Google Scholar] [CrossRef] [PubMed]
- Sassi, M.; Augagneur, Y.; Mauro, T.; Ivain, L.; Chabelskaya, S.; Hallier, M.; Sallou, O.; Felden, B. SRD: A Staphylococcus regulatory RNA database. RNA 2015, 21, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Huang, D.; Cheung, M.K.; Nong, W.; Huang, Q.; Kwan, H.S. BSRD: A repository for bacterial small regulatory RNA. Nucleic Acids Res. 2012, 41, D233–D238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistuzzi, F.U.; Hedges, S.B. A Major Clade of Prokaryotes with Ancient Adaptations to Life on Land. Mol. Biol. Evol. 2008, 26, 335–343. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. BioMed. Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Robinson, D. Chlamydia trachomatis and sexually transmitted disease. BMJ 1994, 308, 150–151. [Google Scholar] [CrossRef] [Green Version]
- Weinstock, G.M.; Hardham, J.M.; McLeod, M.P.; Sodergren, E.J.; Norris, S.J. The genome of Treponema pallidum: New light on the agent of syphilis. FEMS Microbiol. Rev. 1998, 22, 323–332. [Google Scholar] [CrossRef]
- Le Moal, G.; Landron, C.; Grollier, G.; Robert, R.; Burucoa, C. Meningitis due to Capnocytophaga canimorsus after receipt of a dog bite: Case report and review of the literature. Clin. Infect. Dis. 2003, 36, e42–e46. [Google Scholar] [CrossRef] [Green Version]
- Sena-Vélez, M.; Holland, S.D.; Aggarwal, M.; Cogan, N.G.; Jain, M.; Gabriel, D.W.; Jones, K.M. Growth dynamics and survival of Liberibacter crescens BT-1, an important model organism for the citrus Huanglongbing pathogen “Candidatus Liberibacter asiaticus”. Appl. Environ. Microbiol. 2019, 85, e01656-19. [Google Scholar] [CrossRef] [Green Version]
- Hess, C.; Enichlmayr, H.; Jandreski-Cvetkovic, D.; Liebhart, D.; Bilic, I.; Hess, M. Riemerella anatipestifer outbreaks in commercial goose flocks and identification of isolates by MALDI-TOF mass spectrometry. Avian Pathol. 2013, 42, 151–156. [Google Scholar] [CrossRef]
- Saad, T.; Atallah, S. Studies on bacterial infection in marine fish. J. Arab. Aquac. Soc. 2014, 374, 1–20. [Google Scholar] [CrossRef]
- Marnocha, C.L.; Levy, A.T.; Powell, D.H.; Hanson, T.E.; Chan, C.S. Mechanisms of extracellular S0 globule production and degradation in Chlorobaculum tepidum via dynamic cell–globule interactions. Microbiology 2016, 162, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Wunsch, C.M.; Lewis, J.P. Porphyromonas gingivalis as a Model Organism for Assessing Interaction of Anaerobic Bacteria with Host Cells. J. Vis. Exp. 2015, 106, e53408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albrecht, M.; Sharma, C.M.; Reinhardt, R.; Vogel, J.; Rudel, T. Deep sequencing-based discovery of the Chlamydia trachomatis transcriptome. Nucleic Acids Res. 2009, 38, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, J.; Rao, G.V.; Runac, J.; Hackstadt, T.; Grieshaber, S.S.; Grieshaber, N.A. Translation Inhibition of the Developmental Cycle Protein HctA by the Small RNA IhtA Is Conserved across Chlamydia. PLoS ONE 2012, 7, e47439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Fernandez, M.F.; Eoyang, L.; August, J. Factor fraction required for the synthesis of bacteriophage Qβ-RNA. Nature 1968, 219, 588–590. [Google Scholar] [CrossRef]
- Olejniczak, M.; Storz, G. ProQ/FinO-domain proteins: Another ubiquitous family of RNA matchmakers? Mol. Microbiol. 2017, 104, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Blount, Z.D. The natural history of model organisms: The unexhausted potential of E. coli. Elife 2015, 4, e05826. [Google Scholar] [CrossRef]
- Crick, F.; Barnett, L.; Brenner, S.; Watts-Tobin, R.J. General nature of the genetic code for proteins. Nature 1961, 192, 1227–1232. [Google Scholar] [CrossRef]
- Garai, P.; Gnanadhas, D.P.; Chakravortty, D. Salmonella enterica serovars Typhimurium and Typhi as model organisms: Revealing paradigm of host-pathogen interactions. Virulence 2012, 3, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Padalon-Brauch, G.; Hershberg, R.; Elgrably-Weiss, M.; Baruch, K.; Rosenshine, I.; Margalit, H.; Altuvia, S. Small RNAs encoded within genetic islands of Salmonella typhimurium show host-induced expression and role in virulence. Nucleic Acids Res. 2008, 36, 1913–1927. [Google Scholar] [CrossRef] [PubMed]
- Archer, G.L. Staphylococcus aureus: A well-armed pathogen. Rev. Infect. Dis. 1998, 26, 1179–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drevets, D.A.; Bronze, M.S. Listeria monocytogenes: Epidemiology, human disease, and mechanisms of brain invasion. FEMS Immunol. Med. Microbiol. 2008, 53, 151–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyściak, W.; Pluskwa, K.; Jurczak, A.; Kościelniak, D. The pathogenicity of the Streptococcus genus. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1361–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errington, J.; van der Aart, L.T. Microbe Profile: Bacillus subtilis: Model organism for cellular development, and industrial workhorse. Microbiology 2020, 166, 425. [Google Scholar] [CrossRef]
- Ranjan, K.P.; Ranjan, N. Citrobacter: An emerging health care associated urinary pathogen. Urol. Ann. 2013, 5, 313–314. [Google Scholar] [CrossRef]
- Killackey, S.A.; Sorbara, M.T.; Girardin, S.E. Cellular Aspects of Shigella Pathogenesis: Focus on the Manipulation of Host Cell Processes. Front. Cell. Infect. Microbiol. 2016, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Sanders, W.E., Jr.; Sanders, C.C. Enterobacter spp.: Pathogens poised to flourish at the turn of the century. Clin. Microbiol. Rev. 1997, 10, 220–241. [Google Scholar] [CrossRef]
- Bi, D.; Jiang, X.; Sheng, Z.-K.; Ngmenterebo, D.; Tai, C.; Wang, M. Mapping the resistance-associated mobilome of a carbapenem-resistant Klebsiella pneumoniae strain reveals insights into factors shaping these regions and facilitates generation of a ‘resistance-disarmed’model organism. J. Antimicrob. Chemother. 2015, 70, 2770–2774. [Google Scholar] [CrossRef] [Green Version]
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [Green Version]
- Achtman, M.; Zurth, K.; Morelli, G.; Torrea, G.; Guiyoule, A.; Carniel, E. Yersinia pestis, the cause of plague, is a recently emerged clone of Yersinia pseudotuberculosis. Proc. Natl. Acad. Sci. USA 1999, 96, 14043–14048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuss, A.M.; Heroven, A.K.; Waldmann, B.; Reinkensmeier, J.; Jarek, M.; Beckstette, M.; Dersch, P. Transcriptomic Profiling of Yersinia pseudotuberculosis Reveals Reprogramming of the Crp Regulon by Temperature and Uncovers Crp as a Master Regulator of Small RNAs. PLoS Genet. 2015, 11, e1005087. [Google Scholar] [CrossRef] [PubMed]
- Arnvig, K.B.; Young, D.B. Identification of small RNAs in Mycobacterium tuberculosis. Mol. Microbiol. 2009, 73, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Righetti, F.; Nuss, A.M.; Twittenhoff, C.; Beele, S.; Urban, K.; Will, S.; Bernhart, S.H.; Stadler, P.F.; Dersch, P.; Narberhaus, F. Temperature-responsive in vitro RNA structurome of Yersinia pseudotuberculosis. Proc. Natl. Acad. Sci. USA 2016, 113, 7237–7242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiano, C.A.; Koo, J.T.; Schipma, M.J.; Caulfield, A.J.; Jafari, N.; Lathem, W.W. Genome-Wide Analysis of Small RNAs Expressed by Yersinia pestis Identifies a Regulator of the Yop-Ysc Type III Secretion System. J. Bacteriol. 2014, 196, 1659–1670. [Google Scholar] [CrossRef] [Green Version]
- Sittka, A.; Lucchini, S.; Papenfort, K.; Sharma, C.M.; Rolle, K.; Binnewies, T.T.; Hinton, J.C.D.; Vogel, J. Deep Sequencing Analysis of Small Noncoding RNA and mRNA Targets of the Global Post-Transcriptional Regulator, Hfq. PLoS Genet. 2008, 4, e1000163. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Peng, X.; Wang, N.; Wu, Q. Identification of novel sRNAs in Brucella abortus 2308. FEMS Microbiol. Lett. 2014, 354, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Schmidtke, C.; Findeiss, S.; Sharma, C.M.; Kuhfuß, J.; Hoffmann, S.; Vogel, J.; Stadler, P.F.; Bonas, U. Genome-wide transcriptome analysis of the plant pathogen Xanthomonas identifies sRNAs with putative virulence functions. Nucleic Acids Res. 2011, 40, 2020–2031. [Google Scholar] [CrossRef]
- Mayoral, J.G.; Hussain, M.; Joubert, D.A.; Iturbe-Ormaetxe, I.; O’Neill, S.L.; Asgari, S. Wolbachia small noncoding RNAs and their role in cross-kingdom communications. Proc. Natl. Acad. Sci. USA 2014, 111, 18721–18726. [Google Scholar] [CrossRef] [Green Version]
- Reichenbach, B.; Maes, A.; Kalamorz, F.; Hajnsdorf, E.; Görke, B. The small RNA GlmY acts upstream of the sRNA GlmZ in the activation of glmS expression and is subject to regulation by polyadenylation in Escherichia coli. Nucleic Acids Res. 2008, 36, 2570–2580. [Google Scholar] [CrossRef]
- Rivas, E.; Klein, R.J.; Jones, T.A.; Eddy, S.R. Computational identification of noncoding RNAs in E. coli by comparative genomics. Curr. Biol. 2001, 11, 1369–1373. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.H.; Vogel, J. Two Seemingly Homologous Noncoding RNAs Act Hierarchically to Activate glmS mRNA Translation. PLOS Biol. 2008, 6, e64. [Google Scholar] [CrossRef] [PubMed]
- Tjaden, B.; Saxena, R.M.; Stolyar, S.; Haynor, D.R.; Kolker, E.; Rosenow, C. Transcriptome analysis of Escherichia coli using high-density oligonucleotide probe arrays. Nucleic Acids Res. 2002, 30, 3732–3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argaman, L.; Hershberg, R.; Vogel, J.; Bejerano, G.; Wagner, E.H.; Margalit, H.; Altuvia, S. Novel small RNA-encoding genes in the intergenic regions of Escherichia coli. Curr. Biol. 2001, 11, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.M.; Quinones, M.; Pratt, J.; Ding, Y.; Waldor, M.K. Characterization of the Small Untranslated RNA RyhB and Its Regulon in Vibrio cholerae. J. Bacteriol. 2005, 187, 4005–4014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottesman, S. Micros for microbes: Non-coding regulatory RNAs in bacteria. Trends Genet. 2005, 21, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef] [Green Version]
- Neuhaus, K.; Landstorfer, R.; Simon, S.; Schober, S.; Wright, P.R.; Smith, C.; Backofen, R.; Wecko, R.; Keim, D.A.; Scherer, S. Differentiation of ncRNAs from small mRNAs in Escherichia coli O157:H7 EDL933 (EHEC) by combined RNAseq and RIBOseq—ryhB encodes the regulatory RNA RyhB and a peptide, RyhP. BMC Genom. 2017, 18, 216. [Google Scholar] [CrossRef] [Green Version]
- Porcheron, G.; Habib, R.; Houle, S.; Caza, M.; Lépine, F.; Daigle, F.; Massé, E.; Dozois, C.M. The Small RNA RyhB Contributes to Siderophore Production and Virulence of Uropathogenic Escherichia coli. Infect. Immun. 2014, 82, 5056–5068. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, S.; Wu, N.; Yuan, Y.; Zhang, W.; Zhang, Y. Small Non-coding RNA RyhB Mediates Persistence to Multiple Antibiotics and Stresses in Uropathogenic Escherichia coli by Reducing Cellular Metabolism. Front. Microbiol. 2018, 9, 136. [Google Scholar] [CrossRef]
- Cui, Y.; Chatterjee, A.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Identification of a global repressor gene, rsmA, of Erwinia carotovora subsp. carotovora that controls extracellular enzymes, N-(3-oxohexanoyl)-L-homoserine lactone, and pathogenicity in soft-rotting Erwinia spp. J. Bacteriol. 1995, 177, 5108–5115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Chatterjee, A.; Yang, H.; Chatterjee, A.K. Regulatory network controlling extracellular proteins in Erwinia carotovora subsp. carotovora: FlhDC, the master regulator of flagellar genes, activates rsmB regulatory RNA production by affecting gacA and hexA (lrhA) expression. J. Bacteriol. 2008, 190, 4610–4623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heroven, A.K.; Sest, M.; Pisano, F.; Scheb-Wetzel, M.; Steinmann, R.; Böhme, K.; Klein, J.; Münch, R.; Schomburg, D.; Dersch, P. Crp Induces Switching of the CsrB and CsrC RNAs in Yersinia pseudotuberculosis and Links Nutritional Status to Virulence. Front. Cell. Infect. Microbiol. 2012, 2, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.Y.; Gui, G.; Wei, B.; Preston, J.F.; Oakford, L.; Yüksel, U.; Giedroc, D.; Romeo, T. The RNA Molecule CsrB Binds to the Global Regulatory Protein CsrA and Antagonizes Its Activity in Escherichia coli. J. Biol. Chem. 1997, 272, 17502–17510. [Google Scholar] [CrossRef] [Green Version]
- Mei, L.; Xu, S.; Lu, P.; Lin, H.; Guo, Y.; Wang, Y. CsrB, a noncoding regulatory RNA, is required for BarA-dependent expression of biocontrol traits in Rahnella aquatilis HX2. PLoS ONE 2017, 12, e0187492. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Peng, Q.; Zhang, Q.; Yi, X.; Choi, C.J.; Reedy, R.M.; Charkowski, A.O.; Yang, C.-H. Dynamic Regulation of GacA in Type III Secretion, Pectinase Gene Expression, Pellicle Formation, and Pathogenicity of Dickeya dadantii (Erwinia chrysanthemi 3937). Mol. Plant-Microbe Interact. 2008, 21, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Khokhani, D.; Wu, X.; Yang, F.; Biener, G.; Koestler, B.J. Cross-talk between a regulatory small RNA, cyclic-di-GMP signalling and flagellar regulator FlhDC for virulence and bacterial behaviours. Environ. Microbiol. 2015, 17, 4745–4763. [Google Scholar] [CrossRef]
- Guillier, M.; Gottesman, S. Remodelling of the Escherichia coli outer membrane by two small regulatory RNAs. Mol. Microbiol. 2005, 59, 231–247. [Google Scholar] [CrossRef]
- Holmqvist, E.; Reimegård, J.; Sterk, M.; Grantcharova, N.; Römling, U.; Wagner, E.G.H. Two antisense RNAs target the transcriptional regulator CsgD to inhibit curli synthesis. EMBO J. 2010, 29, 1840–1850. [Google Scholar] [CrossRef] [Green Version]
- Wassarman, K.M.; Repoila, F.; Rosenow, C.; Storz, G.; Gottesman, S. Identification of novel small RNAs using comparative genomics and microarrays. Genes Dev. 2001, 15, 1637–1651. [Google Scholar] [CrossRef] [Green Version]
- Weilbacher, T.; Suzuki, K.; Dubey, A.; Wang, X.; Gudapaty, S.; Morozov, I.; Baker, C.S.; Nagdellis, D.; Babitzke, P.; Romeo, T. A novel sRNA component of the carbon storage regulatory system of Escherichia coli. Mol. Microbiol. 2003, 48, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.; Wang, J.X.; Bogue, J.; Yang, J.; Corbino, K.; Moy, R.H.; Breaker, R.R. Comparative genomics reveals 104 candidate structured RNAs from bacteria, archaea, and their metagenomes. Genome Biol. 2010, 11, R31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livny, J.; Brencic, A.; Lory, S.; Waldor, M.K. Identification of 17 Pseudomonas aeruginosa sRNAs and prediction of sRNA-encoding genes in 10 diverse pathogens using the bioinformatic tool sRNAPredict2. Nucleic Acids Res. 2006, 34, 3484–3493. [Google Scholar] [CrossRef]
- Wen, Y.; Feng, J.; Sachs, G. Helicobacter pylori 5′ureB-sRNA, a cis-Encoded Antisense Small RNA, Negatively Regulates ureAB Expression by Transcription Termination. J. Bacteriol. 2012, 195, 444–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuilleumier, S.; Chistoserdova, L.; Lee, M.-C.; Bringel, F.; Lajus, A.; Zhou, Y.; Gourion, B.; Barbe, V.; Chang, J.; Cruveiller, S.; et al. Methylobacterium Genome Sequences: A Reference Blueprint to Investigate Microbial Metabolism of C1 Compounds from Natural and Industrial Sources. PLoS ONE 2009, 4, e5584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saïdi, F.; Jolivet, N.Y.; Lemon, D.J.; Nakamura, A.; Belgrave, A.M.; Garza, A.G.; Veyrier, F.J.; Islam, S.T. Bacterial glycocalyx integrity drives multicellular swarm biofilm dynamism. Mol. Microbiol. 2021, 116, 1151–1172. [Google Scholar] [CrossRef] [PubMed]
- Setubal, J.C.; dos Santos, P.; Goldman, B.S.; Ertesvåg, H.; Espin, G.; Rubio, L.M.; Valla, S.; Almeida, N.; Balasubramanian, D.; Cromes, L.; et al. Genome Sequence of Azotobacter vinelandii, an Obligate Aerobe Specialized To Support Diverse Anaerobic Metabolic Processes. J. Bacteriol. 2009, 191, 4534–4545. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, C.A., III; Chuang, R.-Y.; Noskov, V.N.; Assad-Garcia, N.; Deerinck, T.J.; Ellisman, M.H. Design and synthesis of a minimal bacterial genome. Science 2016, 351, aad6253. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, H.; Masroor, T.; Parmar, S.A.; Panigrahi, D. Urinary tract infection by a rare pathogen Cedecea neteri in a pregnant female with Polyhydramnios: Rare case report from UAE. BMC Infect. Dis. 2021, 21, 637. [Google Scholar] [CrossRef]
- Aiba, H. Mechanism of RNA silencing by Hfq-binding small RNAs. Curr. Opin. Microbiol. 2007, 10, 134–139. [Google Scholar] [CrossRef]
- Bouché, F.; Bouché, J.P. Genetic evidence that DicF, a second division inhibitor encoded by the Escherichia coli dicB operon, is probably RNA. Mol. Microbiol. 1989, 3, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.; Kontomichalou, P. Penicillinase synthesis controlled by infectious R factors in Enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Delihas, N.; Forst, S. MicF: An antisense RNA gene involved in response of Escherichia coli to global stress factors. J. Mol. Biol. 2001, 313, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dühring, U.; Axmann, I.M.; Hess, W.R.; Wilde, A. An internal antisense RNA regulates expression of the photosynthesis gene isiA. Proc. Natl. Acad. Sci. USA 2006, 103, 7054–7058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faubladier, M.; Bouché, J.-P. Division inhibition gene dicF of Escherichia coli reveals a widespread group of prophage sequences in bacterial genomes. J. Bacteriol. 1994, 176, 1150–1156. [Google Scholar] [CrossRef] [Green Version]
- Georg, J.; Hess, W.R. cis-antisense RNA, another level of gene regulation in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Gerhart, E.; Wagner, H.; Nordström, K. Structural analysis of an RNA molecule involved in replication control of plasmid Rl. Nucleic Acids Res. 1986, 14, 2523–2538. [Google Scholar] [CrossRef]
- Hajjar, R.; Ambaraghassi, G.; Sebajang, H.; Schwenter, F.; Su, S.-H. Raoultella ornithinolytica: Emergence and resistance. Infect. Drug Resist. 2020, 13, 1091. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Liu, X.; Law, C.O.; Wang, Y.; Lo, W.U.; Weng, X.; Chan, T.F.; Ho, P.; Lau, T.C. The CTX-M-14 plasmid pHK01 encodes novel small RNAs and influences host growth and motility. FEMS Microbiol. Ecol. 2017, 93, fix090. [Google Scholar] [CrossRef]
- Khanna, A.; Khanna, M.; Aggarwal, A. Serratia marcescens-a rare opportunistic nosocomial pathogen and measures to limit its spread in hospitalized patients. J. Clin. Diagn. Res. JCDR 2013, 7, 243. [Google Scholar]
- Kittle, J.; Simons, R.W.; Lee, J.; Kleckner, N. Insertion sequence IS10 anti-sense pairing initiates by an interaction between the 5′ end of the target RNA and a loop in the anti-sense RNA. J. Mol. Biol. 1989, 210, 561–572. [Google Scholar] [CrossRef]
- Kluyver, T.; Ragan-Kelley, B.; Pérez, F.; Granger, B.E.; Bussonnier, M.; Frederic, J.; Kelley, K.; Hamrick, J.B.; Grout, J.; Corlay, S. Jupyter Notebooks—A Publishing Format for Reproducible Computational Workflows; IOS Press: Amsterdam, The Netherlands, 2016; Volume 2016. [Google Scholar]
- Light, J.; Molin, S. Post-transcriptional control of expression of the repA gene of plasmid R1 mediated by a small RNA molecule. EMBO J. 1983, 2, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.M.; Livny, J.; Lawrence, M.S.; Kimball, M.D.; Waldor, M.K.; Camilli, A. Experimental discovery of sRNAs in Vibrio cholerae by direct cloning, 5S/tRNA depletion and parallel sequencing. Nucleic Acids Res. 2009, 37, e46. [Google Scholar] [CrossRef] [PubMed]
- Mandin, P.; Repoila, F.; Vergassola, M.; Geissmann, T.; Cossart, P. Identification of new noncoding RNAs in Listeria monocytogenes and prediction of mRNA targets. Nucleic Acids Res. 2007, 35, 962–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashko, O.N.; Lin-Chao, S. Escherichia coli responds to environmental changes using enolasic degradosomes and stabilized DicF sRNA to alter cellular morphology. Proc. Natl. Acad. Sci. USA 2017, 114, E8025–E8034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordgren, S.; Slagter-Jäger, J.G.; Wagner, E.G.H. Real time kinetic studies of the interaction between folded antisense and target RNAs using surface plasmon resonance. J. Mol. Biol. 2001, 310, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.M.; Hoffmann, S.; Darfeuille, F.; Reignier, J.; Findeiß, S.; Sittka, A.; Chabas, S.; Reiche, K.; Hackermüller, J.; Reinhardt, R. The primary transcriptome of the major human pathogen Helicobacter pylori. Nature 2010, 464, 250–255. [Google Scholar] [CrossRef]
- Tétart, F.; Bouché, J.P. Regulation of the expression of the cell-cycle gene ftsZ by DicF antisense RNA. Division does not require a fixed number of FtsZ molecules. Mol. Microbiol. 1992, 6, 615–620. [Google Scholar] [CrossRef]
- Thomason, M.K.; Storz, G. Bacterial antisense RNAs: How many are there, and what are they doing? Annu. Rev. Genet. 2010, 44, 167–188. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.K.; Sharkady, S.M. Expanding spectrum of opportunistic Cedecea infections: Current clinical status and multidrug resistance. Int. J. Infect. Dis. 2020, 100, 461–469. [Google Scholar] [CrossRef]
- Vanderpool, C.K. Physiological consequences of small RNA-mediated regulation of glucose-phosphate stress. Curr. Opin. Microbiol. 2007, 10, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Vanderpool, C.K.; Gottesman, S. The novel transcription factor SgrR coordinates the response to glucose-phosphate stress. J. Bacteriol. 2007, 189, 2238–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Zhang, Q.; Zhang, Y.; Zhang, X.; Zhan, L.; Zhao, X.; Xu, S.; Sheng, X.; Huang, X. The novel cis-encoded antisense RNA AsrC positively regulates the expression of rpoE-rseABC operon and thus enhances the motility of Salmonella enterica serovar typhi. Front. Microbiol. 2015, 6, 990. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum Group | sRNAs |
|---|---|
| Acidobacteria | 4 |
| Aquificae | 1 |
| Calditrichaeota | 1 |
| Dictyoglomi | 1 |
| FCB group 1 | 16 |
| Fusobacteria | 2 |
| Nitrospirae | 3 |
| PVC group 2 | 8 |
| Proteobacteria | 345 |
| Spirochaetes | 6 |
| Synergistetes | 1 |
| Terrabacteria group | 210 |
| Thermodesulfobacteria | 1 |
| Thermotogae | 1 |
| Genus | Nb of Distinct sRNAs 1 | Description | Ref |
|---|---|---|---|
| Proteobacteria | |||
| Salmonella | 119 | Model organism to study host-pathogen interactions | [40] |
| Escherichia | 99 | Most well-understood bacteria | [38] |
| Citrobacter | 88 | Third most common urinary pathogen | [46] |
| Shigella | 85 | Causative pathogen of shigellosis | [47] |
| Enterobacter | 78 | Responsible for nosocomial infections | [48] |
| Klebsiella | 74 | Nosocomial pathogen, model organism to study drug resistance | [49] |
| Terrabacteria group | |||
| Streptococcus | 55 | Responsible for most cases of pneumonia worldwide | [44] |
| Staphylococcus | 46 | Most prevalent cause of infection in hospitalized patient | [42] |
| Listeria | 35 | Foodborne human pathogens causing central nervous system infections | [43] |
| Bacillus | 26 | Most-studied Gram-positive bacteria, model organisms for cellular development | [45] |
| Enterococcus | 25 | Principal cause of the healthcare-associated death worldwide | [48,50] |
| sRNA | Description | Rfam ID | sRNA Expression | Discovered in | Ref |
|---|---|---|---|---|---|
| Ysr197 | Yersinia sRNA 197 | RF02849 | Expressed in exponential phase | Yersinia pseudotuberculosis | [52] |
| 5_ureB_sRNA | - | RF02514 | Downregulate expression of operon ureAB | Helicobacter pylori | [84] |
| sX9 | Xanthomonas sRNA sX9 | RF02228 | - | Xanthomonas campestris pv. vesicatoria (Xcv) | [58] |
| Ysr224 | Yersinia sRNA 224 | RF02770 | Temperature-responsive | Yersinia pseudotuberculosis | [52,54] |
| Ysr141 | Yersinia sRNA 141 | RF02675 | Influence the expression of Yop-Ysc type III secretion system (T3SS) (critical system for virulence) | Yersinia pestis | [55] |
| isrK | isrK Hfq binding RNA | RF01394 | Stationary phase, low oxygen, low magnesium | Salmonella typhimurium | [41,56] |
| Flavo-1 | - | RF01705 | - | Bacteroidetes | [82] |
| BASRCI153 | Brucella sRNA CI153 | RF02604 | Putative target: BAB1_1361 | Brucella abortus | [57] |
| GlmZ_SraJ | GlmZ RNA activator of glmS mRNA | RF00083 | activator of glmS mRNA | Escherichia coli | [54,60,61,62] |
| t44 | - | RF00127 | - | Escherichia coli | [63] |
| GlmY_tke1 | GlmZ RNA activator of glmS mRNA | RF00128 | activator of glmS mRNA | Escherichia coli | [54,60,61,62] |
| BASRCI408 | Brucella sRNA CI408 | RF02599 | Putative target: BAB1_2002 | Brucella abortus | [57] |
| RyhB | - | RF00057 | Iron metabolism [67], regulates siderophore production and virulence [69], persistence regulation [70] | Escherichia coli | [64,65,66,67,68,69,70] |
| STnc100 | Gammaproteobacterial sRNA STnc100 | RF02076 | - | Salmonella sp. | [56] |
| CsrB | CsrB/RsmB RNA family | RF00018 | Binds the CrsA protein | Escherichia coli | [54,71,72,73,74,75,76,77] |
| Bacillaceae-1 | - | RF01690 | - | Bacteroidetes | [82] |
| OmrA-B | - | RF00079 | Target several genes encoding outer membrane proteins | Escherichia coli | [64,78,79,80] |
| CsrC | - | RF00084 | Binds the CrsA protein | Escherichia coli | [64,81] |
| Ysr276 | Yersinia sRNA 276 | RF02850 | - | Yersinia pseudotuberculosis | [52] |
| WsnRNA46 | Wolbachia sRNA 46 | RF02625 | Expressed in cells infected by parasitic microbe Wolbachia | Wolbachia sp. | [59] |
| P26 | Pseudomonas sRNA P26 | RF00630 | - | Pseudomonas aeruginosa | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boutet, E.; Djerroud, S.; Perreault, J. Small RNAs beyond Model Organisms: Have We Only Scratched the Surface? Int. J. Mol. Sci. 2022, 23, 4448. https://doi.org/10.3390/ijms23084448
Boutet E, Djerroud S, Perreault J. Small RNAs beyond Model Organisms: Have We Only Scratched the Surface? International Journal of Molecular Sciences. 2022; 23(8):4448. https://doi.org/10.3390/ijms23084448
Chicago/Turabian StyleBoutet, Emilie, Samia Djerroud, and Jonathan Perreault. 2022. "Small RNAs beyond Model Organisms: Have We Only Scratched the Surface?" International Journal of Molecular Sciences 23, no. 8: 4448. https://doi.org/10.3390/ijms23084448