
Physiological and Molecular Responses of Zoysia japonica to Rust Infection

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Whole-Plant Observations of Z. japonica Leaves after Inoculation
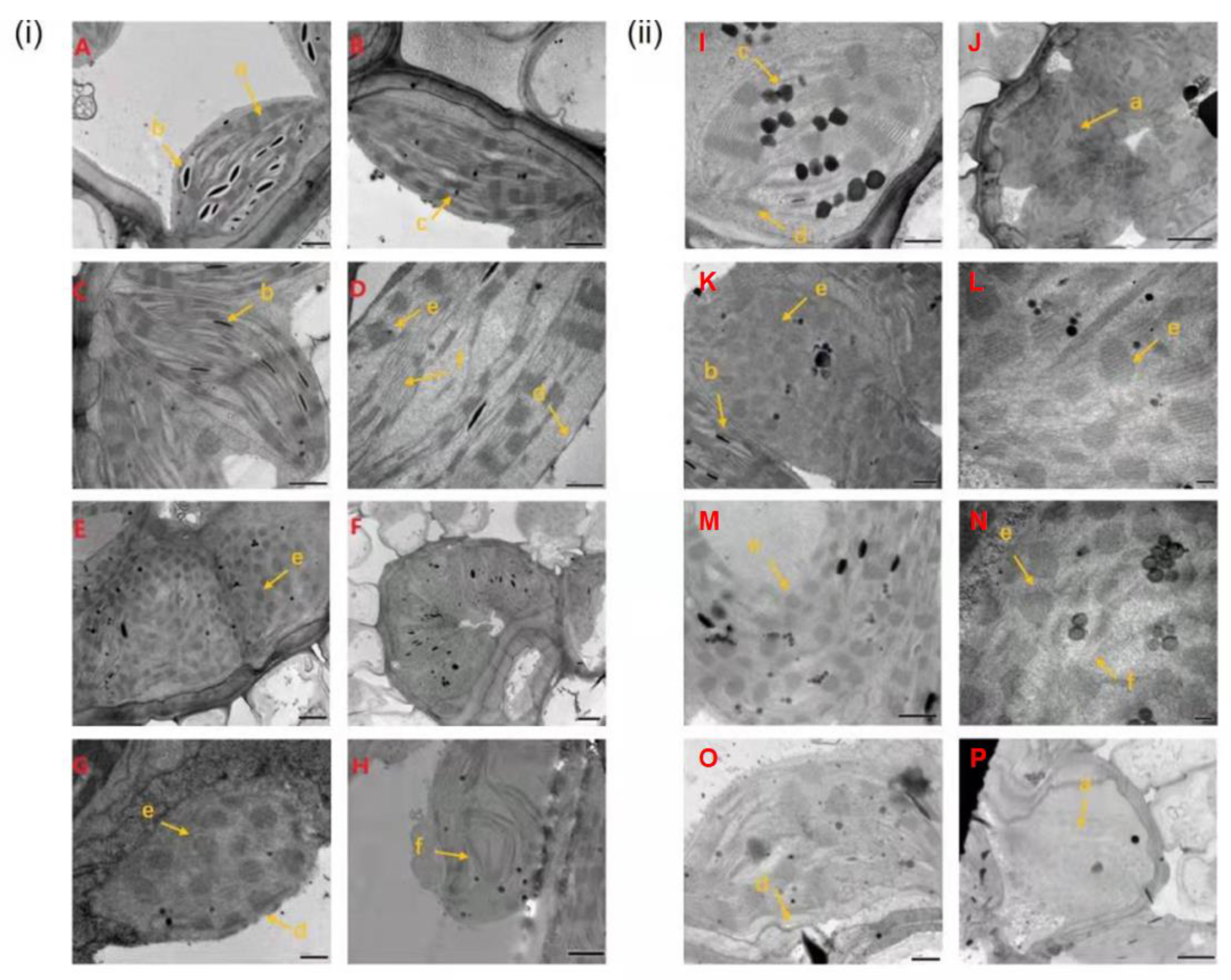
2.2. Cellular Observations of Z. japonica Leaves after Inoculation
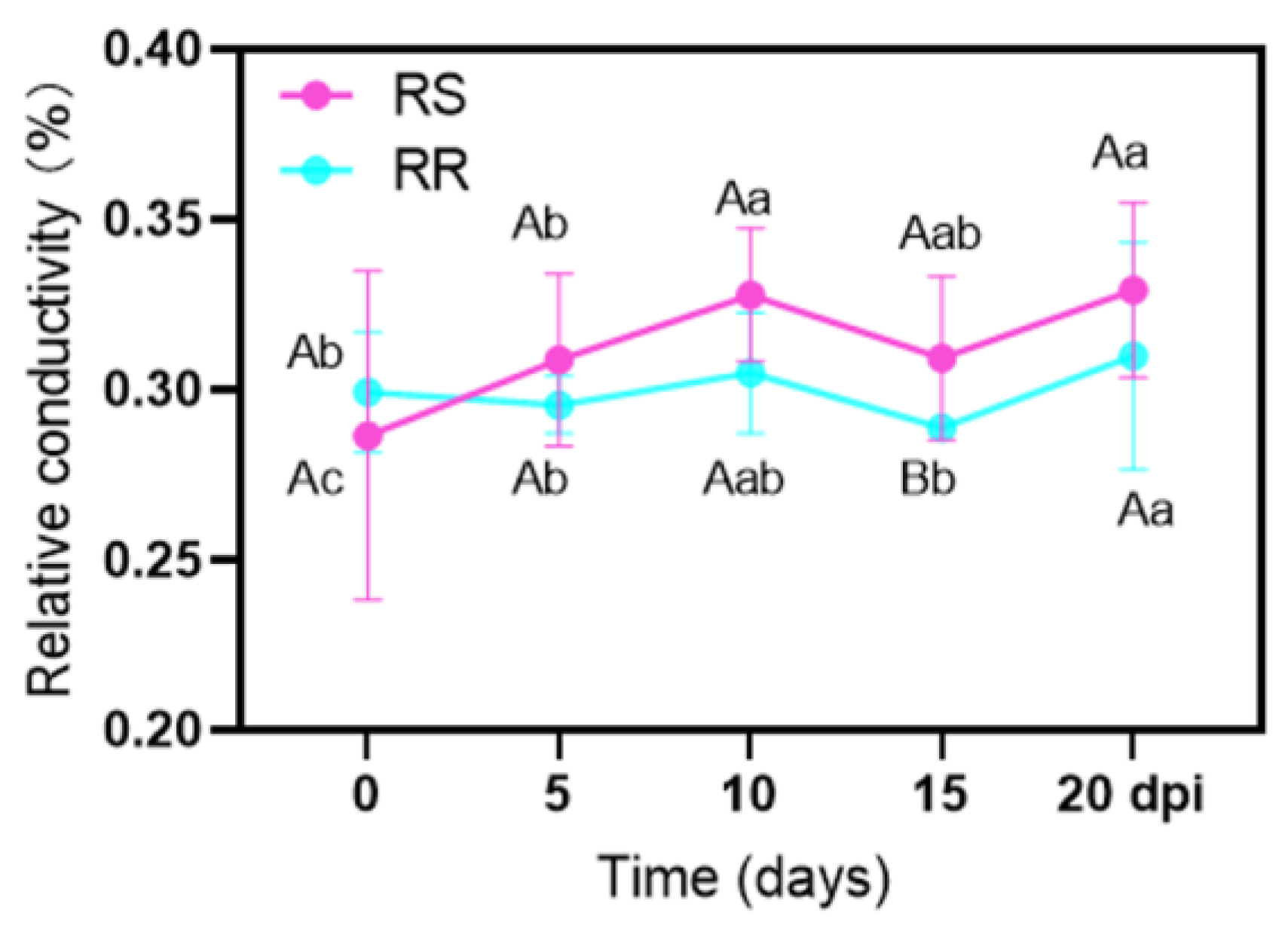
2.3. Electrolyte Leakage (EL) and Chlorophyll Content (Chl)
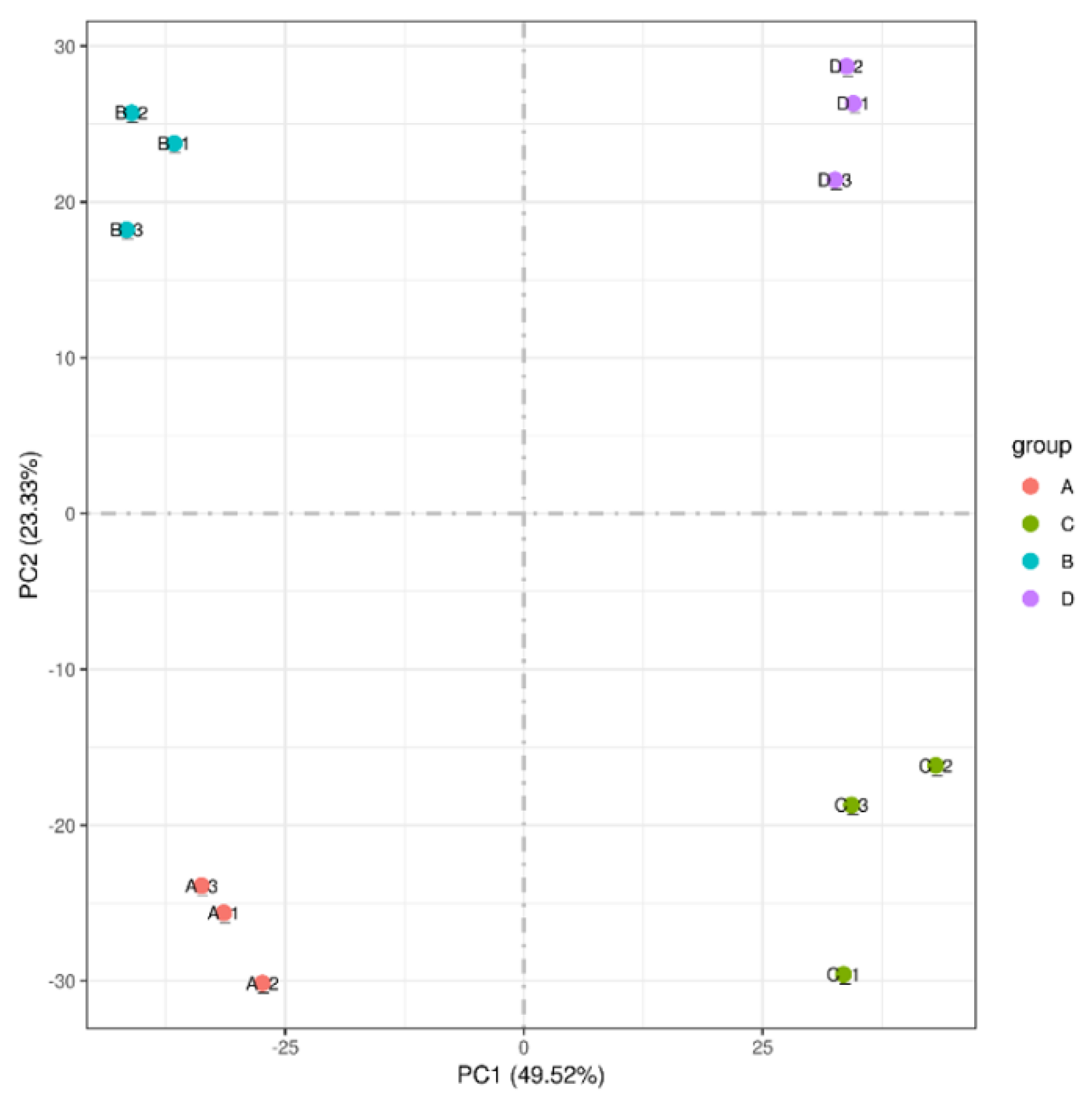
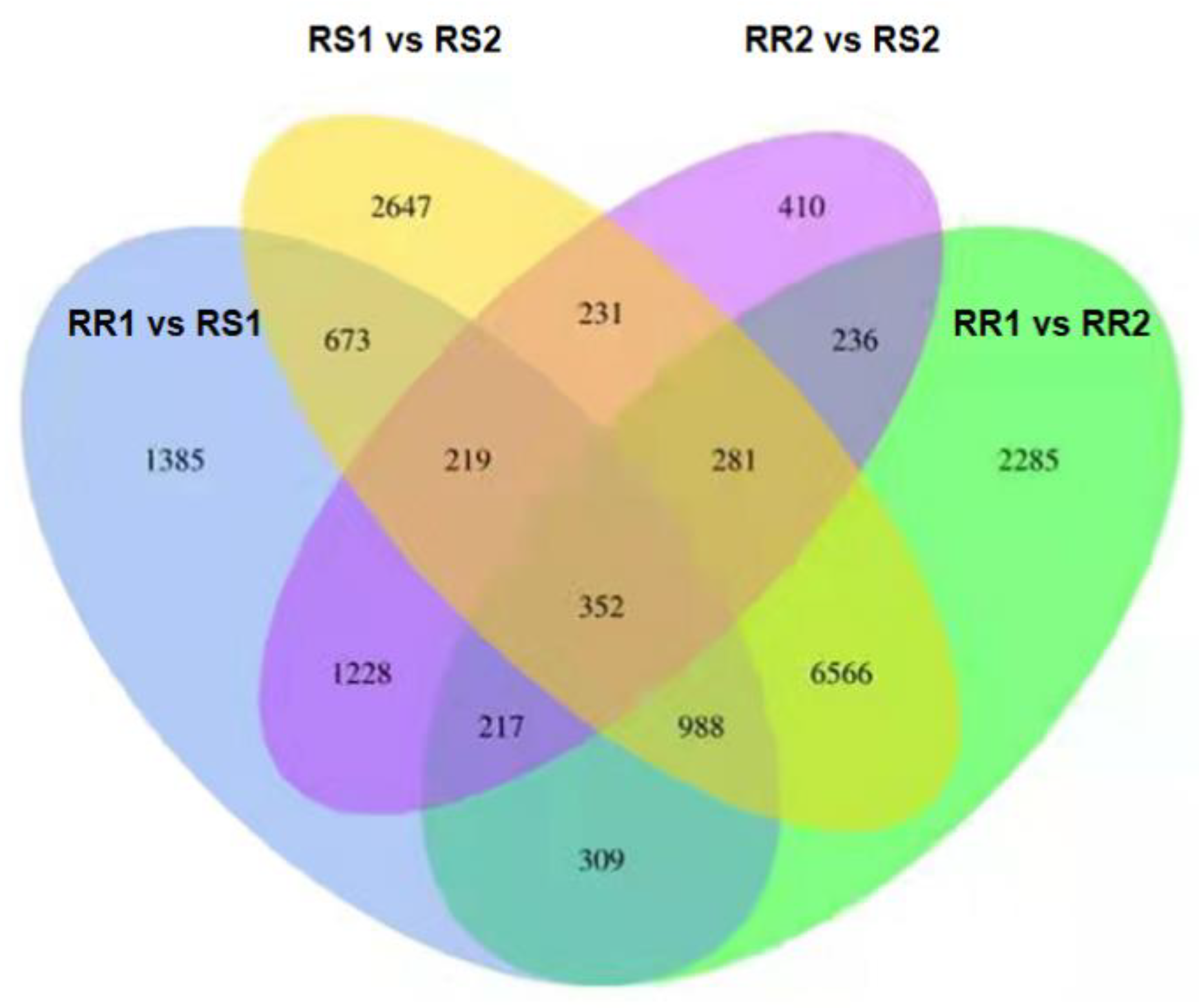
2.4. Transcriptome Analysis
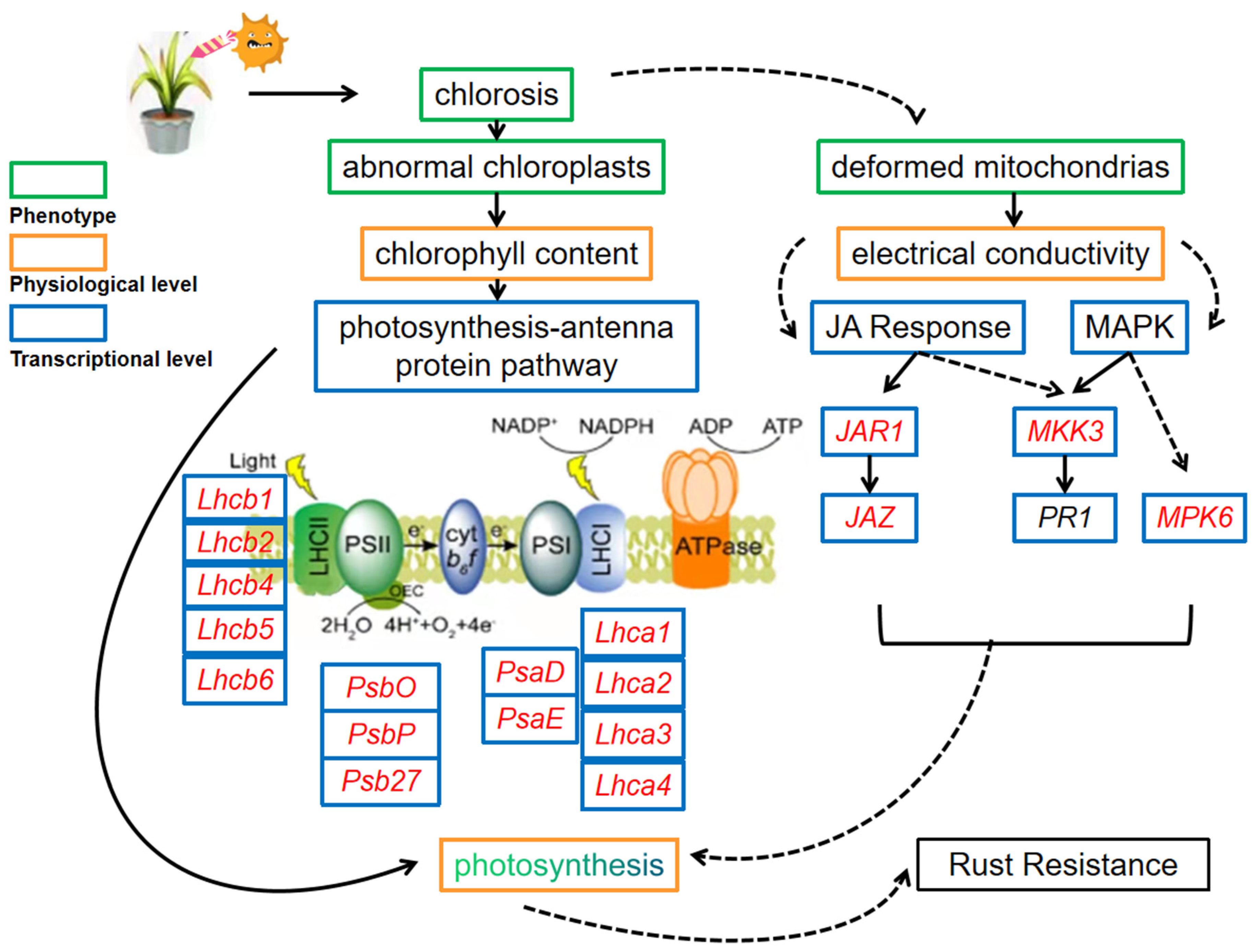
3. Discussion
3.1. Leaf Cell Structure and Physiological Changes after Inoculation with Rust Pathogen
3.2. Transcriptomic Response to Inoculation with Rust Pathogen
3.2.1. Photosynthesis Was Closely Related to the Rust Resistance of Z. japonica
3.2.2. Other Defense Systems Closely Related to the Response of Z. japonica to Pathogen Infection
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.1.1. Materials for Inoculation
4.1.2. Methods for Inoculation
4.2. Observations of the Inoculated Leaves
4.3. Determination of Electrolyte Leakage
4.4. Chlorophyll Determination
4.5. Transcriptome Sequencing and Analysis
4.6. Quantitative RT-PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Yoshiyuki, M. Effects of calcium application on the salt tolerance and sodium excretion from salt glands in zoysiagrass (Zoysia japonica). Grassl. Sci. 2019, 65, 3. [Google Scholar] [CrossRef]
- Yang, X.; Guo, H. Evaluation on trampling tolerance of two warm-season turfgrass varieties. Caoye Kexue 2018, 12, 54–62. [Google Scholar]
- He, Q.; Liu, J. Advancement in rust research on warm season turfgrass in China. Zhongguo Yesheng Zhiwu Ziyuan 2005, 24, 8–10. [Google Scholar]
- Leppik, E.E. Some Viewpoints on the Phylogeny of Rust Fungi. III. Origin of Grass Rusts. Mycologia 2018, 51, 4. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, X.; Chen, W.; Liu, T.; Zhong, S.; Huang, Q.; Ren, T.; Li, Z.; Lou, P. Resistance performance of wheat stripe rust resistance gene Yr41 and its effect on yield parameters in F2 populations under field conditions. Crop Prot. 2020, 134. [Google Scholar] [CrossRef]
- Li, M.; Li, W.; Sun, Y.; Mao, P.; Qi, X.; Wang, Y. Analysis of leaf tissue structures between rust-resistant and rust-susceptible Zoysiagrass (Zoysia japonica). Acta Physiol. Plant 2018, 40, 75. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Gangwar, O.P.; Prasad, P. Immunity to rusts in wheat: Theory, fact and practice. Indian Phytopathol. 2021, 74, 355–363. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, Y.; Ran, L.X.; Jiang, H.B. Transcriptome Analysis of the Tea Leaves (Camellia sinensis var. assamica) Infected by Tea Blister Blight. J. Tea Sci. 2020, 40, 113–124. [Google Scholar] [CrossRef]
- Dong, H.; Duan, J.; Ma, M.; Li, Z.; Li, H. Reactive oxygen species induce cyanide-resistant respiration in potato infected by Erwinia carotovora subsp. Carotovora. J. Plant Physiol. 2020, 246–247, 153132. [Google Scholar] [CrossRef]
- Liu, Q.G.; Zhang, J.Y.; Feng, M.S.; Wang, Z.Z. Effect of toxin produced by Dickeya zeae on reactive oxidative specifies (ROS) metabolism and cell ultrastructure in tobacco. Acta Ecol. Sin. 2009, 39, 262–271. [Google Scholar] [CrossRef]
- Wang, Y. Cell Biology, Reactive Oxygen Metabolism and Cell Hypersensitive Response of Poplar-Canker Pathogen Interaction. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2007. [Google Scholar]
- Kang, Z.; Wang, Y.; Huang, L.; Wei, G.; Zhao, J. Histology and Ultrastructure of Incompatible Combination between Puccinia striiformis and Wheat Cultivars with Resistance of Low Reaction Type. Sci. Agric. Sin. 2003, 36, 1026–1031. [Google Scholar]
- Dong, J.; Wang, Y.; Xian, Q.; Chen, X.; Xu, J. Transcriptome analysis reveals ethylene-mediated defense responses to Fusarium oxysporum f. sp. cucumerinum infection in Cucumis sativus L. BMC Plant Biol. 2020, 20, 1. [Google Scholar] [CrossRef]
- Ding, T.; Wang, X.; Duan, Y. Effect of disease on photosynthesis, nutrition, and nodulation of leguminous forage. Pratacult. Sci. 2020, 36, 152–160. [Google Scholar] [CrossRef]
- Yang, C.; DU, S.; Zhang, D.; Li, X.; Shi, Y.; Shao, Y.; Wang, H.; Fang, B. Method for estimating relative chlorophyll content in wheat leaves based on chlorophyll fluorescence parameters. J. Appl. Ecol. 2021, 32, 175–181. [Google Scholar] [CrossRef]
- Nan, Z.; Li, C.; Wang, Y.; Wang, Y. Lucerne common leaf spot forage quality, photosynthesis rate and field resistance. Acta Pratacult. Sin. 2001, 10, 26–34. [Google Scholar]
- Fan, Q.; Li, Y. The effect of Phoma medicaginis on the photosynthetic physiology of Medicago sativa. Acta Pratacult. Sin. 2017, 26, 112–121. [Google Scholar] [CrossRef]
- Wang, S.; Li, Q.; Wang, J.; Yan, Y. YR36/WKS1-Mediated Phosphorylation of PsbO, an Extrinsic Member of Photosystem II, Inhibits Photosynthesis and Confers Stripe Rust Resistance in Wheat. Mol. Plant 2019, 12, 1639–1650. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wellings, C.; Chen, X.; Kang, Z.; Liu, T. Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp. tritici. Mol. Plant Pathol. 2014, 15, 433–446. [Google Scholar] [CrossRef]
- Gou, J.; Li, K.; Wu, K.; Wang, X.; Lin, H.; Dario, C.; Cristobal, U.; Albor, D.; Takamufi, M.; Kentaro, I.; et al. Wheat Stripe Rust Resistance Protein WKS1 Reduces the Ability of the Thylakoid-Associated Ascorbate Peroxidase to Detoxify Reactive Oxygen Species. Plant Cell 2015, 27, 1755–1770. [Google Scholar] [CrossRef]
- Wang, M. Molecular Mechanism of ABA Pathway Response to Zoysia Japonia Being Infected by Rhizoctonia solani. Master’s Thesis, Beijing Forestry University, Beijing, China, 2016. [Google Scholar]
- Hao, Y.; Wang, T.; Wang, K. Transcriptome analysis provides insights into the mechanisms underlying wheat plant resistance to stripe rust at the adult plant stage. PLoS ONE 2016, 11, e150717. [Google Scholar] [CrossRef]
- Zhang, L.M. Identification of Zoysiagrass (Zoysia japonica Steud.) Germplasm to Rust and Screening of SSR Markers Linked with Resistant Genes. Master’s Thesis, China Agricultural University, Beijing, China, 2010. [Google Scholar]
- Gu, G. Germplasm Evaluation for Rust Resistance and Polymorphic Analysis of SSR Markers between Resistant and Susceptible Parents in Zoysiagrass (Zoysia japonica Steud.). Master’s Thesis, China Agricultural University, Beijing, China, 2013. [Google Scholar]
- Zhang, D.; Zeng, H.; Zhao, L.; Yue, J.; Qi, X.; Li, M. Isolation and Identification Rust Pathogens and the Study of Antioxidant Enzyme Activity and Gene Expression under Rust Infection in Zoysia japonica. Agriculture 2021, 11, 1200. [Google Scholar] [CrossRef]
- Park, E.; Nedo, A.; Caplan, J.; Dinesh, S. Plant-microbe interactions: Organelles and the cytoskeleton in action. New Phytol. 2018, 217, 1012–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeleneva, Y.; Sudnikova, V. Research and restocking of wheat varieties to pathogens of especially dangerous fungal diseases. E3S Web Conf. 2021, 254, 01008. [Google Scholar] [CrossRef]
- Cao, Y. Study on Some Physiological Characters of Resistance to Rhizomania in Sugar Beet. Master’s Thesis, Inner Mongolia Agricultural University, Huhhot, China, 2006. [Google Scholar]
- Laliberte, J.; Sanfacon, H. Cellular Remodelling During Plant Virus Infection. Annu. Rev. Phytopathol. 2010, 48, 69–91. [Google Scholar] [CrossRef]
- Ou, Z. The relationship between chlorophyll content and plant disease resistance. Anhui Agric. Sci. Bull. 2007, 13, 134–135. [Google Scholar] [CrossRef]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Su, X.; Cao, P.; Liu, X.; Chang, W.; Li, M.; Zhang, X.; Liu, Z. Structure of spinach photosystem II-LHCII supercomplex at 3.2 Å resolution. Nature 2016, 534, 69–74. [Google Scholar] [CrossRef]
- Schubert, H.; Blumenthal, R.; Cheng, X. Many paths to methyltransfer: A chronicle of convergence. Trends Biochem. Sci. 2003, 28, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Moy, P.; Qutob, D.; Chapman, B.; Atkinson, I.; Gijzen, M. Patterns of gene expression upon infection of soybean plants by Phytophthora sojae. Mol. Plant-Microbe Interact. 2004, 17, 1051–1062. [Google Scholar] [CrossRef] [Green Version]
- Arianne, T.; Parsa, H.; Nadim, W.; Li, S.; Benjamin, F. Transcriptome analysis of a compatible response by Glycine max to Phakopsora pachyrhizi infection. Plant Sci. 2010, 179, 183–193. [Google Scholar] [CrossRef]
- Lorrain, C.; Petre, B.; Duplessis, S. Show me the way: Rust effector targets in heterologous plant systems. Curr. Opin. Microbiol. 2018, 46, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, Y.; Cai, H.; Guo, M.; Chai, M.; She, Z.; Ye, L.; Cheng, Y.; Wang, B.; Qin, Y. The bZIP Transcription Factor GmbZIP15 Negatively Regulates Salt- and Drought-Stress Responses in Soybean. Int. J. Mol. Sci. 2020, 21, 7778. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Fang, W.; Yang, X.; Xie, D.; Li, W.; Tang, Z.; Li, D. Cloning and Analysis of Upland Cotton (Gossypium hirsutum) Dirigent-like Gene (GhDIR). Acta Agric. Boreali-Sin. 2011, 26, 29–33. [Google Scholar]
- Wang, J. Transformation of Herbicide and Blast Resistant Genes to Rice and their Inheritance Behavior. Ph.D. Thesis, Northeast Normal University, Changchun, China, 2002. [Google Scholar]
- Zhao, X. Preliminary Study on the Oligochitosan Induced Plant Resistant and Its Mechanisms. Ph.D. Thesis, Chinese Academy of Sciences, Dalian, China, 2006. [Google Scholar]
- Zhao, T. The Role of NbPIF1 in Harpin and SsCut-Induced Plant Immunity. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2015. [Google Scholar]
- Fan, S.X. Multi-Location Evaluation of Zoysiagrass (Zoysia japonica Steud.) Resistant Germplasm and Screening of SSR Markers with Resistant Genes. Master’s Thesis, China Agricultural University, Beijing, China, 2012. [Google Scholar]
- Nyassé, S.; Despréaux, D.; Cilas, C. Validity of a leaf inoculation test to assess the resistance to Phytophthora megakarya in a cocoa (Theobroma cacao L.) diallel mating design. Euphytica 2002, 123, 395–399. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–593. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combinations | All | Up | Down | Threshold |
|---|---|---|---|---|
| RR1 vs. RS1 | 5371 | 2709 | 2662 | DESeq2 padj < 0.05 |
| RR1 vs. RR2 | 11,234 | 5382 | 5852 | DESeq2 padj < 0.05 |
| RS1 vs. RS2 | 11,957 | 5900 | 6057 | DESeq2 padj < 0.05 |
| RR2 vs. RS2 | 3174 | 1427 | 1747 | DESeq2 padj < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Tang, J.; Wei, K.; Jia, S.; Jiang, Y.; Cai, H.; Mao, P.; Li, M. Physiological and Molecular Responses of Zoysia japonica to Rust Infection. Int. J. Mol. Sci. 2022, 23, 4185. https://doi.org/10.3390/ijms23084185
Zhang D, Tang J, Wei K, Jia S, Jiang Y, Cai H, Mao P, Li M. Physiological and Molecular Responses of Zoysia japonica to Rust Infection. International Journal of Molecular Sciences. 2022; 23(8):4185. https://doi.org/10.3390/ijms23084185
Chicago/Turabian StyleZhang, Di, Jun Tang, Kai Wei, Shangang Jia, Yiwei Jiang, Hongwei Cai, Peisheng Mao, and Manli Li. 2022. "Physiological and Molecular Responses of Zoysia japonica to Rust Infection" International Journal of Molecular Sciences 23, no. 8: 4185. https://doi.org/10.3390/ijms23084185