
Released ATP Mediates Spermatozoa Chemotaxis Promoted by Uterus-Derived Factor (UDF) in Ascaris suum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
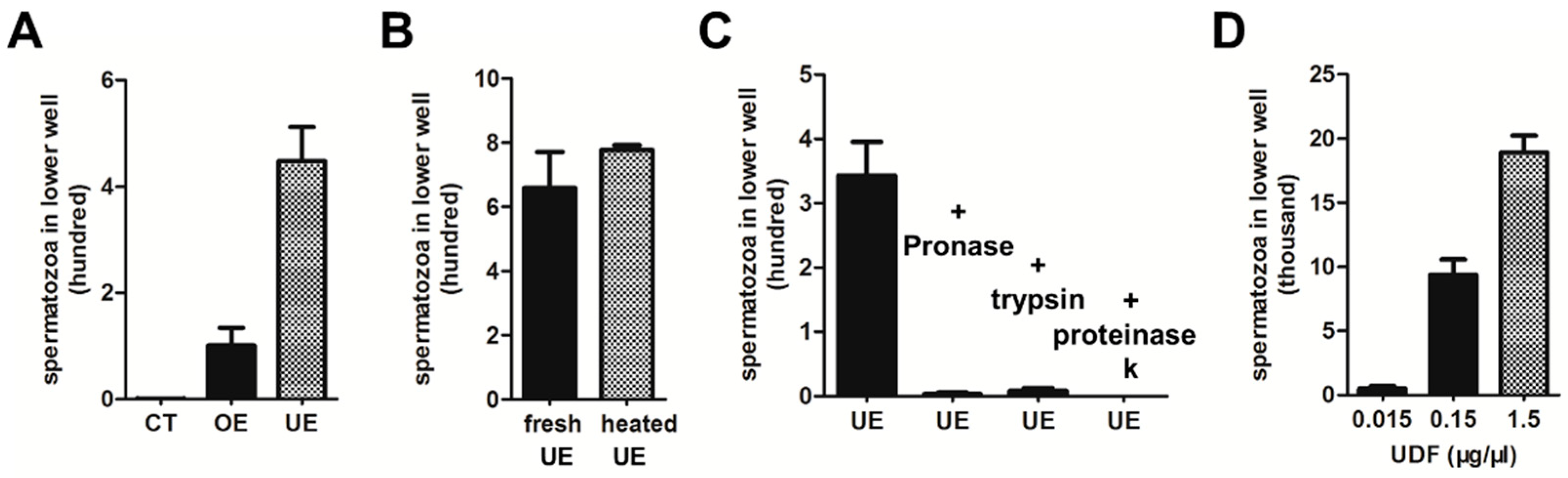
2.1. Characterization of Uterus-Derived Factor (UDF)
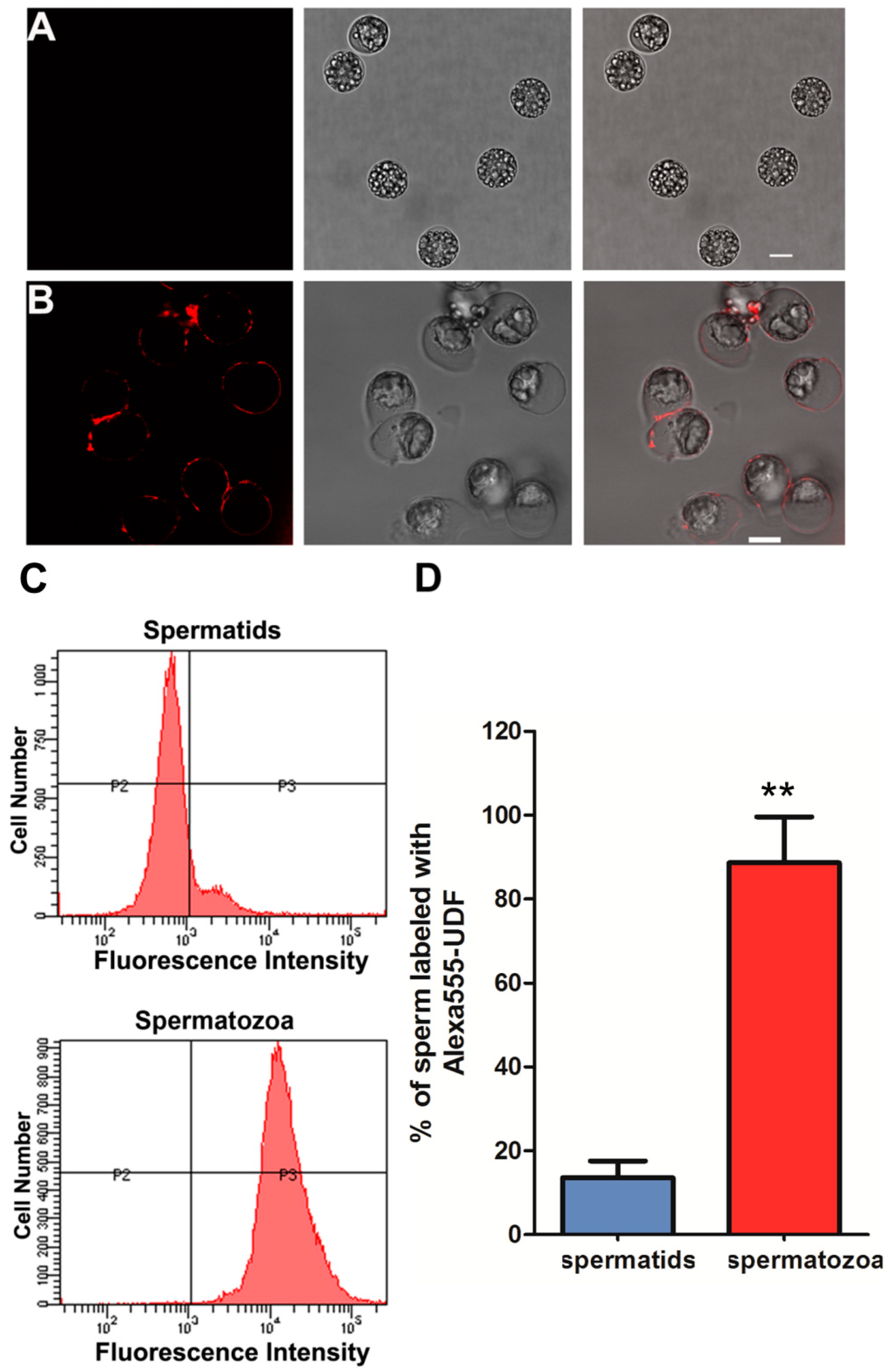
2.2. UDF Binds Spermatozoa but Not Spermatids
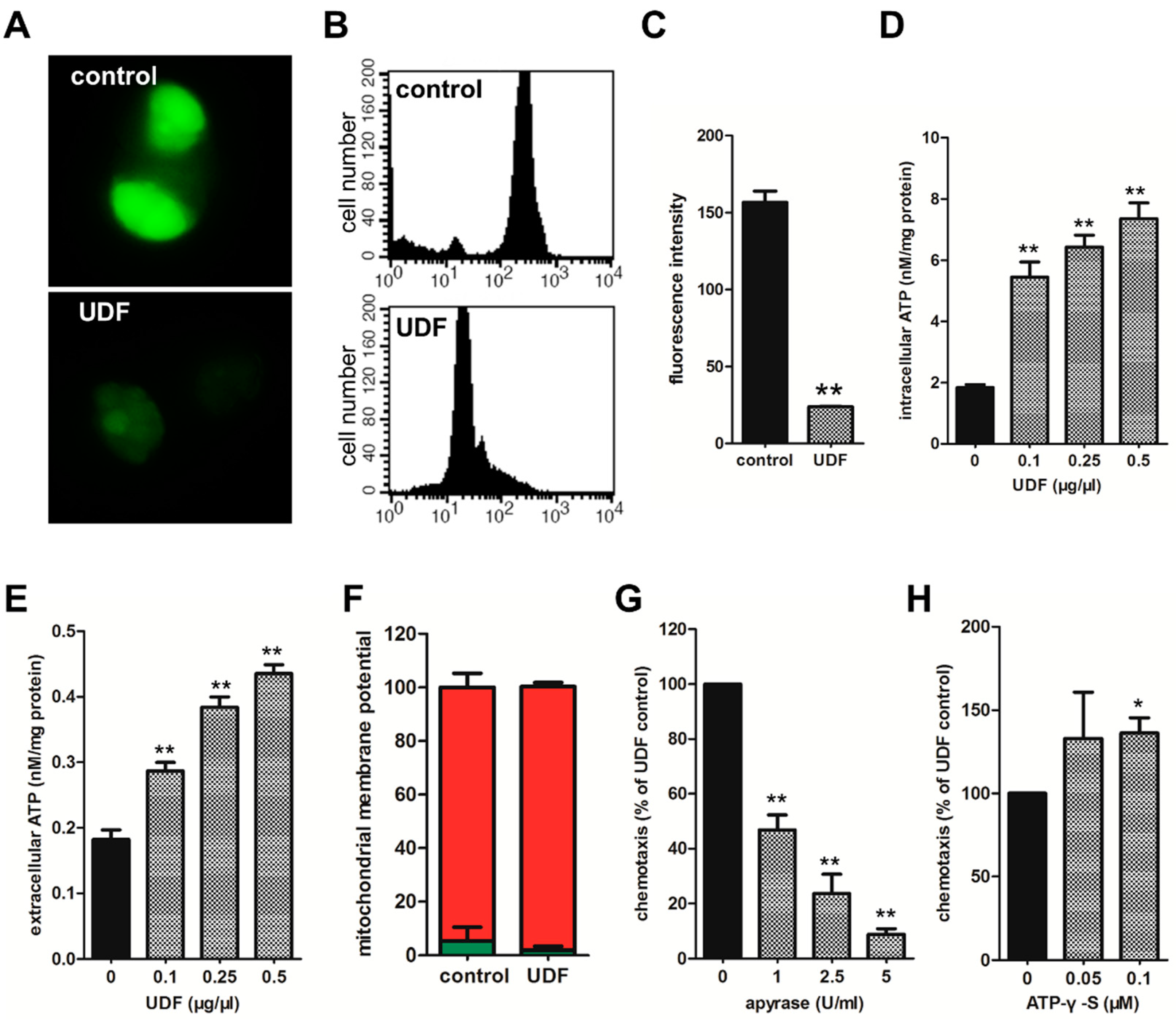
2.3. UDF Induces ATP to Release from Spermatozoa and Released ATP Is Essential for Sperm Chemotaxis Promoted by UDF
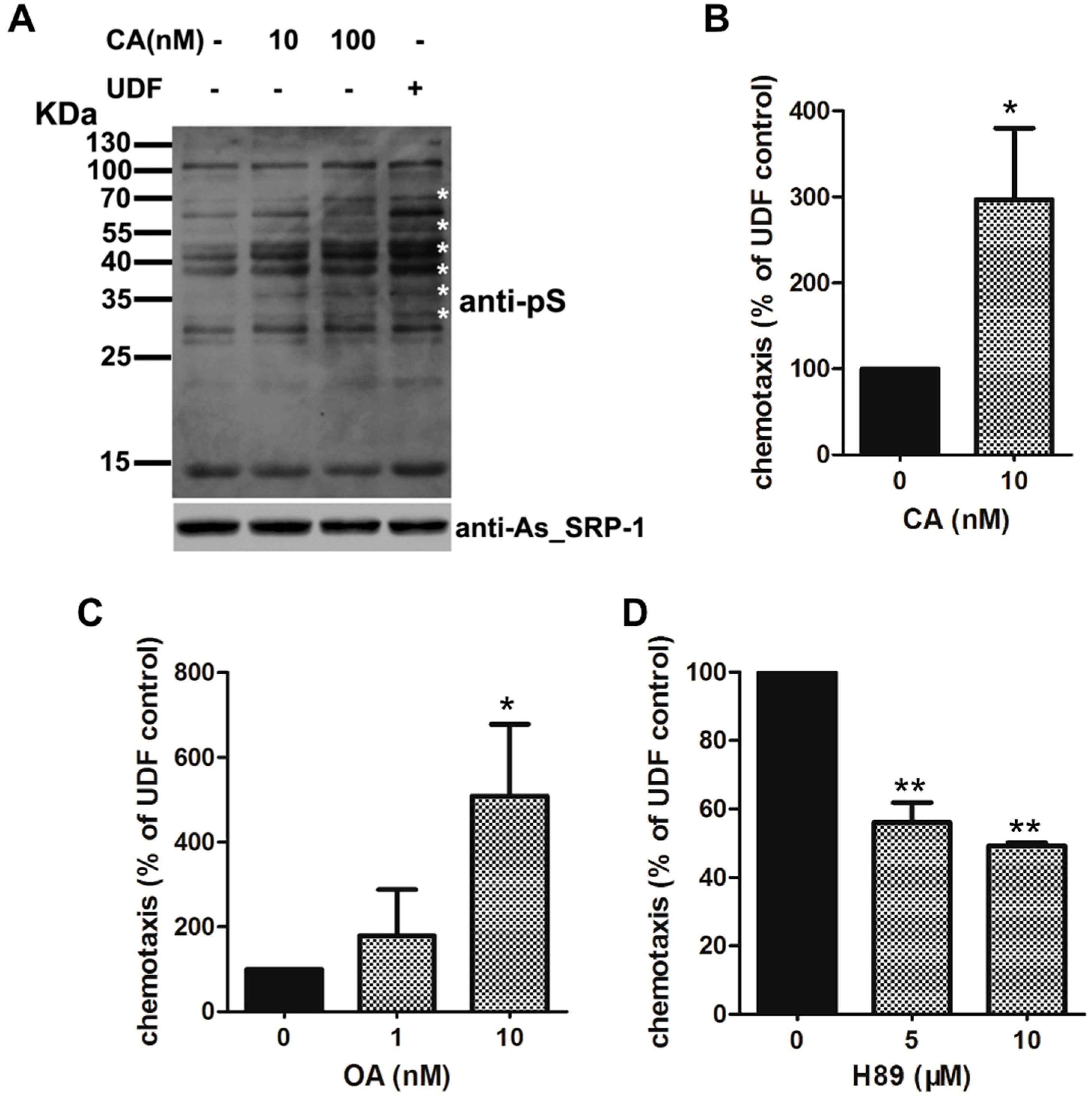
2.4. UDF Elevates Protein Serine Phosphorylation Level of Spermatozoa
3. Discussion
4. Materials and Methods
4.1. Preparation and Purification of UDF
4.2. UDF Binding Assays
4.3. Transwell Chemotaxis Assays
4.4. Intracellular and Extracellular ATP Measurements
4.5. Quinacrine and FM 1-43 Labeling
4.6. Mitochondrial Membrane Potential Measurement
4.7. Immunoblot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Larsson, L.; Laska, M. Ultra-high olfactory sensitivity for the human sperm-attractant aromatic aldehyde bourgeonal in CD-1 mice. Neurosci. Res. 2011, 71, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Y.H.; Guasto, J.S.; Zimmer, R.K.; Stocker, R.; Riffell, J.A. Sperm chemotaxis promotes individual fertilization success in sea urchins. J. Exp. Biol. 2016, 219, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, E.; Kaupp, U.B. Sperm Chemotaxis: A Primer. Ann. N. Y. Acad. Sci. 2005, 106, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Hansbrough, J.; Garbers, D. Speract. Purification and characterization of a peptide associated with eggs that activates spermatozoa. J. Biol. Chem. 1981, 256, 1447–1452. [Google Scholar] [CrossRef]
- Kaupp, U.; Hildebrand, E.; Weyand, I. Sperm chemotaxis in marine invertebrates—molecules and mechanisms. J. Cell. Physiol. 2006, 208, 487–494. [Google Scholar] [CrossRef]
- Xiang, X.; Kittelson, A.; Olson, J.; Bieber, A.; Chandler, D. Allurin, a 21 kD sperm chemoattractant, is rapidly released from the outermost jelly layer of theXenopus egg by diffusion and medium convection. Mol. Reprod. Dev. 2004, 70, 344–360. [Google Scholar] [CrossRef]
- Burnett, L.; Anderson, D.M.; Rawls, A.; Bieber, A.L.; Chandler, D.E. Mouse sperm exhibit chemotaxis to allurin, a truncated member of the cysteine-rich secretory protein family. Dev. Biol. 2011, 360, 318–328. [Google Scholar] [CrossRef]
- Hoang, H.D.; Prasain, J.K.; Dorand, D.; Miller, M.A. A Heterogeneous Mixture of F-Series Prostaglandins Promotes Sperm Guidance in the Caenorhabditis elegans Reproductive Tract. PLoS Genet. 2013, 9, e1003271. [Google Scholar] [CrossRef]
- Eisenbach, M.; Giojalas, L. Sperm guidance in mammals—An unpaved road to the egg. Nat. Rev. Mol. Cell Biol. 2006, 7, 276–285. [Google Scholar] [CrossRef]
- Kubagawa, H.M.; Watts, J.L.; Corrigan, C.; Edmonds, J.W.; Sztul, E.; Browse, J.; Miller, M.A. Oocyte signals derived from poly-unsaturated fatty acids control sperm recruitment in vivo. Nat. Cell Biol. 2006, 8, 1143–1148. [Google Scholar] [CrossRef]
- Tiwary, E.; Hu, M.; Parasin, J.K. Sperm-Guiding Unconventional Prostaglandins in C. elegans: Synthesis and Signaling. Metabolites 2021, 11, 853. [Google Scholar] [CrossRef] [PubMed]
- Pichlo, M.; Bungert-Plümke, S.; Weyand, I.; Seifert, R.; Bönigk, W.; Strünker, T.; Kashikar, D.; Goodwin, N.; Müller, A.; Körschen, H.G.; et al. High density and ligand affinity confer ultrasensitive signal detection by a guanylyl cyclase chemoreceptor. J. Cell Biol. 2014, 206, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nature 2011, 471, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Jansen, V.; Jikeli, J.F.; Hamzeh, H.; Alvarez, L.; Dombrowski, M.; Balbach, M.; Strünker, T.; Seifert, R.; Kaupp, U.B.; et al. A novel biosensor to study cAMP dynamics in cilia and flagella. eLife 2016, 5, e14052. [Google Scholar] [CrossRef]
- Kirichok, Y.; Lishko, P.V. Rediscovering sperm ion channels with the patch-clamp technique. Mol. Hum. Reprod. 2011, 17, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Shiba, K.; Sakamoto, A.; Ikenaga, J.; Matsunaga, S.; Inaba, K.; Yoshida, M. Ca2+ efflux via plasma membrane Ca2+-ATPase mediates chemotaxis in ascidian sperm. Sci. Rep. 2018, 8, 16622. [Google Scholar] [CrossRef]
- Ortiz-Ramírez, C.; Michard, E.; Simon, A.A.; Damineli, D.S.C.; Hernández-Coronado, M.; Becker, J.D.; Feijó, J.A. GLUTAMATE RECEPTOR-LIKE channels are essential for chemotaxis and reproduction in mosses. Nature 2017, 549, 91. [Google Scholar] [CrossRef]
- Strunker, T.; Goodwin, N.; Brenker, C.; Kashikar, N.D.; Weyand, I.; Seifert, R.; Kaupp, U.B. The CatSper channel mediates pro-gesterone-induced Ca2+ influx in human sperm. Nature 2011, 471, 382–386. [Google Scholar] [CrossRef]
- Mizuno, K.; Shiba, K.; Okai, M.; Takahashi, Y.; Shitaka, Y.; Oiwa, K.; Tanokura, M.; Inaba, K. Calaxin drives sperm chemotaxis by Ca 2+ -mediated direct modulation of a dynein motor. Proc. Natl. Acad. Sci. USA 2012, 109, 20497–20502. [Google Scholar] [CrossRef]
- Alvarez, L.; Dai, L.; Friedrich, B.M.; Kashikar, N.D.; Gregor, I.; Pascal, R.; Kaupp, U.B. The rate of change in Ca2+ concentration controls sperm chemotaxis. J. Cell Biol. 2012, 196, 653–663. [Google Scholar] [CrossRef]
- Kaupp, U.B.; Solzin, J.; Hildebrand, E.; Brown, J.E.; Helbig, A.; Hagen, V.; Beyermann, M.; Pampaloni, F.; Weyand, I. The signal flow and motor response controling chemotaxis of sea urchin sperm. Nat. Cell Biol. 2003, 5, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; Wang, H.-K.; Ye, C.-Q.; Ge, W.; Chen, Y.; Jiang, Z.-L.; Wu, C.-P.; Poo, M.-M.; Duan, S. ATP Released by Astrocytes Mediates Glutamatergic Activity-Dependent Heterosynaptic Suppression. Neuron 2003, 40, 971–982. [Google Scholar] [CrossRef]
- Dou, Y.; Wu, H.-J.; Li, H.-Q.; Qin, S.; Wang, Y.-E.; Li, J.; Lou, H.-F.; Chen, Z.; Li, X.-M.; Luo, Q.-M.; et al. Microglial migration mediated by ATP-induced ATP release from lysosomes. Cell Res. 2012, 22, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Pascual, O.; Casper, K.B.; Kubera, C.; Zhang, J.; Revilla-Sanchez, R.; Sul, J.-Y.; Takano, H.; Moss, S.J.; McCarthy, K.; Haydon, P.G. Astrocytic Purinergic Signaling Coordinates Synaptic Networks. Science 2005, 310, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Hirsh, M.I.; Chen, Y.; Woehrle, T.; Karande, A.A.; Junger, W.G. ATP release and autocrine signaling through P2X4 receptors regulate γδ T cell activation. J. Leukoc. Biol. 2012, 92, 787–794. [Google Scholar] [CrossRef]
- Junger, W.G. Immune cell regulation by autocrine purinergic signalling. Nat. Rev. Immunol. 2011, 11, 201–212. [Google Scholar] [CrossRef]
- Bao, Y.; Chen, Y.; Ledderose, C.; Li, L.; Junger, W.G. Pannexin 1 Channels Link Chemoattractant Receptor Signaling to Local Excitation and Global Inhibition Responses at the Front and Back of Polarized Neutrophils. J. Biol. Chem. 2013, 288, 22650–22657. [Google Scholar] [CrossRef]
- Chen, Y.; Corriden, R.; Inoue, Y.; Yip, L.; Hashiguchi, N.; Zinkernagel, A.; Nizet, V.; Insel, P.A.; Junger, W.G. ATP Release Guides Neutrophil Chemotaxis via P2Y2 and A3 Receptors. Science 2006, 314, 1792–1795. [Google Scholar] [CrossRef]
- Alkayed, F.; Kashimata, M.; Koyama, N.; Hayashi, T.; Tamura, Y.; Azuma, Y. P2Y11 Purinoceptor Mediates the ATP-Enhanced Chemotactic Response of Rat Neutrophils. J. Pharmacol. Sci. 2012, 120, 288–295. [Google Scholar] [CrossRef]
- Caballero-Campo, P.; Buffone, M.G.; Benencia, F.; Conejo-Garcia, J.; Rinaudo, P.F.; Gerton, G.L. A Role for the Chemokine Receptor CCR6 in Mammalian Sperm Motility and Chemotaxis. J. Cell. Physiol. 2013, 229, 68–78. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, W.; Zhang, P.; Chi, H.; Zhang, M.-J.; Song, C.-Q.; Ma, X.; Shang, Y.; Wang, B.; Hu, Y.; et al. Nematode sperm maturation triggered by protease involves sperm-secreted serine protease inhibitor (Serpin). Proc. Natl. Acad. Sci. USA 2012, 109, 1542–1547. [Google Scholar] [CrossRef] [PubMed]
- Figueiras-Fierro, D.; Acevedo, J.J.; Martínez-López, P.; Escoffier, J.; Sepúlveda, F.V.; Balderas, E.; Orta, G.; Visconti, P.E.; Dar-szon, A. Electrophysiological evidence for the presence of cystic fibrosis transmembrane conductance regulator (CFTR) in mouse sperm. J. Cell. Physiol. 2013, 228, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Loiola, E.C.; Ventura, A.L.M. Release of ATP from avian Müller glia cells in culture. Neurochem. Int. 2010, 58, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Inaba, K. Lipid rafts function in Ca2+ signaling responsible for activation of sperm motility and chemotaxis in the as-cidian Ciona intestinalis. Mol. Reprod. Dev. 2011, 78, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Fabbro, A.; Skorinkin, A.; Grandolfo, M.; Nistri, A.; Giniatullin, R. Quantal release of ATP from clusters of PC12 cells. J. Physiol. 2004, 560, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Isfort, K.; Ebert, F.; Bornhorst, J.; Sargin, S.; Kardakaris, R.; Pasparakis, M.; Bähler, M.; Schwerdtle, T.; Schwab, A.; Hanley, P.J. Real-time Imaging Reveals That P2Y2 and P2Y12 Receptor Agonists Are Not Chemoattractants and Macrophage Chemotaxis to Complement C5a Is Phosphatidylinositol 3-Kinase (PI3K)- and p38 Mitogen-activated Protein Kinase (MAPK)-independent*. J. Biol. Chem. 2011, 286, 44776–44787. [Google Scholar] [CrossRef]
- Yi, K.; Wang, X.; Emmett, M.R.; Marshall, A.G.; Stewart, M.; Roberts, T.M. Dephosphorylation of Major Sperm Protein (MSP) Fiber Protein 3 by Protein Phosphatase 2A during Cell Body Retraction in the MSP-based Amoeboid Motility ofAscarisSperm. Mol. Biol. Cell 2009, 20, 3200–3208. [Google Scholar] [CrossRef]
- Buzzi, N.; Bilbao, P.S.; Boland, R.; de Boland, A.R. Extracellular ATP activates MAP kinase cascades through a P2Y purinergic receptor in the human intestinal Caco-2 cell line. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2009, 1790, 1651–1659. [Google Scholar] [CrossRef]
- Nasu-Tada, K.; Koizumi, S.; Inoue, K. Involvement of β1 integrin in microglial chemotaxis and proliferation on fibronectin: Different regulations by ADP through PKA. Glia 2005, 52, 98–107. [Google Scholar] [CrossRef]
- Teves, M.E.; Guidobaldi, H.A.; Uñates, D.R.; Sanchez, R.; Miska, W.; Publicover, S.J.; Garcia, A.A.M.; Giojalas, L.C. Molecular Mechanism for Human Sperm Chemotaxis Mediated by Progesterone. PLoS ONE 2009, 4, e8211. [Google Scholar] [CrossRef]
- Burton, K.A.; McKnight, G.S. PKA, Germ Cells, and Fertility. Physiology 2007, 22, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Shariatmadari, R.; Sipila, P.; Vierula, M.; Tornquist, K.; Huhtaniemi, I.; Poutanen, M. Adenosine Triphosphate Induces Ca2+ Signal in Epithelial Cells of the Mouse Caput Epididymis Through Activation of P2X and P2Y Purinergic Receptors 1. Biol. Reprod. 2003, 68, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Luria, A.; Rubinstein, S.; Lax, Y.; Breitbart, H. Extracellular adenosine triphosphate stimulates acrosomal exocytosis in bovine spermatozoa via P2 purinoceptor. Biol. Reprod. 2002, 66, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Foresta, C.; Rossato, M.; Di Virgilio, F. Extracellular ATP is a trigger for the acrosome reaction in human spermatozoa. J. Biol. Chem. 1992, 267, 19443–19447. [Google Scholar] [CrossRef]
- Honda, S.; Sasaki, Y.; Ohsawa, K.; Imai, Y.; Nakamura, Y.; Inoue, K.; Kohsaka, S. Extracellular ATP or ADP Induce Chemotaxis of Cultured Microglia through Gi/o-Coupled P2Y Receptors. J. Neurosci. 2001, 21, 1975–1982. [Google Scholar] [CrossRef]
- Yi, K.; Buttery, S.M.; Stewart, M.; Roberts, T.M. A Ser/Thr Kinase Required for Membrane-associated Assembly of the Major Sperm Protein Motility Apparatus in the Amoeboid Sperm of Ascaris. Mol. Biol. Cell 2007, 18, 1816–1825. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Cain, G. In vitro activation and behavior of the ameboid sperm of Ascaris suum (Nematoda). Cell Tissue Res. 1979, 200, 273–284. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; He, R.; Zhang, Q.; Shan, J.; Zhao, Y.; Wang, X. Released ATP Mediates Spermatozoa Chemotaxis Promoted by Uterus-Derived Factor (UDF) in Ascaris suum. Int. J. Mol. Sci. 2022, 23, 4069. https://doi.org/10.3390/ijms23074069
Wang Q, He R, Zhang Q, Shan J, Zhao Y, Wang X. Released ATP Mediates Spermatozoa Chemotaxis Promoted by Uterus-Derived Factor (UDF) in Ascaris suum. International Journal of Molecular Sciences. 2022; 23(7):4069. https://doi.org/10.3390/ijms23074069
Chicago/Turabian StyleWang, Qiushi, Ruijun He, Qi Zhang, Jin Shan, Yanmei Zhao, and Xia Wang. 2022. "Released ATP Mediates Spermatozoa Chemotaxis Promoted by Uterus-Derived Factor (UDF) in Ascaris suum" International Journal of Molecular Sciences 23, no. 7: 4069. https://doi.org/10.3390/ijms23074069