
The Growth, Lipid Accumulation and Fatty Acid Profile Analysis by Abscisic Acid and Indol-3-Acetic Acid Induced in Chlorella sp. FACHB-8


Abstract
:1. Introduction
2. Results
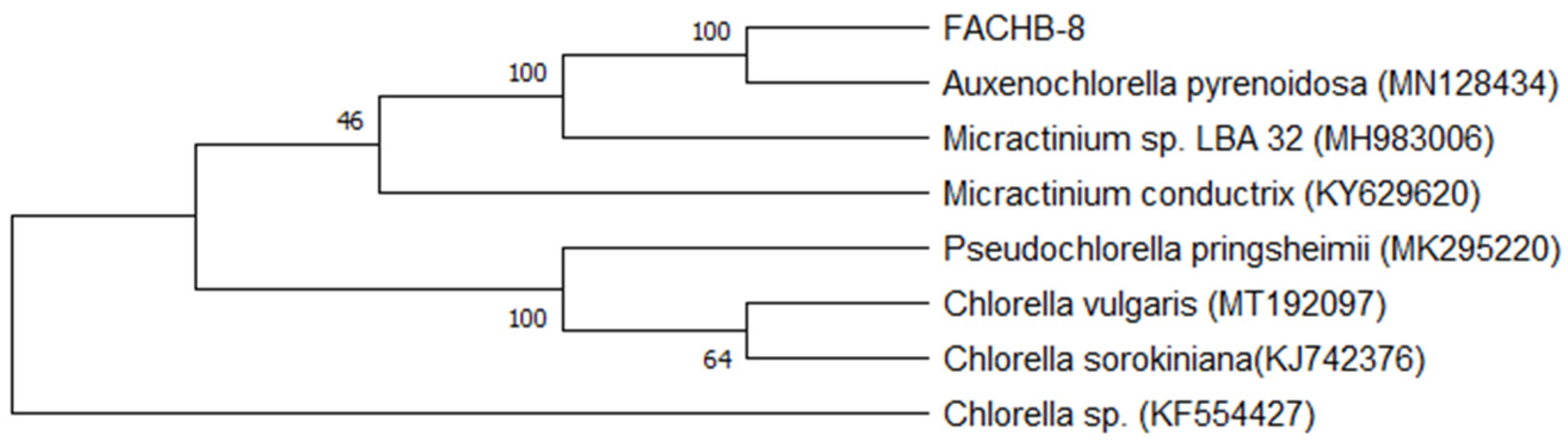
2.1. The Taxonomic Assignment of FACHB-8
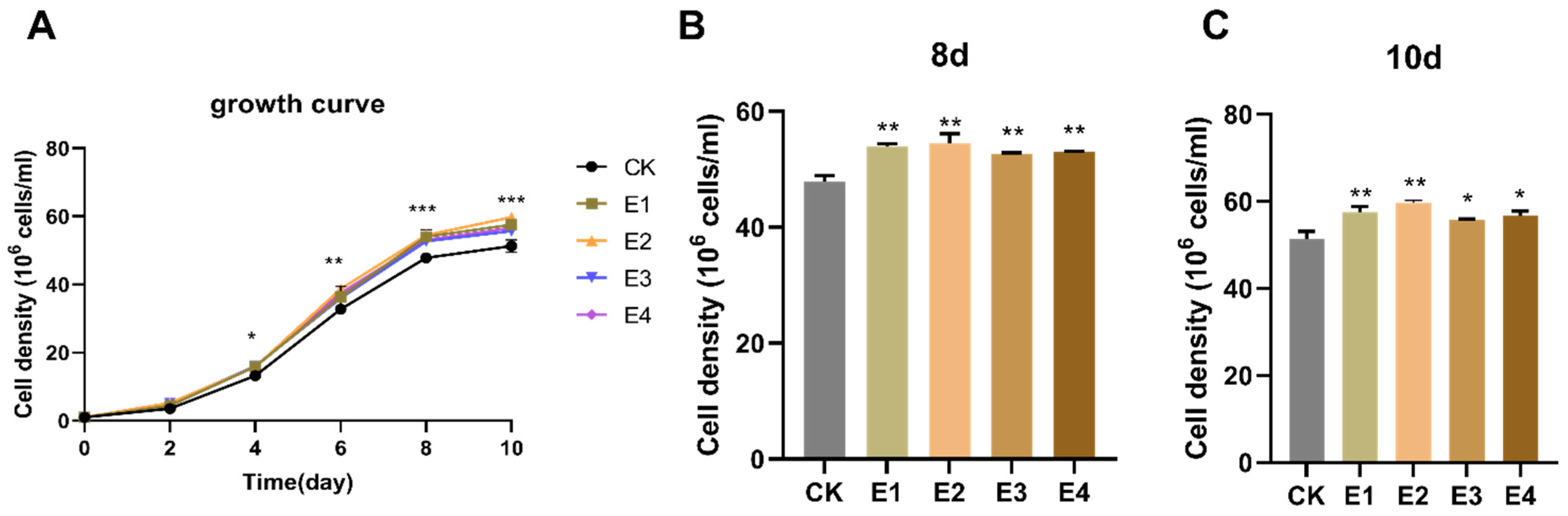
2.2. The Hormone Effect on Growth in Chlorella sp. FACHB-8
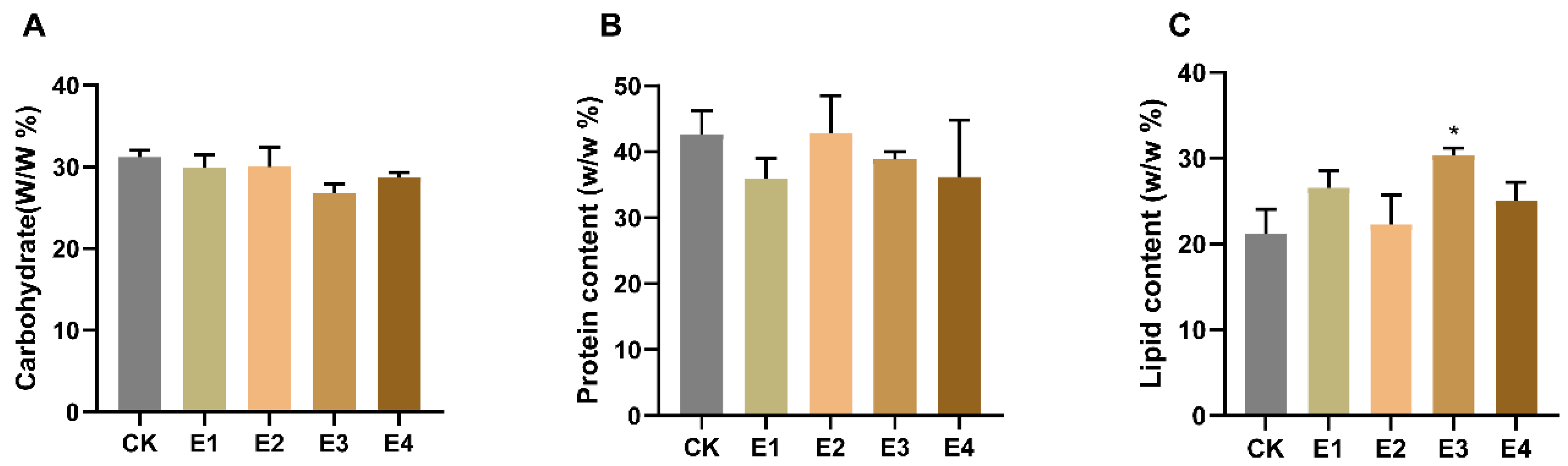
2.3. The Hormone Effect on Carbohydrate, Protein, and Lipid Accumulation
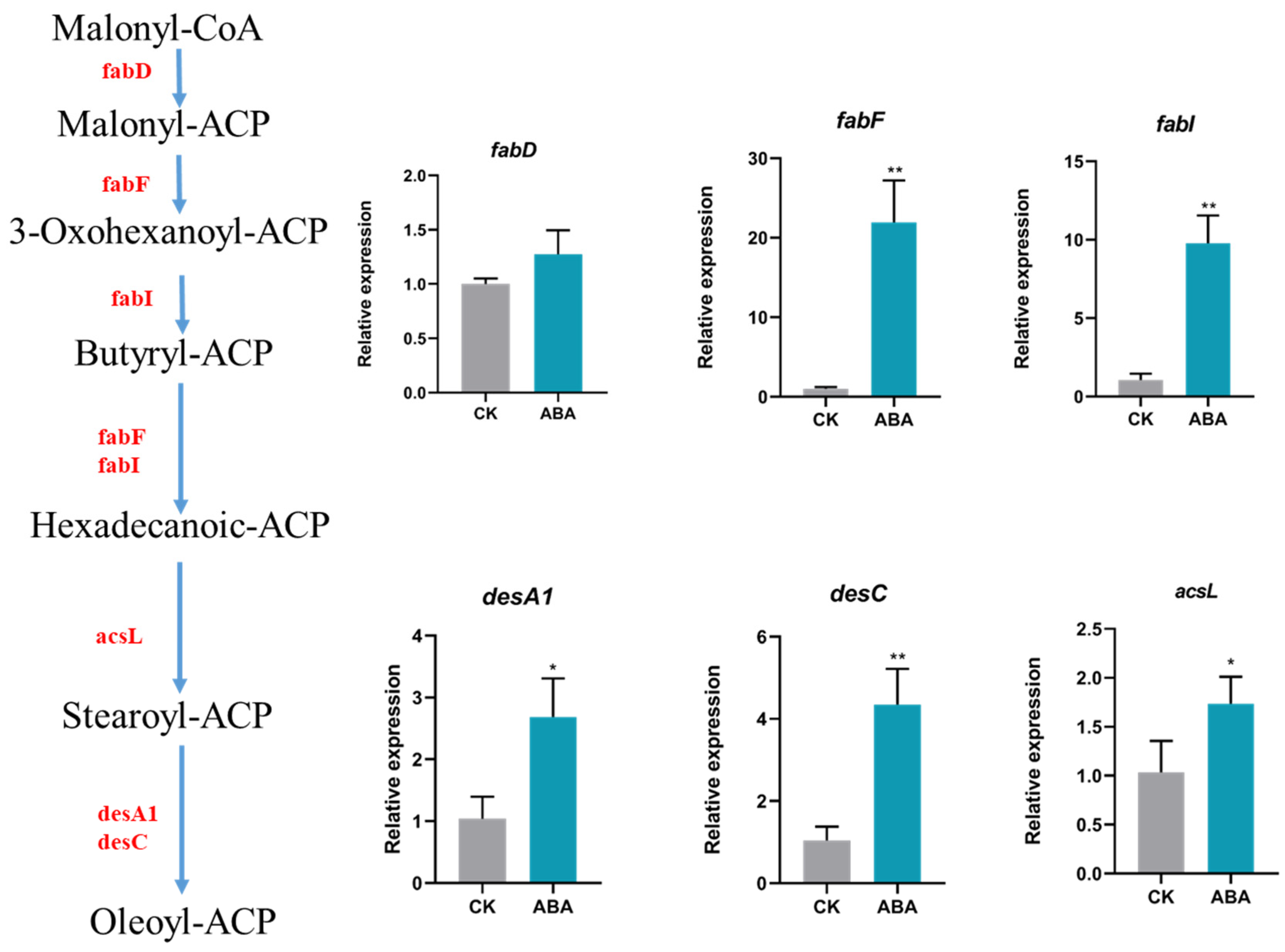
2.4. The Hormone Effect on Fatty Acid Composition
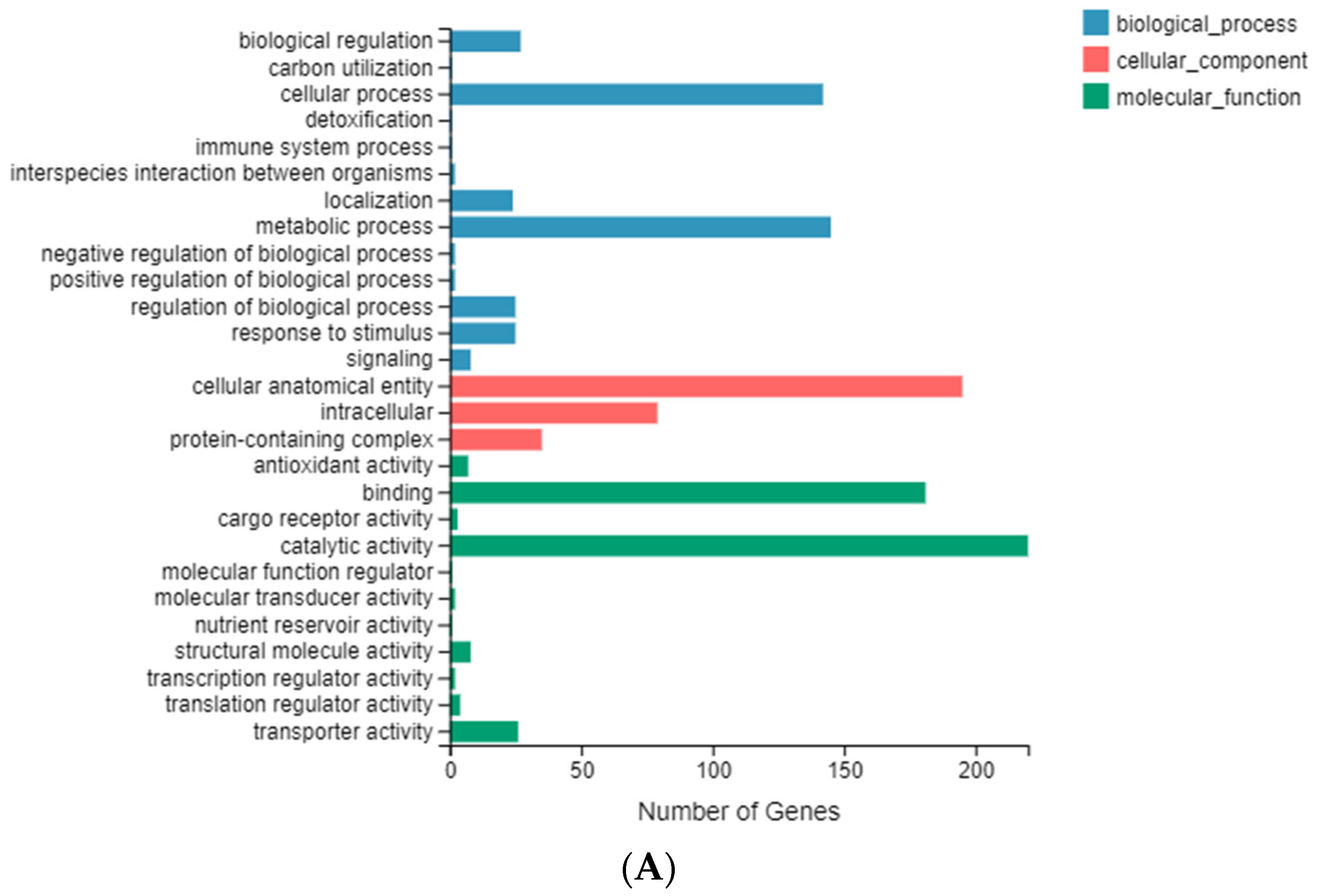
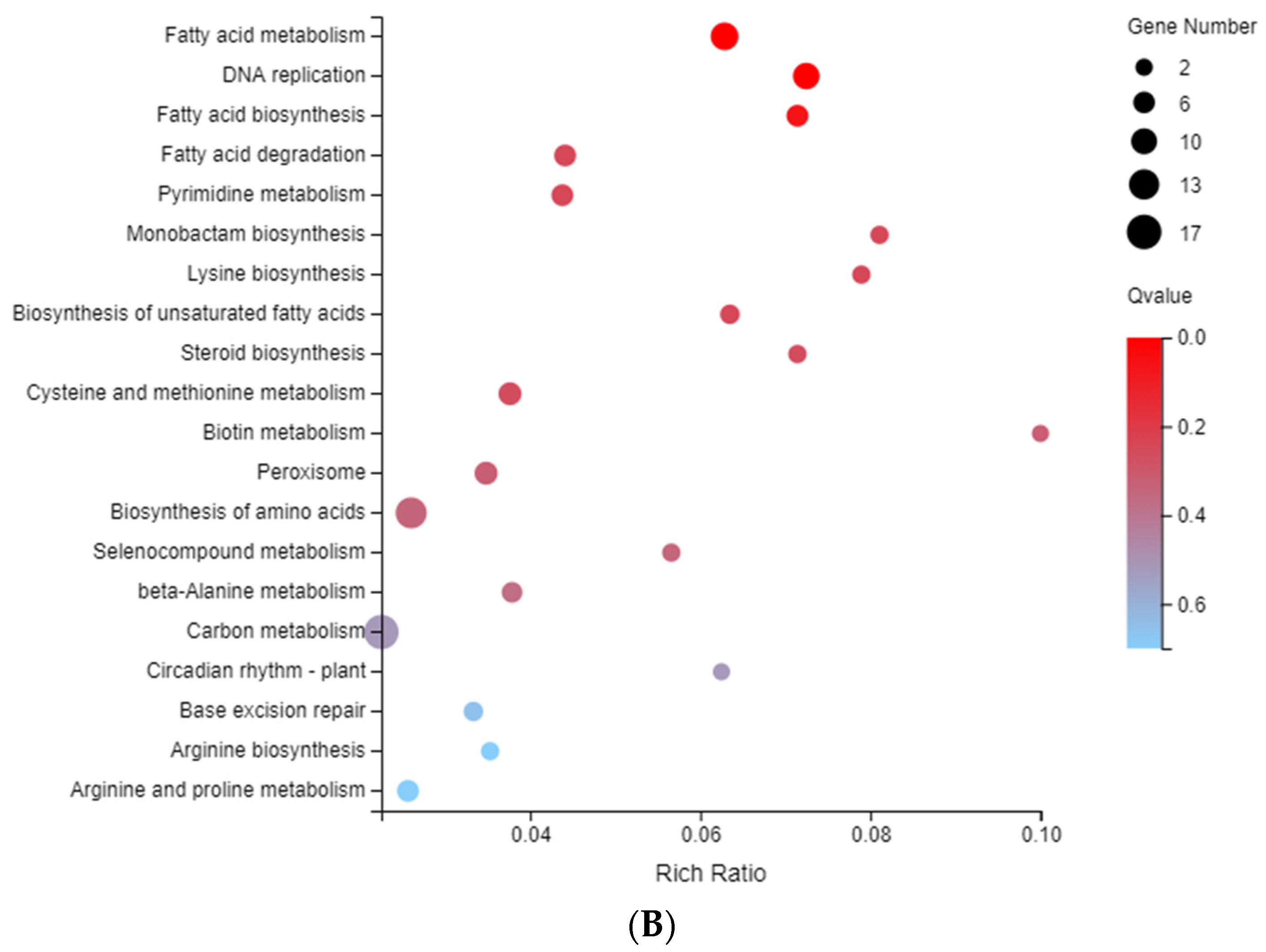
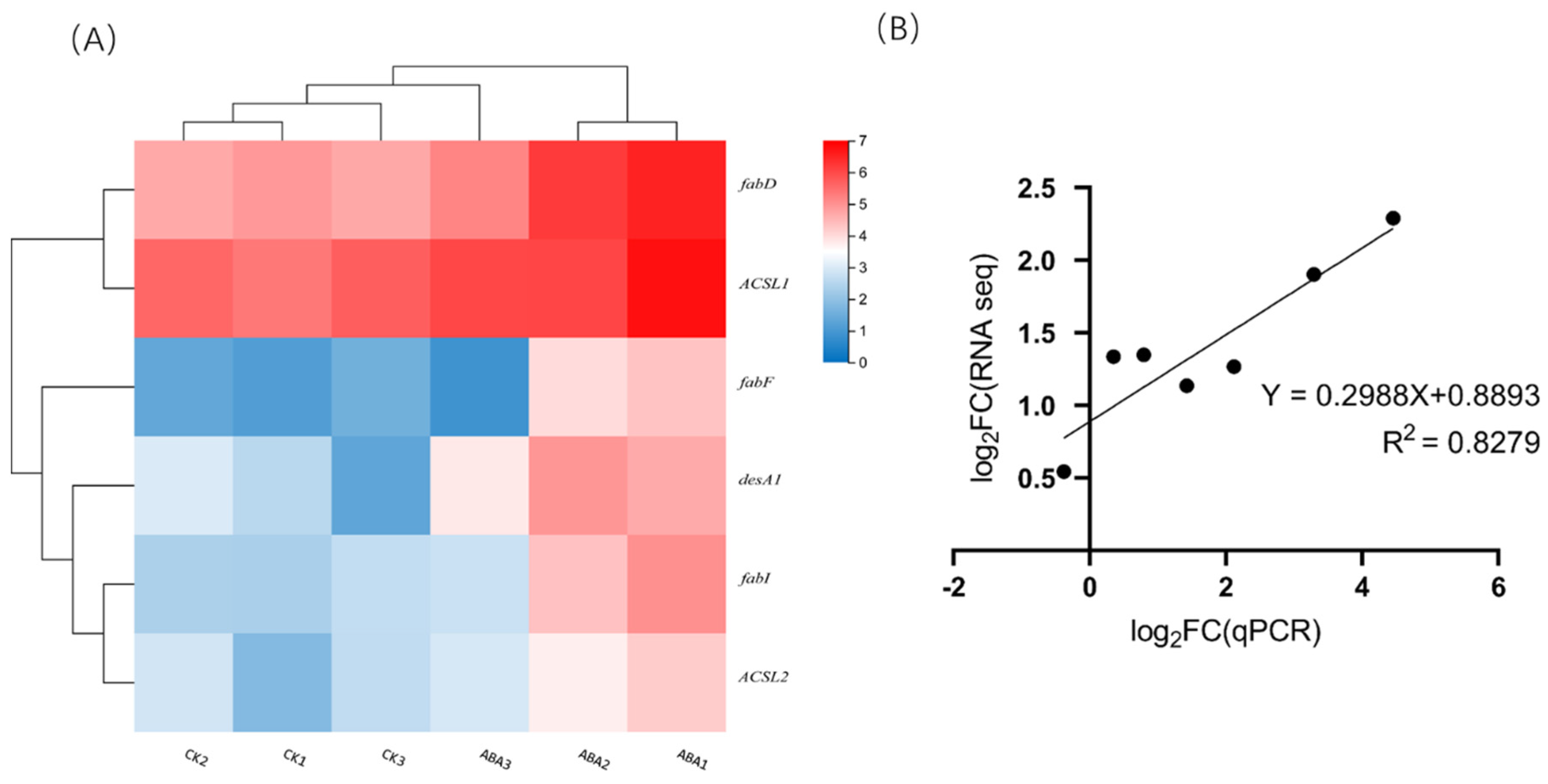
2.5. Enrichment and Identification of Differentially Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Microalgae Strain and Culture Conditions
4.2. Hormone Treatments
4.3. Analyses of Microalgae Growth, Protein and Carbohydrate Contents
4.4. Total Lipid and Fatty Acid Profile Determination
4.5. RNA-Seq
4.6. De Novo Assembly and Transcriptome Analysis
4.7. Taxonomic Assignment of FACHB-8 Strain
4.8. qRT–PCR Confirmation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duffy, J.E.; Canuel, E.A.; Adey, W.; Swaddle, J.P. Biofuels: Algae. Science 2009, 326, 1345. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K. Fatty acids of microalgae: Diversity and applications. Rev. Environ. Sci. Biotechnol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Deshmukh, S.; Kumar, R.; Bala, K. Microalgae biodiesel: A review on oil extraction, fatty acid composition, properties and effect on engine performance and emissions. Fuel Process. Technol. 2019, 191, 232–247. [Google Scholar] [CrossRef]
- Thomas, J.; Thomas, C.J.; Radcliffe, J.; Itsiopoulos, C. Omega-3 Fatty Acids in Early Prevention of Inflammatory Neurodegenerative Disease: A Focus on Alzheimer′s Disease. Biomed. Res. Int. 2015, 2015, 172801. [Google Scholar] [CrossRef] [Green Version]
- Wysoczański, T.; Sokoła-Wysoczańska, E.; Pękala, J.; Lochyński, S.; Czyż, K.; Bodkowski, R.; Herbinger, G.; Patkowska-Sokoła, B.; Librowski, T. Omega-3 Fatty Acids and their Role in Central Nervous System-A Review. Curr. Med. Chem. 2016, 23, 816–831. [Google Scholar] [CrossRef]
- Msanne, J.; Xu, D.; Konda, A.R.; Casas-Mollano, J.A.; Awada, T.; Cahoon, E.B.; Cerutti, H. Metabolic and gene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgae Chlamydomonas reinhardtii and Coccomyxa sp. C-169. Phytochem 2012, 75, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Cha, T.S.; Chen, J.W.; Goh, E.G.; Aziz, A.; Loh, S.H. Differential regulation of fatty acid biosynthesis in two Chlorella species in response to nitrate treatments and the potential of binary blending microalgae oils for biodiesel application. Bioresour. Technol. 2011, 102, 10633–10640. [Google Scholar] [CrossRef]
- Srivastava, G.; Nishchal; Goud, V.V. Salinity induced lipid production in microalgae and cluster analysis (ICCB 16-BR_047). Bioresour. Technol. 2017, 242, 244–252. [Google Scholar] [CrossRef]
- Xia, L.; Song, S.; Hu, C. High temperature enhances lipid accumulation in nitrogen-deprived Scenedesmus obtusus XJ-15. J. Appl. Phycol. 2016, 28, 831–837. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Susttain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotechnol. Adv. 2020, 44, 107612. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Han, B.; Yu, X. Coupling of abiotic stresses and phytohormones for the production of lipids and high-value by-products by microalgae: A review. Bioresour. Technol. 2019, 274, 549–556. [Google Scholar] [CrossRef]
- Contreras-Pool, P.Y.; Peraza-Echeverria, S.; Ku-González, Á.F.; Herrera-Valencia, V.A. The phytohormone abscisic acid increases triacylglycerol content in the green microalga Chlorella saccharophila (Chlorophyta). Algae 2016, 31, 267–276. [Google Scholar] [CrossRef]
- Yu, X.; Chen, L.; Zhang, W. Chemicals to enhance microalgal growth and accumulation of high-value bioproducts. Front. Microbiol. 2015, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Gao, Z.; Du, H.; Lin, B.; Yan, Y.; Li, G.; Guo, Y.; Fu, S.; Wei, G.; Wang, M.; et al. The effects of abscisic acid, salicylic acid and jasmonic acid on lipid accumulation in two freshwater Chlorella strains. J. Gen. Appl. Microbiol. 2018, 64, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Kozlova, T.A.; Hardy, B.P.; Krishna, P.; Levin, D.B. Effect of phytohormones on growth and accumulation of pigments and fatty acids in the microalgae Scenedesmus quadricauda. Algal Res. 2017, 27, 325–334. [Google Scholar] [CrossRef]
- Lin, B.; Ahmed, F.; Du, H.; Li, Z.; Yan, Y.; Huang, Y.; Cui, M.; Yin, Y.; Li, B.; Wang, M.; et al. Plant growth regulators promote lipid and carotenoid accumulation in Chlorella vulgaris. J. Appl Phycol. 2018, 30, 1549–1561. [Google Scholar] [CrossRef]
- Kozlova, T.A.; Hardy, B.P.; Levin, D.B. The combined influence of 24-epibrassinolide and 3-indoleacetic acid on growth and accumulation of pigments and fatty acids in the microalgae Scenedesmus quadricauda (CPCC-158). Algal Res. 2018, 35, 22–32. [Google Scholar] [CrossRef]
- Renuka, N.; Guldhe, A.; Singh, P.; Bux, F. Combined effect of exogenous phytohormones on biomass and lipid production in Acutodesmus obliquus under nitrogen limitation. Energy Convers. Manag. 2018, 168, 522–528. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, D.; Xu, J.; Zhao, P.; Li, T.; Ma, H.; Yu, X. Melatonin enhances lipid production in Monoraphidium sp. QLY-1 under nitrogen deficiency conditions via a multi-level mechanism. Bioresour. Technol. 2018, 259, 46–53. [Google Scholar] [CrossRef]
- Stirk, W.A.; Ördög, V.; Novák, O.; Rolčík, J.; Strnad, M.; Bálint, P.; van Staden, J. Auxin and cytokinin relationships in 24 microalgal strains1. J. Phycol. 2013, 49, 459–467. [Google Scholar] [CrossRef]
- Yoshida, K.; Igarashi, E.; Mukai, M.; Hirata, K.; Miyamoto, K. Induction of tolerance to oxidative stress in the green alga, Chlamydomonas reinhardtii, by abscisic acid. Plant Cell Environ. 2003, 26, 451–457. [Google Scholar] [CrossRef]
- Khan, N.A.; Nazar, R.; Iqbal, N.; Anjum, N.A. Phytohormones and Abiotic Stress Tolerance in Plants; Springer: Berlin, Germany, 2012. [Google Scholar]
- Dao, G.; Wu, G.; Wang, X.; Zhuang, L.; Zhang, T.; Hu, H. Enhanced growth and fatty acid accumulation of microalgae Scenedesmus sp. LX1 by two types of auxin. Bioresour. Technol. 2018, 247, 561–567. [Google Scholar] [CrossRef]
- Chu, J.; Li, Y.; Cui, Y.; Qin, S. The influences of phytohormones on triacylglycerol accumulation in an oleaginous marine diatom Phaeodactylum tricornutum. J. Appl. Phycol. 2019, 31, 1009–1019. [Google Scholar] [CrossRef]
- Norlina, R.; Norashikin, M.N.; Loh, S.H.; Aziz, A.; Cha, T.S. Exogenous Abscisic Acid Supplementation at Early Stationary Growth Phase Triggers Changes in the Regulation of Fatty Acid Biosynthesis in Chlorella vulgaris UMT-M1. Appl. Biochem. Biotechnol. 2020, 191, 1653–1669. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.K.; Chanu, N.K.; Chaurasia, N. Exogenous addition of indole acetic acid and kinetin under nitrogen-limited medium enhances lipid yield and expression of glycerol-3-phosphate acyltransferase & diacylglycerol acyltransferase genes in indigenous microalgae: A potential approach for biodiesel production. Bioresour. Technol. 2020, 297, 122439. [Google Scholar]
- Alsenani, F.; Wass, T.J.; Ma, R.; Eltanahy, E.; Netzel, M.E.; Schenk, P.M. Transcriptome-wide analysis of Chlorella reveals auxin-induced carotenogenesis pathway in green microalgae. Algal Res. 2019, 37, 320–335. [Google Scholar] [CrossRef]
- Park, W.; Yoo, G.; Moon, M.; Kim, C.W.; Choi, Y.; Yang, J. Phytohormone Supplementation Significantly Increases Growth of Chlamydomonas reinhardtii Cultivated for Biodiesel Production. Appl. Biochem. Biotechnol. 2013, 171, 1128–1142. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhou, W.; Wei, L.; Li, J.; Jia, J.; Li, F.; Smith, S.M.; Xu, J. Regulation of the cholesterol biosynthetic pathway and its integration with fatty acid biosynthesis in the oleaginous microalga Nannochloropsis oceanica. Biotechnol. Biofuels 2014, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Sivaramakrishnan, R.; Incharoensakdi, A. Plant hormone induced enrichment of Chlorella sp. omega-3 fatty acids. Biotechnol. Biofuels 2020, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jusoh, M.; Loh, S.H.; Chuah, T.S.; Aziz, A.; Cha, T.S. Indole-3-acetic acid (IAA) induced changes in oil content, fatty acid profiles and expression of four fatty acid biosynthetic genes in Chlorella vulgaris at early stationary growth phase. Phytochem 2015, 111, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Ahmed, F.; Lin, B.; Li, Z.; Huang, Y.; Sun, G.; Ding, H.; Wang, C.; Meng, C.; Gao, Z. The Effects of Plant Growth Regulators on Cell Growth, Protein, Carotenoid, PUFAs and Lipid Production of Chlorella pyrenoidosa ZF Strain. Energies 2017, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Salama, E.; Kabra, A.N.; Ji, M.; Kim, J.R.; Min, B.; Jeon, B. Enhancement of microalgae growth and fatty acid content under the influence of phytohormones. Bioresour. Technol. 2014, 172, 97–103. [Google Scholar] [CrossRef]
- Salama, E.; Jeon, B.; Chang, S.W.; Lee, S.; Roh, H.; Yang, I.; Kurade, M.B.; El-Dalatony, M.M.; Kim, D.; Kim, K.; et al. Interactive effect of indole-3-acetic acid and diethyl aminoethyl hexanoate on the growth and fatty acid content of some microalgae for biodiesel production. J. Clean. Prod. 2017, 168, 1017–1024. [Google Scholar] [CrossRef]
- Wu, T.; Yu, L.; Zhang, Y.; Liu, J. Characterization of fatty acid desaturases reveals stress-induced synthesis of C18 unsaturated fatty acids enriched in triacylglycerol in the oleaginous alga Chromochloris zofingiensis. Biotechnol. Biofuels 2021, 14, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Lei, A.; Chen, H.; Shen, G.; Hu, Z.; Chen, L.; Wang, J. Expression of fatty acid synthesis genes and fatty acid accumulation in haematococcus pluvialis under different stressors. Biotechnol. Biofuels 2012, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szechynska-Hebda, M.; Skrzypek, E.; Browska, G.D.; Cielniak, J.B.; Filek, M.; Dzony, M.W. The role of oxidative stress induced by growth regulators in the regeneration process of wheat. Acta Physiol. Plant. 2007, 29, 327–337. [Google Scholar] [CrossRef]
- Synková, H.; Semorádová, S.; Schnablová, R.; Witters, E.; Hušák, M.; Valcke, R. Cytokinin-induced activity of antioxidant enzymes in transgenic Pssu-ipt tobacco during plant ontogeny. Biol. Plant. 2006, 50, 31–41. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef] [PubMed]
- Cavonius, L.R.; Carlsson, N.; Undeland, I. Quantification of total fatty acids in microalgae: Comparison of extraction and transesterification methods. Anal. Bioanal Chem 2014, 406, 7313–7322. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, gix120. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinform. 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | CK | E1 | E2 | E3 | E4 |
|---|---|---|---|---|---|
| C14:0 (%) | 1.51 ± 0.26 | 1.60 ± 0.05 | 1.34 ± 0.08 | 1.93 ± 0.37 | 1.61 ± 0.31 |
| C15:0 (%) | 1.08 ± 0.21 | 1.13 ± 0.02 | 0.96 ± 0.04 | 1.39 ± 0.26 | 1.15 ± 0.23 |
| C16:0 (%) | 18.74 ± 1.17 | 18.65 ± 0.06 | 18.72 ± 0.42 | 23.85 ± 1.26 *** | 20.21 ± 1.05 |
| C18:0 (%) | 2.43 ± 0.04 | 2.18 ± 0.14 | 1.68 ± 0.08 | 3.12 ± 0.54 | 2.49 ± 0.18 |
| SFA (%) | 23.78 ± 0.66 | 23.56 ± 0.004 | 22.69 ± 0.23 | 30.28 ± 0.09 *** | 25.46 ± 0.33 * |
| C16:1 (%) | 2.92 ± 0.67 | 2.95 ± 0.72 | 3.54 ± 0.14 | 4.14 ± 0.08 | 3.61 ± 0.03 |
| C18:1N9C (%) | 1.86 ± 0.15 | 1.79 ± 0.02 | 1.80 ± 0.41 | 2.63 ± 0.47 | 2.10 ± 0.58 |
| MUFA (%) | 4.78 ± 0.52 | 4.74 ± 0.70 | 5.34 ± 0.55 | 6.77 ± 0.39 ** | 5.71 ± 0.61 |
| C16:2 (%) | 3.32 ± 0.03 | 3.04 ± 0.09 | 2.25 ± 0.84 | 3.62 ± 0.15 | 2.56 ± 0.80 |
| C16:3 (%) | 11.94 ± 0.27 | 10.28 ± 0.28 | 10.59 ± 0.31 | 10.74 ± 0.86 | 10.19 ± 0.36 |
| C16:4 (%) | 11.21 ± 1.04 | 13.61 ± 0.33 ** | 13.95 ± 0.17 ** | 6.73 ± 3.44 *** | 12.38 ± 1.00 |
| C18:2n6c (%) | 11.86 ± 0.92 | 10.56 ± 0.33 | 10.40 ± 0.13 | 12.51 ± 1.19 | 11.10 ± 0.65 |
| C18:3n3 (%) | 33.12 ± 0.82 | 34.22 ± 0.32 | 34.78 ± 0.10 | 29.35 ± 1.54 *** | 32.60 ± 0.19 |
| PUFA (%) | 71.44 ± 1.19 | 71.70 ± 0.70 | 71.97 ± 0.32 | 62.95 ± 0.30 *** | 68.83 ± 0.28 *** |
| UFA (%) | 76.22 ± 0.66 | 76.44 ± 0.004 | 77.31 ± 0.22 | 69.72 ± 0.09 *** | 74.54 ± 0.33 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Dai, Y.; Xu, W.; Wu, X.; Li, Y.; Zhu, H.; Zhou, H. The Growth, Lipid Accumulation and Fatty Acid Profile Analysis by Abscisic Acid and Indol-3-Acetic Acid Induced in Chlorella sp. FACHB-8. Int. J. Mol. Sci. 2022, 23, 4064. https://doi.org/10.3390/ijms23074064
Lin Y, Dai Y, Xu W, Wu X, Li Y, Zhu H, Zhou H. The Growth, Lipid Accumulation and Fatty Acid Profile Analysis by Abscisic Acid and Indol-3-Acetic Acid Induced in Chlorella sp. FACHB-8. International Journal of Molecular Sciences. 2022; 23(7):4064. https://doi.org/10.3390/ijms23074064
Chicago/Turabian StyleLin, Yihua, Yue Dai, Weinan Xu, Xiaobin Wu, Yanyan Li, Hongmei Zhu, and Hantao Zhou. 2022. "The Growth, Lipid Accumulation and Fatty Acid Profile Analysis by Abscisic Acid and Indol-3-Acetic Acid Induced in Chlorella sp. FACHB-8" International Journal of Molecular Sciences 23, no. 7: 4064. https://doi.org/10.3390/ijms23074064