
Direct Reprogramming and Induction of Human Dermal Fibroblasts to Differentiate into iPS-Derived Nucleus Pulposus-like Cells in 3D Culture

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
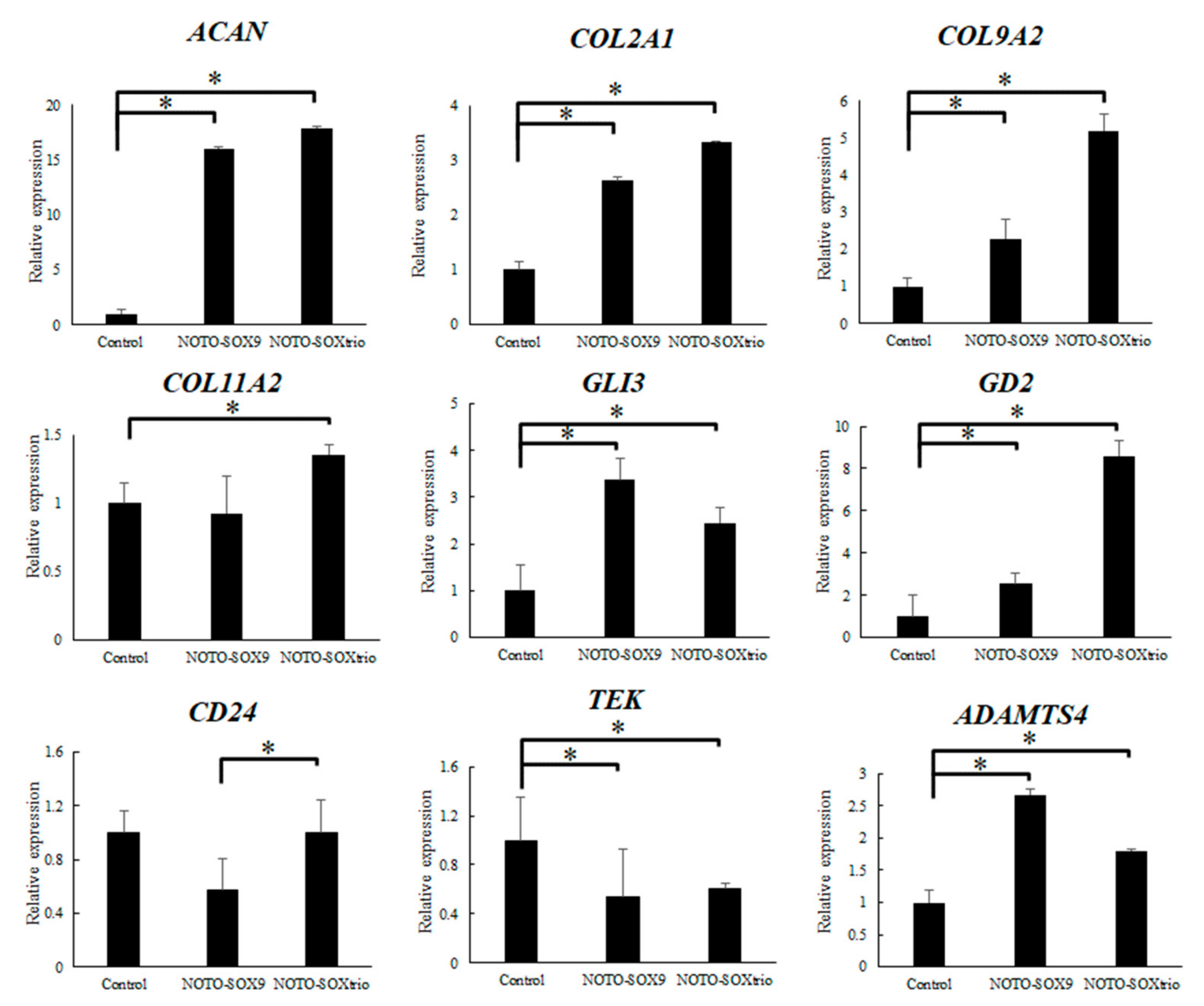
2.1. Transcriptional Analysis of NHDFs
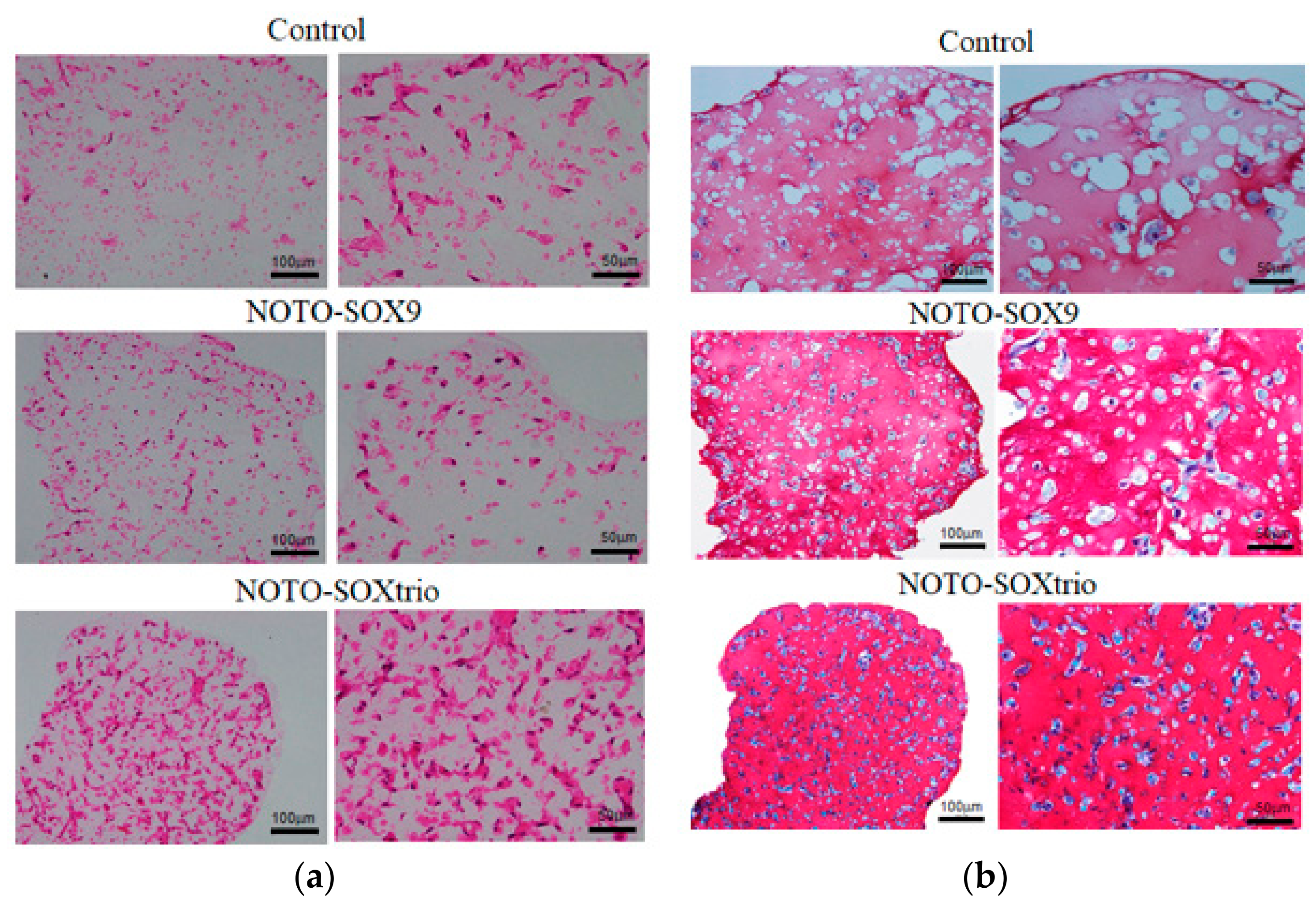
2.2. Analysis of Glycosaminoglyan (CAG) Expression in NHDFs
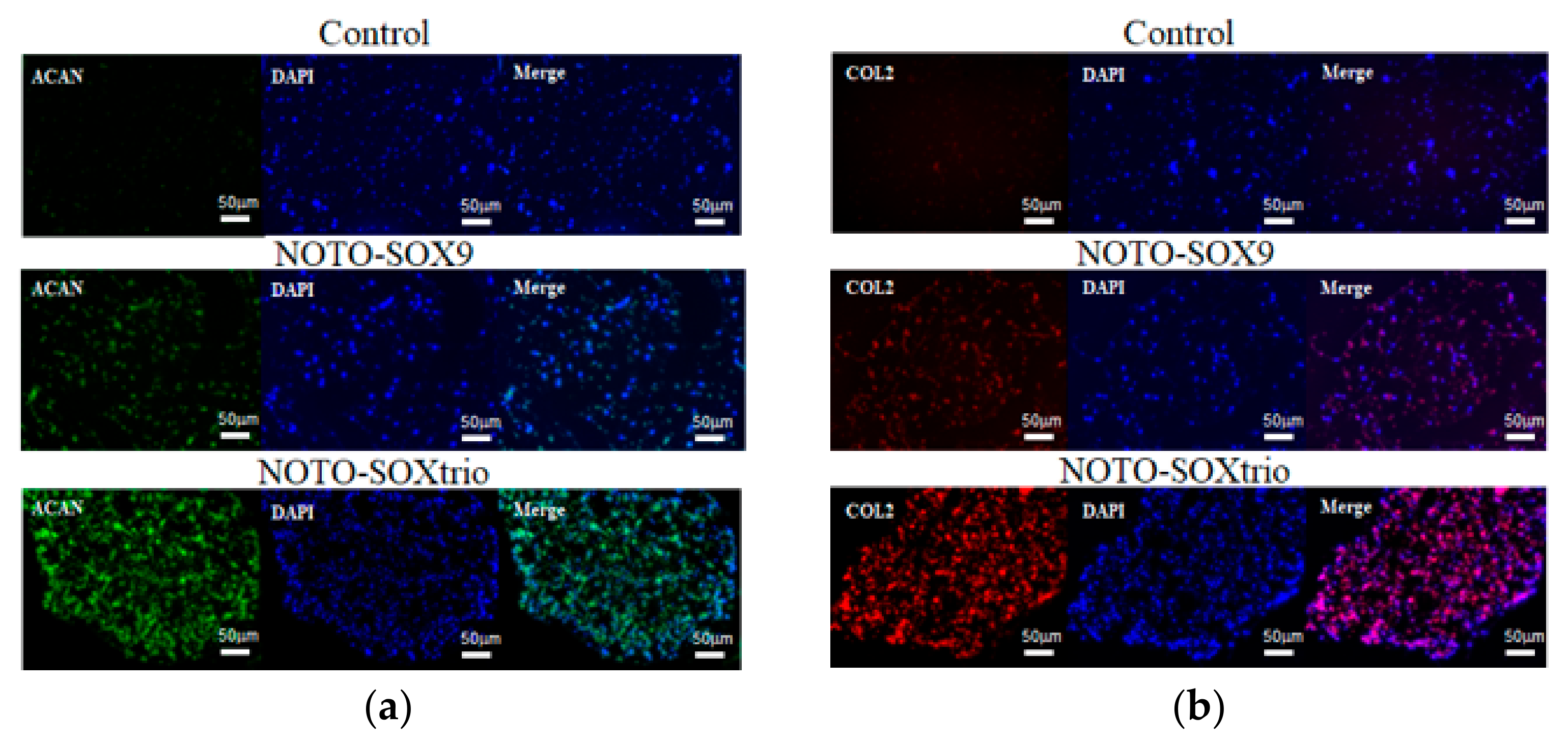
2.3. Immunofluorescence Analysis of the Expression of ECM Components in Cultures of NHDFs
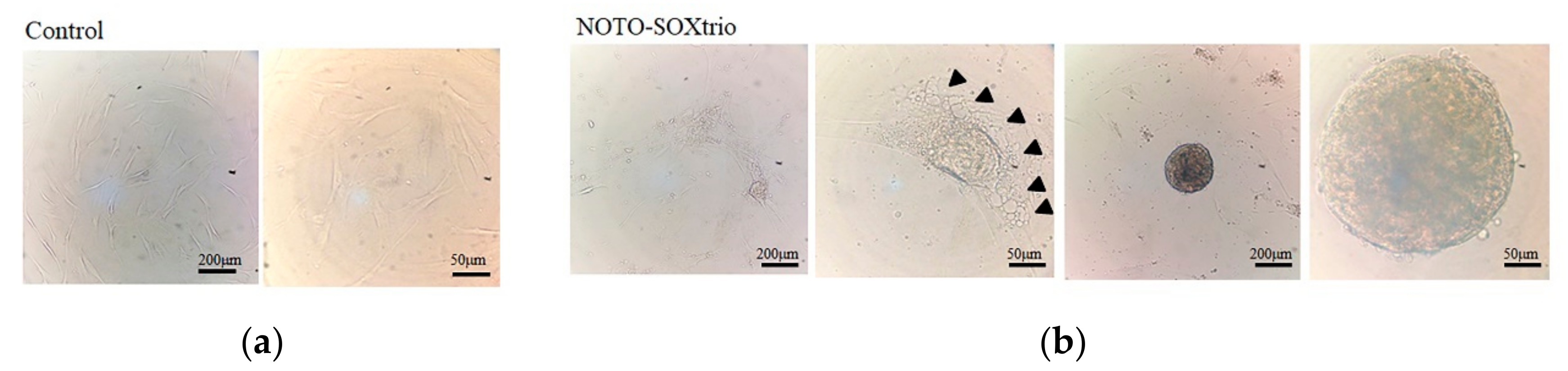
2.4. Detection of Vacuolated Cells in Cultures of the NOTO-SOXtrio Group
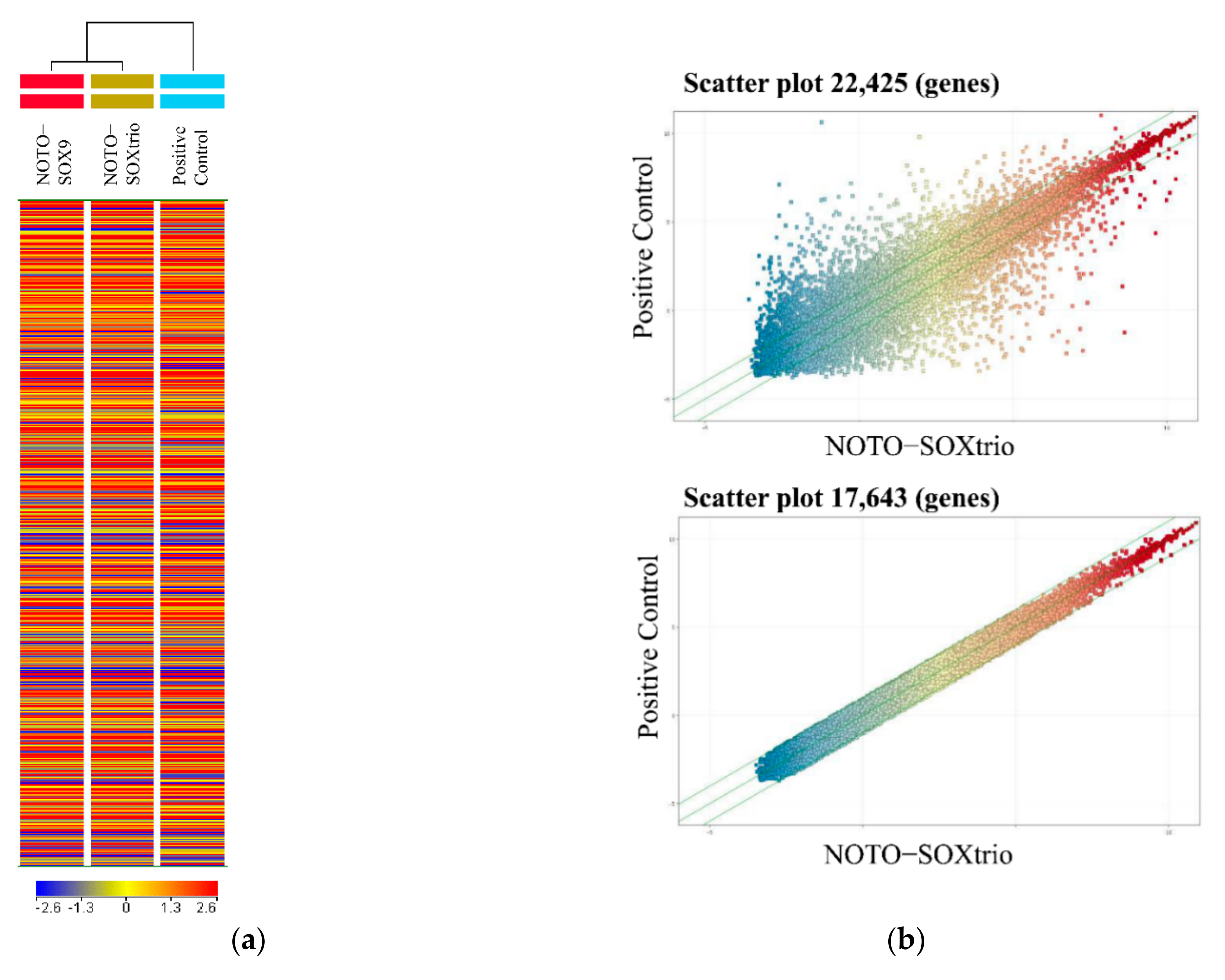
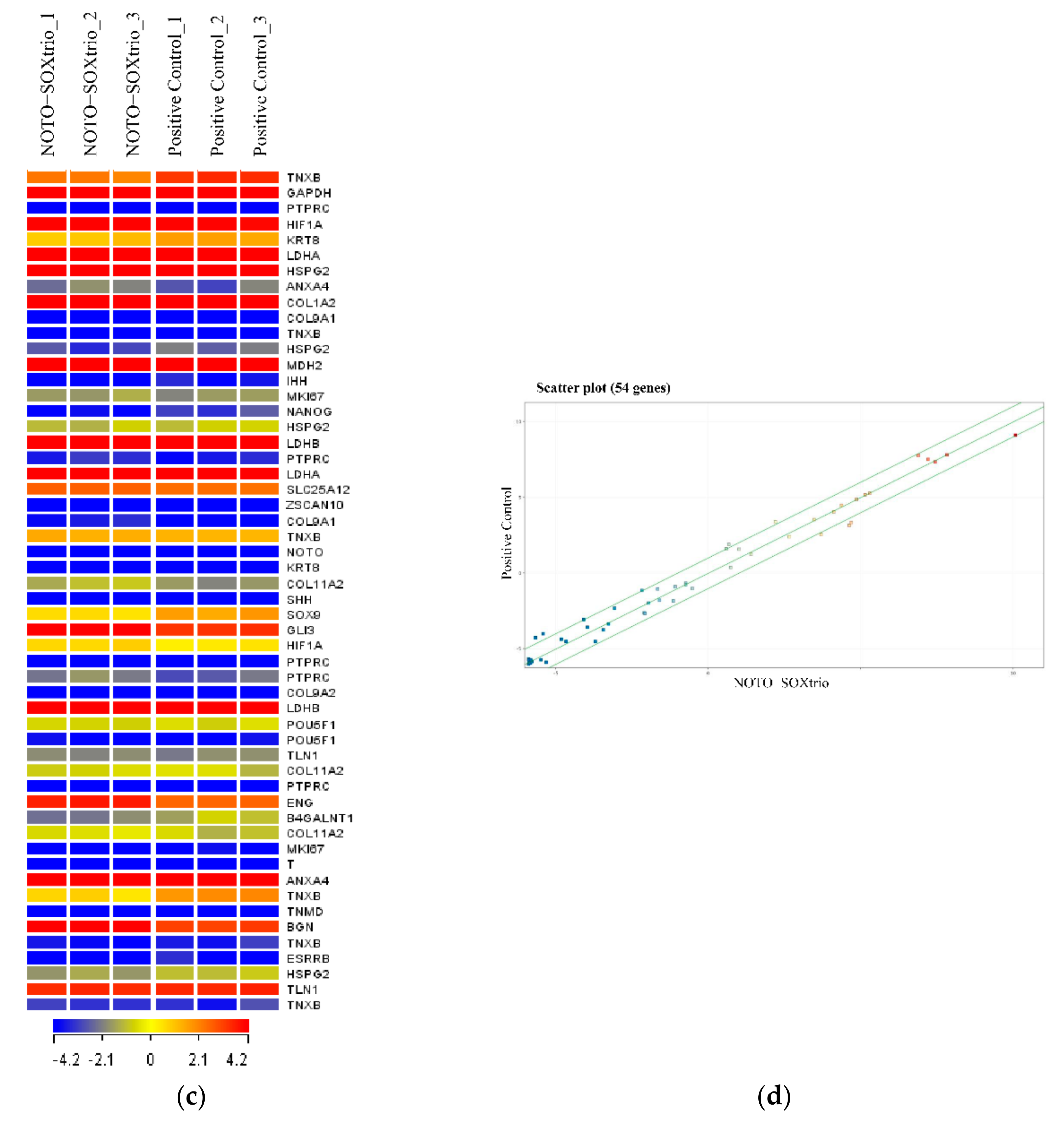
2.5. Genome-Wide Microarray Analysis
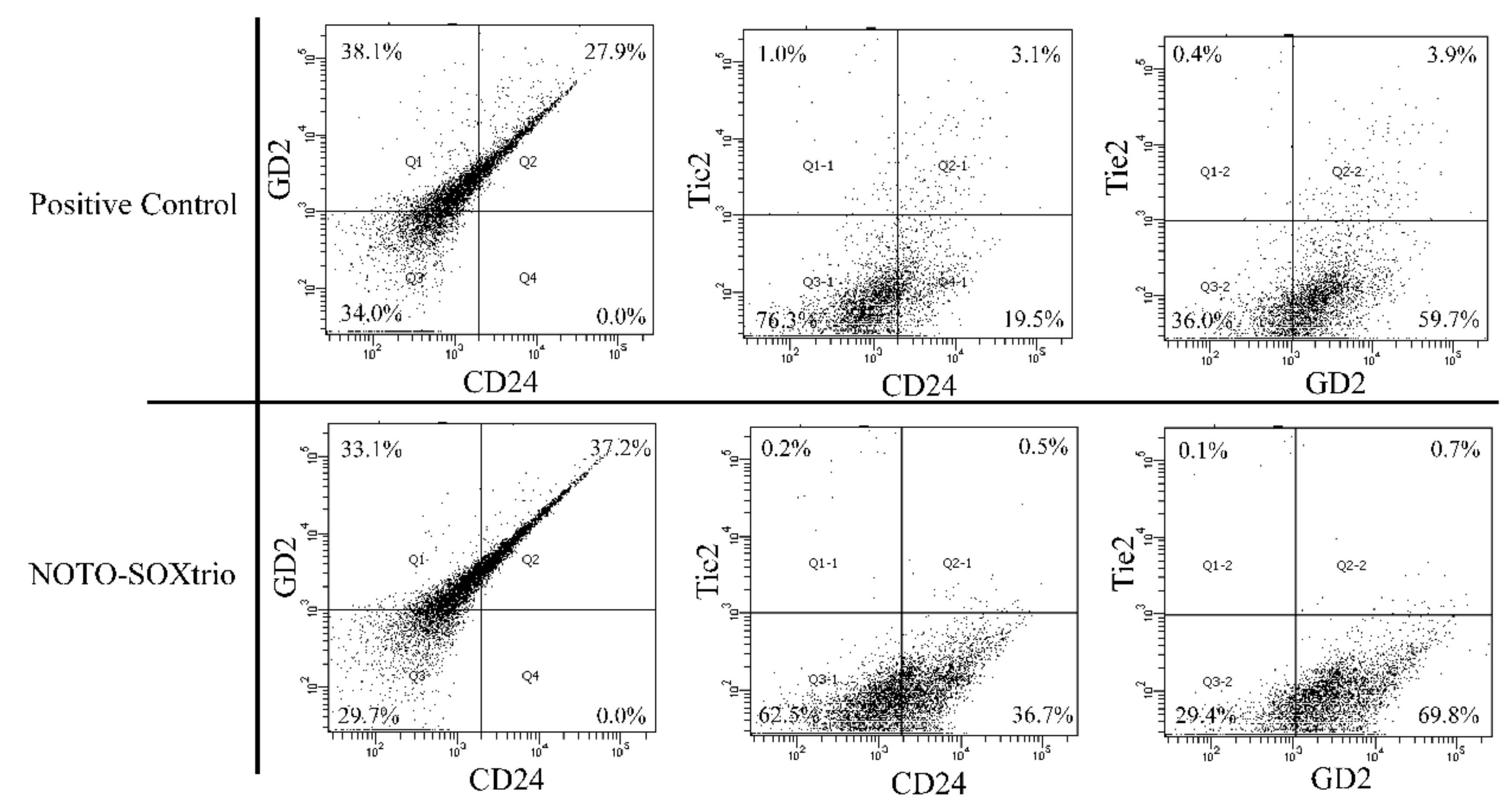
2.6. Fluorescence-Activated Cell Sorting (FACS) Analysis to Determine the State of Differentiation of NP-like Cells
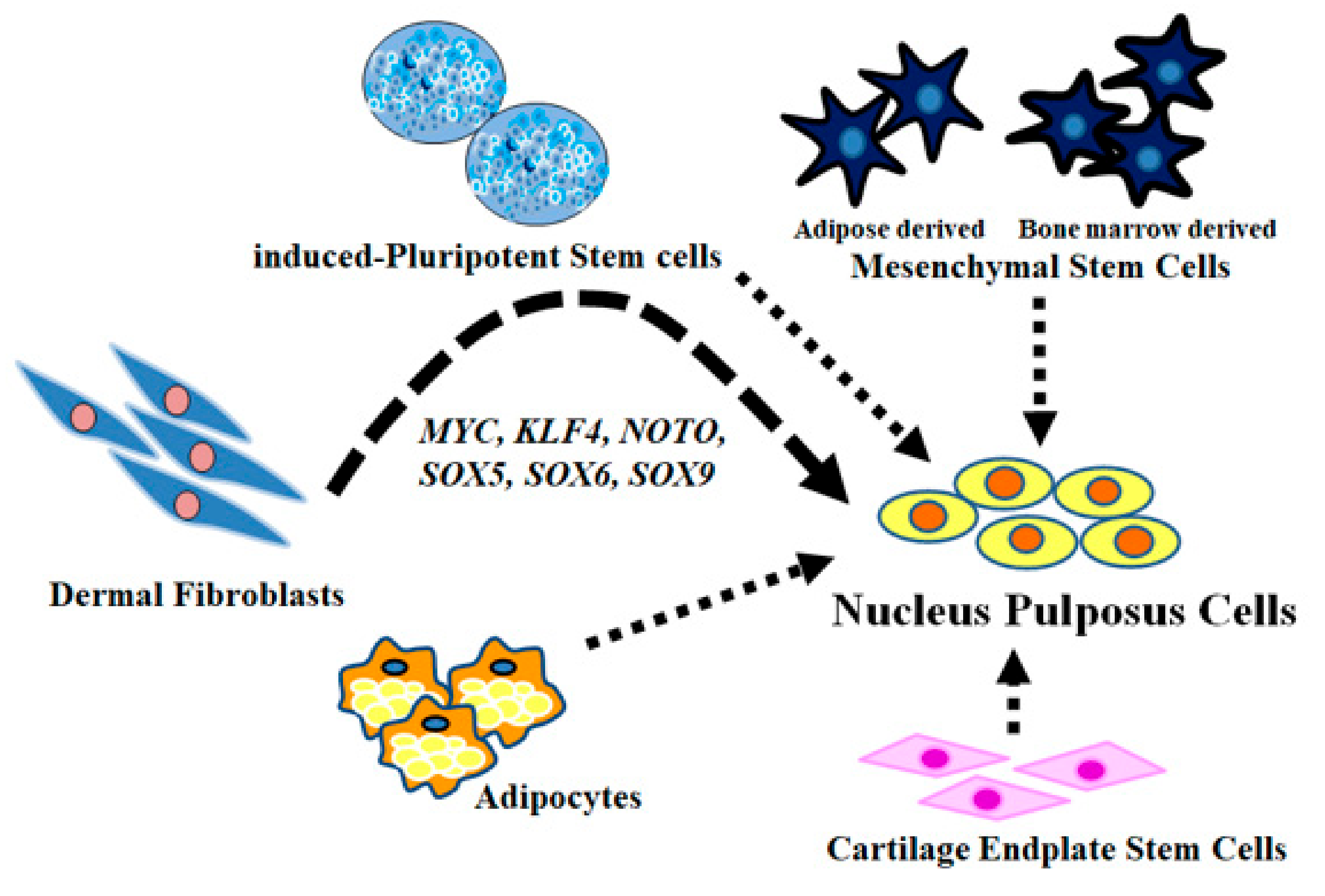
3. Discussion
4. Materials and Methods
4.1. Transfection and Construction of Expression Vectors
4.2. Alginate Beads Culture
4.3. Isolation of Human NP
4.4. qPCR
4.5. Hematoxylin–Eosin and Safranin-O Staining and Immunofluorescence Analysis
4.6. Microarray Analysis
4.7. Flow Cytometry Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAN | Aggrecan |
| ADAMTS4 | ADAM metallopeptidase with thrombospondin type 1 motif 4 |
| AF | Annulus fibrosus |
| BSA | Bovine serum albumin |
| CD24 | CD24 molecule |
| CFU-F | Colony-forming units—fibroblastic |
| CFU-S | Colony-forming units—spherical |
| COL2A1 | Collagen type II alpha 1 chain |
| COL9A2 | Collagen type IX alpha 2 chain |
| COL11A2 | Collagen type XI alpha 2 chain |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s Modified Eagle Medium |
| ECM | Extracellular matrix |
| FBS | Fetal bovine serum |
| FACS | Fluorescence-activated cell sorting |
| FOXA2 | Forkhead box A2 |
| GAG | Glycosaminoglycan |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GD2 | Disialoganglioside GD2 |
| GLI3 | GLI family zinc finger 3 |
| HE | Hematoxylin and eosin |
| IGF | Insulin-like growth factor |
| IVD | Intervertebral disc |
| KLF4 | Kruppel-like factor 4 |
| NHDFs | Normal human dermal fibroblasts |
| NODAL | Nodal growth differentiation factor |
| NOTO | Notochord homeobox |
| NP | Nucleus pulposus |
| MYC | MYC proto-oncogene, bHLH transcription factor |
| PBS | Phosphate-buffered saline |
| qPCR | Quantitative polymerase chain reaction |
| SHH | Sonic hedgehog signaling molecule |
| SOX2 | SRY (sex-determining region Y)-box 2 |
| SOX5 | SRY (sex-determining region Y)-box 5 |
| SOX6 | SRY (sex-determining region Y)-box 6 |
| SOX9 | SRY (sex-determining region Y)-box 9 |
| SOXtrio | SOX5, SOX6, and SOX9 |
| T | Brachyury, T-box transcription factor T |
| TEK | TEK receptor tyrosine kinase |
| TGF-beta 1 | Transforming growth factor-beta 1 |
References
- Lee, C.K.; Langrana, N.A. A review of spinal fusion for degenerative disc disease: Need for alternative treatment approach of disc arthroplasty? Spine J. 2004, 4 (Suppl. S6), 173S–176S. [Google Scholar] [CrossRef] [PubMed]
- Barrick, W.T.; Schofferman, J.A.; Reynolds, J.B.; Goldthwaite, N.D.; McKeehen, M.; Keaney, D.; White, A.H. Anterior lumbar fusion improves discogenic pain at levels of prior posterolateral fusion. Spine 2000, 25, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Largoza, G.; Montenegro, T.S.; Gonzalez, G.A.; Hines, K.; Harrop, J. Lumbar Total Disc Replacement: Current Usage. Neurosurg Clin. N. Am. 2021, 32, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, K.S.; Herkowitz, H.N. Spinal instrumentation in the management of degenerative disorders of the lumbar spine. Clin. Orthop. Relat. Res. 1997, 335, 39–53. [Google Scholar] [CrossRef]
- Kumar, V.; Baburaj, V.; Rajnish, R.K.; Dhatt, S.S. Outcomes of cauda equina syndrome due to lumbar disc herniation after surgical management and the factors affecting it: A systematic review and meta-analysis of 22 studies with 852 cases. Eur. Spine J. 2021, 31, 353–363. [Google Scholar] [CrossRef]
- Schmitz, A.; Collinet, A.; Ntilikina, Y.; Tigan, L.; Charles, Y.P.; Steib, J.P. Revision Surgery of Total Lumbar Disk Replacement: Review of 48 Cases. Clin. Spine Surg. 2021, 34, E315–E322. [Google Scholar] [CrossRef]
- Sakai, D.; Mochida, J.; Iwashina, T.; Watanabe, T.; Nakai, T.; Ando, K.; Hotta, T. Differentiation of mesenchymal stem cells transplanted to a rabbit degenerative disc model: Potential and limitations for stem cell therapy in disc regeneration. Spine 2005, 30, 2379–2387. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.T.; Milby, A.H.; Chiaro, J.A.; Kim, D.H.; Hebela, N.M.; Smith, L.J.; Elliott, D.M.; Mauck, R.L. Translation of an engineered nanofibrous disc-like angle-ply structure for intervertebral disc replacement in a small animal model. Acta Biomater. 2014, 10, 2473–2481. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Fujita, N.; Hosogane, N.; Watanabe, K.; Ishii, K.; Toyama, Y.; Takubo, K.; Horiuchi, K.; Miyamoto, T.; Nakamura, M.; et al. Excessive reactive oxygen species are therapeutic targets for intervertebral disc degeneration. Arthritis Res. Ther. 2015, 17, 316. [Google Scholar] [CrossRef] [Green Version]
- Makino, H.; Seki, S.; Yahara, Y.; Shiozawa, S.; Aikawa, Y.; Motomura, H.; Nogami, M.; Watanabe, K.; Sainoh, T.; Ito, H.; et al. A selective inhibition of c-Fos/activator protein-1 as a potential therapeutic target for intervertebral disc degeneration and associated pain. Sci. Rep. 2017, 7, 16983. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outani, H.; Okada, M.; Yamashita, A.; Nakagawa, K.; Yoshikawa, H.; Tsumaki, N. Direct induction of chondrogenic cells from human dermal fibroblast culture by defined factors. PLoS ONE 2013, 8, e77365. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, K.; Sasagawa, S.; Outani, H.; Nakagawa, K.; Yoshikawa, H.; Tsumaki, N. Generation of hyaline cartilaginous tissue from mouse adult dermal fibroblast culture by defined factors. J. Clin. Investig. 2011, 21, 640–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, V.; Huang, W.; Harley, V.R.; Goodfellow, P.N.; de Crombrugghe, B. SOX9 is a potent activator of the chondrocyte-specific enhancer of the pro alpha1(II) collagen gene. Mol. Cell. Biol. 1997, 17, 2336–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef]
- Barrionuevo, F.; Taketo, M.M.; Scherer, G.; Kispert, A. Sox9 is required for notochord maintenance in mice. Dev. Biol. 2006, 295, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, V.; Li, P.; de Crombrugghe, B. A new long form of Sox5 (L-Sox5), Sox6 and Sox9 are coexpressed in chondrogenesis and cooperatively activate the type II collagen gene. EMBO J. 1998, 17, 5718–5733. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Kamekura, S.; Mabuchi, A.; Kou, I.; Seki, S.; Takato, T.; Nakamura, K.; Kawaguchi, H.; Ikegawa, S.; Chung, U.I. The combination of SOX5, SOX6, and SOX9 (the SOX trio) provides signals sufficient for induction of permanent cartilage. Arthritis Rheum. 2004, 50, 3561–3573. [Google Scholar] [CrossRef]
- Han, Y.; Lefebvre, V. L-Sox5 and Sox6 drive expression of the aggrecan gene in cartilage by securing binding of Sox9 to a far-upstream enhancer. Mol. Cell. Biol. 2008, 28, 4999–5013. [Google Scholar] [CrossRef] [Green Version]
- Smits, P.; Lefebvre, V. Sox5 and Sox6 are required for notochord extracellular matrix sheath formation, notochord cell survival and development of the nucleus pulposus of intervertebral discs. Development 2003, 130, 1135–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombier, P.; Halgand, B.; Chédeville, C.; Chariau, C.; François-Campion, V.; Kilens, S.; Vedrenne, N.; Clouet, J.; David, L.; Guicheux, J.; et al. NOTO Transcription Factor Directs Human Induced Pluripotent Stem Cell-Derived Mesendoderm Progenitors to a Notochordal Fate. Cells 2020, 9, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, M.R.; Tamplin, O.J.; Rossant, J.; Séguin, C.A. Tracing notochord-derived cells using a Noto-cre mouse: Implications for intervertebral disc development. Dis. Model. Mech. 2012, 5, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Z.; Chen, P.; Ma, C.Y.; Li, C.; Au, T.Y.K.; Tam, V.; Peng, Y.; Wu, R.; Cheung, K.M.C.; et al. Directed Differentiation of Notochord-like and Nucleus Pulposus-like Cells Using Human Pluripotent Stem Cells. Cell Rep. 2020, 30, 2791–2806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Kapper, D.; Youngs, B.; Kocsis, V.; Mondal, S.; Kraus, P.; Lufkin, T. Potential biomarkers of the mature intervertebral disc identified at the single cell level. J. Anat. 2019, 234, 16–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues-Pinto, R.; Richardson, S.M.; Hoyland, J.A. An understanding of intervertebral disc development, maturation and cell phenotype provides clues to direct cell-based tissue regeneration therapies for disc degeneration. Eur. Spine J. 2014, 23, 1803–1814. [Google Scholar] [CrossRef]
- Nakamichi, R.; Asahara, H. The transcription factors regulating intervertebral disc development. JOR Spine 2020, 3, e1081. [Google Scholar] [CrossRef] [Green Version]
- Wagner, T.; Wirth, J.; Meyer, J.; Zabel, B.; Held, M.; Zimmer, J.; Pasantes, J.; Bricarelli, F.D.; Keutel, J.; Hustert, E.; et al. Autosomal sex reversal and campomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell 1994, 79, 1111–1120. [Google Scholar] [CrossRef]
- Tsingas, M.; Ottone, O.K.; Haseeb, A.; Barve, R.A.; Shapiro, I.M.; Lefebvre, V.; Risbud, M.V. Sox9 deletion causes severe intervertebral disc degeneration characterized by apoptosis, matrix remodeling, and compartment-specific transcriptomic changes. Matrix Biol. 2020, 94, 110–133. [Google Scholar] [CrossRef]
- Buttitta, L.; Mo, R.; Hui, C.C.; Fan, C.M. Interplays of Gli2 and Gli3 and their requirement in mediating Shh-dependent sclerotome induction. Development 2003, 130, 6233–6243. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Kapper, D.; Mondal, S.; Lufkin, T.; Kraus, P. Quantitative Single-Cell Transcript Assessment of Biomarkers Supports Cellular Heterogeneity in the Bovine IVD. Vet. Sci. 2019, 6, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashbass, P.; Cooke, L.A.; Herrmann, B.G.; Beddington, R.S. A cell autonomous function of Brachyury in T/T embryonic stem cell chimaeras. Nature 1991, 353, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Kwan, K.M.; Mackem, S. Putative oncogene Brachyury (T) is essential to specify cell fate but dispensable for notochord progenitor proliferation and EMT. Proc. Natl. Acad. Sci. USA 2016, 113, 3820–3825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ang, S.L.; Rossant, J. HNF-3 beta is essential for node and notochord formation in mouse development. Cell 1994, 78, 561–574. [Google Scholar] [CrossRef]
- Fujita, N.; Miyamoto, T.; Imai, J.; Hosogane, N.; Suzuki, T.; Yagi, M.; Morita, K.; Ninomiya, K.; Miyamoto, K.; Takaishi, H.; et al. CD24 is expressed specifically in the nucleus pulposus of intervertebral discs. Biochem. Biophys. Res. Commun. 2005, 338, 1890–1896. [Google Scholar] [CrossRef] [PubMed]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.; et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, J.; Häckel, S.; Croft, A.S.; Albers, C.E.; Gantenbein, B. The effects of 3D culture on the expansion and maintenance of nucleus pulposus progenitor cell multipotency. JOR Spine 2020, 4, e1131. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Jacobsen, T.D.; Barati, Z.; Chahine, N.O. Confocal scanning of intervertebral disc cells in 3D: Inside alginate beads and in native microenvironment. JOR Spine 2020, 3, e1106. [Google Scholar] [CrossRef]
- Baffi, M.O.; Moran, M.A.; Serra, R. Tgfbr2 regulates the maintenance of boundaries in the axial skeleton. Dev. Biol. 2006, 296, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Shen, J.; Wang, B.; Wang, M.; Shu, B.; Chen, D. TGF-β signaling plays an essential role in the growth and maintenance of intervertebral disc tissue. FEBS Lett. 2011, 585, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Matta, A.; Karim, M.Z.; Isenman, D.E.; Erwin, W.M. Molecular Therapy for Degenerative Disc Disease: Clues from Secretome Analysis of the Notochordal Cell-Rich Nucleus Pulposus. Sci. Rep. 2017, 7, 45623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Jiang, C.; Yu, C.; Shen, H. Differentiation of transforming growth factor β1-induced mesenchymal stem cells into nucleus pulposus-like cells under simulated microgravity conditions. Cell. Mol. Biol. 2015, 61, 50–55. [Google Scholar] [PubMed]
- Clarke, L.E.; McConnell, J.C.; Sherratt, M.J.; Derby, B.; Richardson, S.M.; Hoyland, J.A. Growth differentiation factor 6 and transforming growth factor-beta differentially mediate mesenchymal stem cell differentiation, composition, and micromechanical properties of nucleus pulposus constructs. Arthritis Res. Ther. 2014, 16, R67. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Gong, J.; Zhang, H.; Qin, J.; Li, C.; Zhang, J.; Tang, Y.; Zhang, Y.; Chen, J.; Zhou, Y.; et al. Cartilage Endplate Stem Cells Transdifferentiate into Nucleus Pulposus Cells via Autocrine Exosomes. Front. Cell Dev. Biol. 2021, 9, 648201. [Google Scholar] [CrossRef]
- Colombier, P.; Clouet, J.; Boyer, C.; Ruel, M.; Bonin, G.; Lesoeur, J.; Moreau, A.; Fellah, B.H.; Weiss, P.; Lescaudron, L.; et al. TGF-β1 and GDF5 Act Synergistically to Drive the Differentiation of Human Adipose Stromal Cells toward Nucleus Pulposus-like Cells. Stem Cells. 2016, 34, 653–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Guan, Y.; Zhang, Z.; Xiong, Y.; Liu, C.; Li, H.; Liu, B. Comparison of the differentiation abilities of bone marrow-derived mesenchymal stem cells and adipose-derived mesenchymal stem cells toward nucleus pulposus-like cells in three-dimensional culture. Exp. Ther. Med. 2021, 22, 1018. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Ueda, Y.; Miyazaki, K.; Koizumi, M.; Takakura, Y. Disc regeneration therapy using marrow mesenchymal cell transplantation: A report of two case studies. Spine 2010, 35, E475–80. [Google Scholar] [CrossRef]
- Li, Y.Y.; Diao, H.J.; Chik, T.K.; Chow, C.T.; An, X.M.; Leung, V.; Cheung, K.M.; Chan, B.P. Delivering mesenchymal stem cells in collagen microsphere carriers to rabbit degenerative disc: Reduced risk of osteophyte formation. Tissue Eng. Part A 2014, 20, 1379–1391. [Google Scholar] [CrossRef] [Green Version]
- Coric, D.; Pettine, K.; Sumich, A.; Boltes, M.O. Prospective study of disc repair with allogeneic chondrocytes presented at the 2012 Joint Spine Section Meeting. J. Neurosurg. Spine 2013, 18, 85–95. [Google Scholar] [CrossRef]
- Kim, D.H.; Martin, J.T.; Elliott, D.M.; Smith, L.J.; Mauck, R.L. Phenotypic stability, matrix elaboration and functional maturation of nucleus pulposus cells encapsulated in photocrosslinkable hyaluronic acid hydrogels. Acta Biomater. 2015, 12, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhou, Y.; Huang, B.; Liu, L.T.; Liu, M.H.; Wang, J.; Li, C.Q.; Zhang, Z.F.; Chu, T.W.; Xiong, C.J. Utilization of stem cells in alginate for nucleus pulposus tissue engineering. Tissue Eng. Part A 2014, 20, 908–920. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seki, S.; Iwasaki, M.; Makino, H.; Yahara, Y.; Miyazaki, Y.; Kamei, K.; Futakawa, H.; Nogami, M.; Tran Canh Tung, N.; Hirokawa, T.; et al. Direct Reprogramming and Induction of Human Dermal Fibroblasts to Differentiate into iPS-Derived Nucleus Pulposus-like Cells in 3D Culture. Int. J. Mol. Sci. 2022, 23, 4059. https://doi.org/10.3390/ijms23074059
Seki S, Iwasaki M, Makino H, Yahara Y, Miyazaki Y, Kamei K, Futakawa H, Nogami M, Tran Canh Tung N, Hirokawa T, et al. Direct Reprogramming and Induction of Human Dermal Fibroblasts to Differentiate into iPS-Derived Nucleus Pulposus-like Cells in 3D Culture. International Journal of Molecular Sciences. 2022; 23(7):4059. https://doi.org/10.3390/ijms23074059
Chicago/Turabian StyleSeki, Shoji, Mami Iwasaki, Hiroto Makino, Yasuhito Yahara, Yoshitaka Miyazaki, Katsuhiko Kamei, Hayato Futakawa, Makiko Nogami, Nguyen Tran Canh Tung, Tatsuro Hirokawa, and et al. 2022. "Direct Reprogramming and Induction of Human Dermal Fibroblasts to Differentiate into iPS-Derived Nucleus Pulposus-like Cells in 3D Culture" International Journal of Molecular Sciences 23, no. 7: 4059. https://doi.org/10.3390/ijms23074059