Cloning and Organelle Expression of Bamboo Mitochondrial Complex I Subunits Nad1, Nad2, Nad4, and Nad5 in the Yeast Saccharomyces cerevisiae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. cDNA Alignments of nad1, nad2, nad4, and nad5 from B. oldhamii and P. edulis with Other Monocots
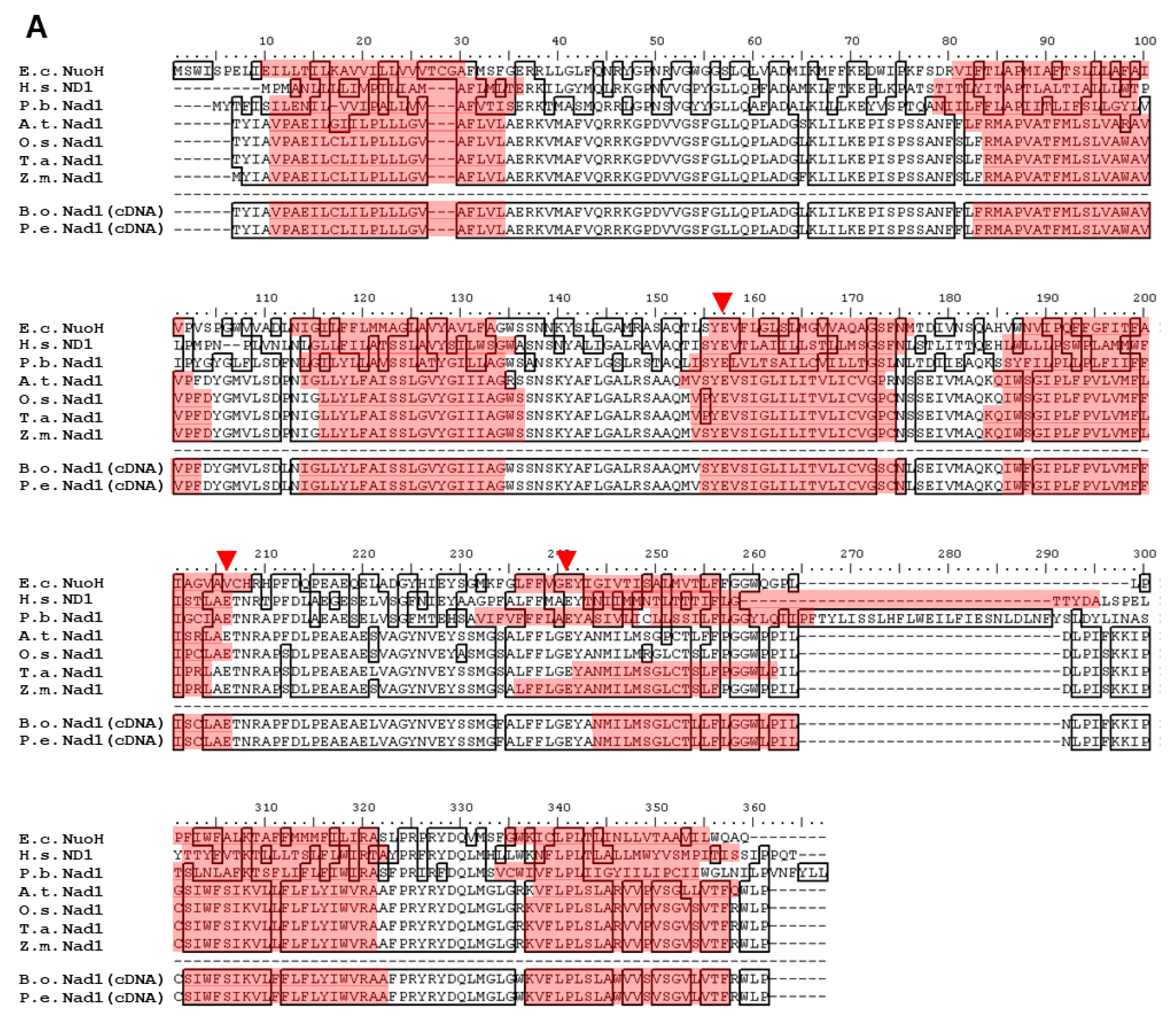
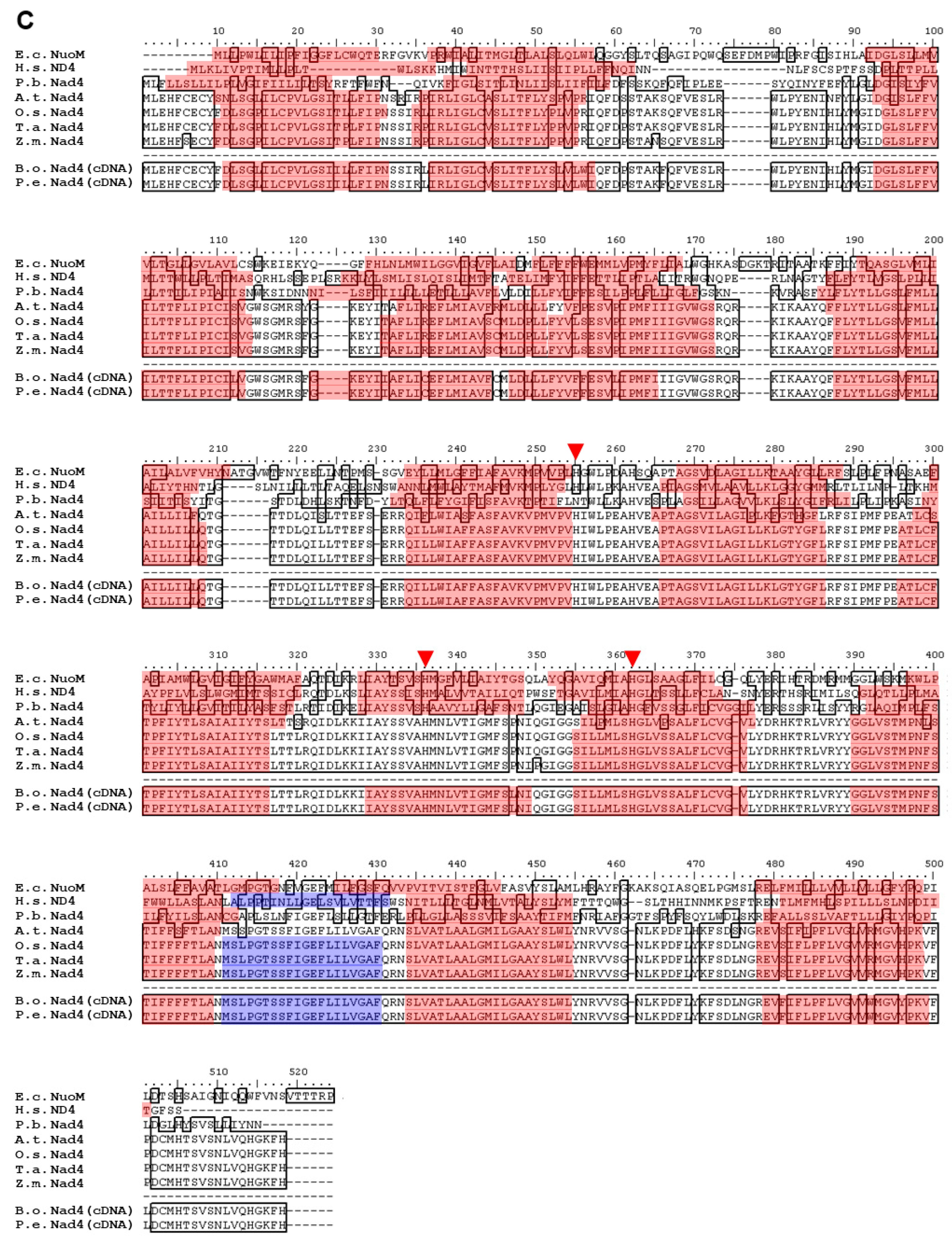
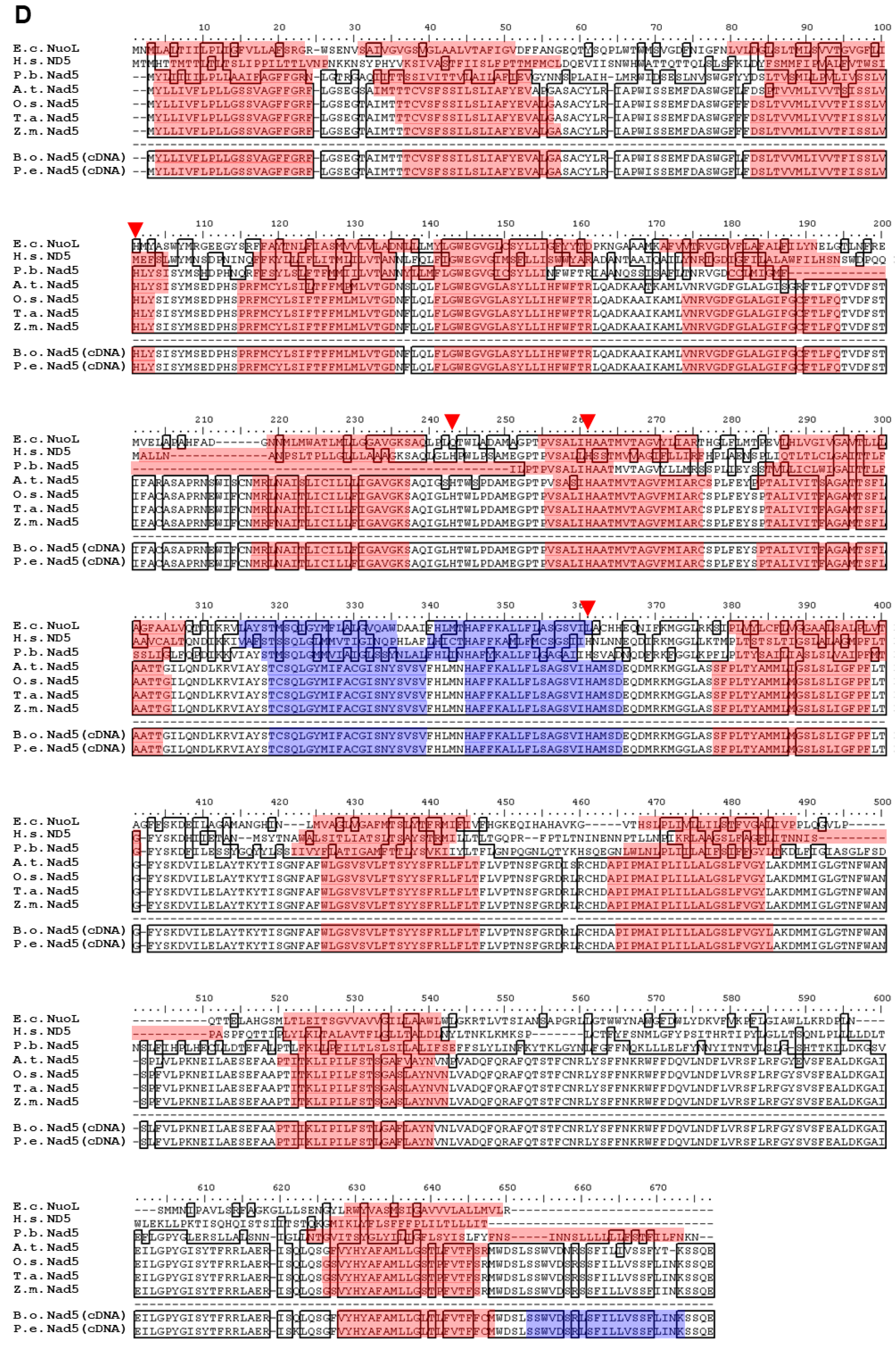
2.2. Polypeptide Alignment of Nad1, Nad2, Nad4, and Nad5 from B. oldhamii and P. edulis with Other Species
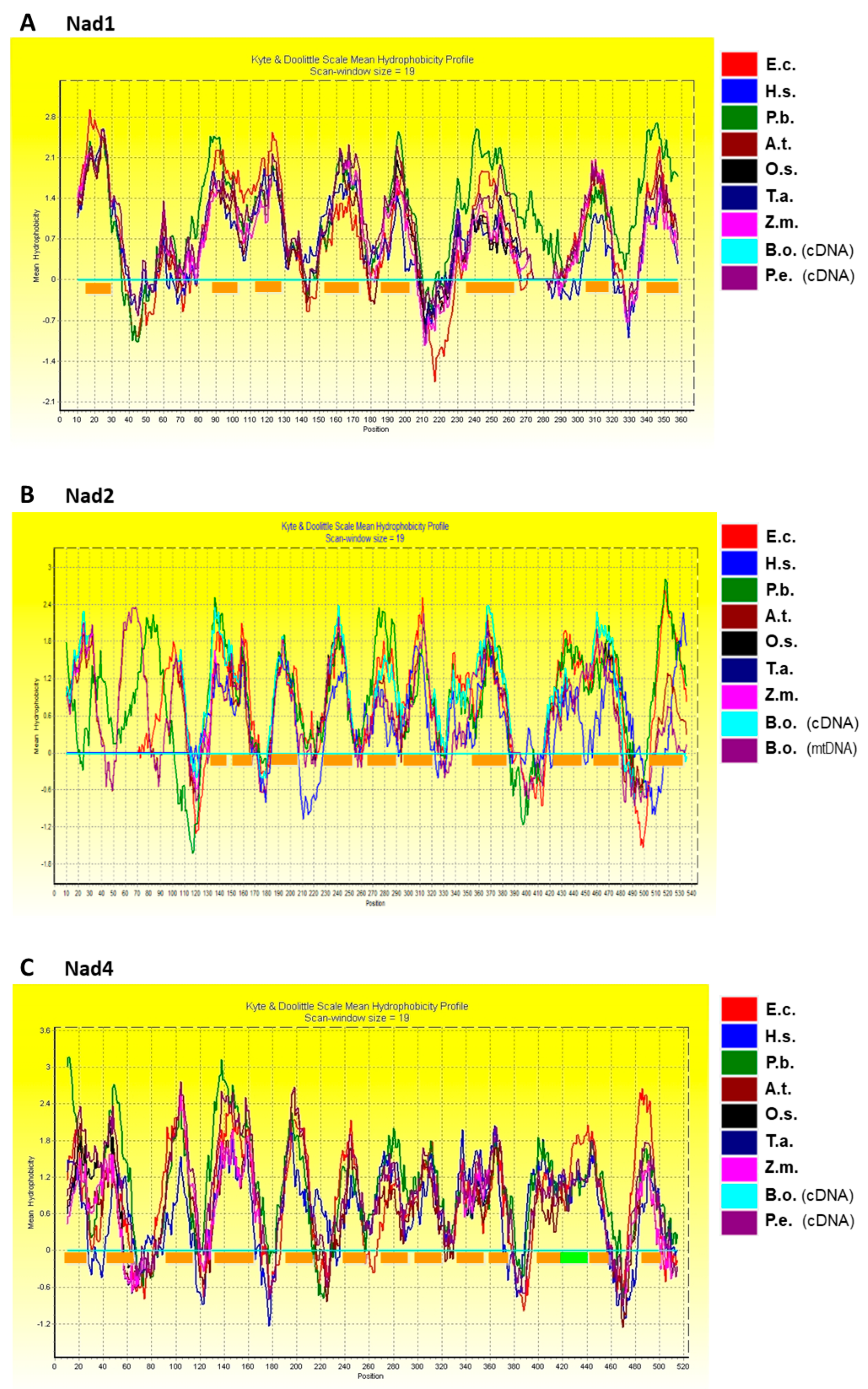
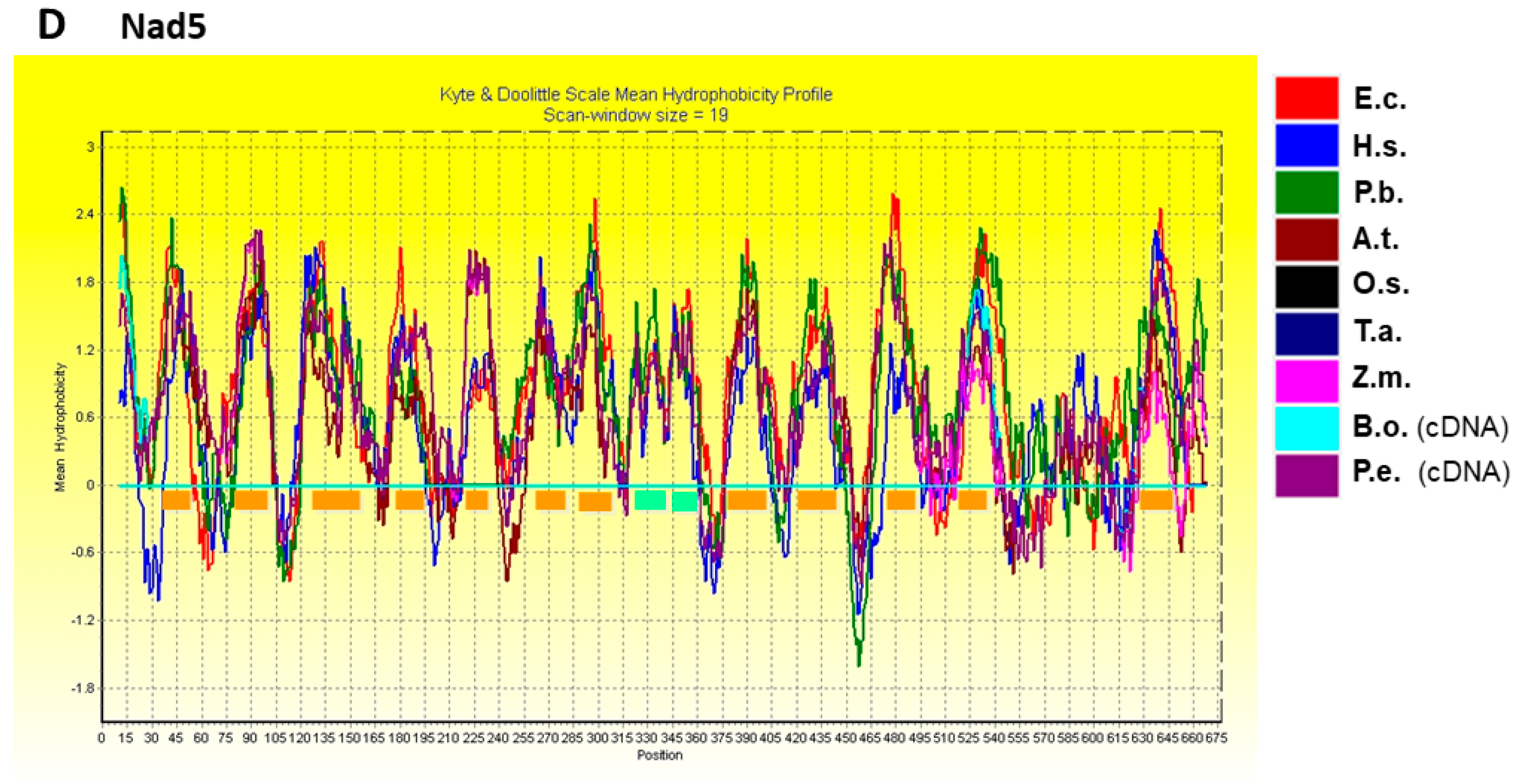
2.3. Hydropathy Plots of Nad1, Nad2, Nad4, and Nad5
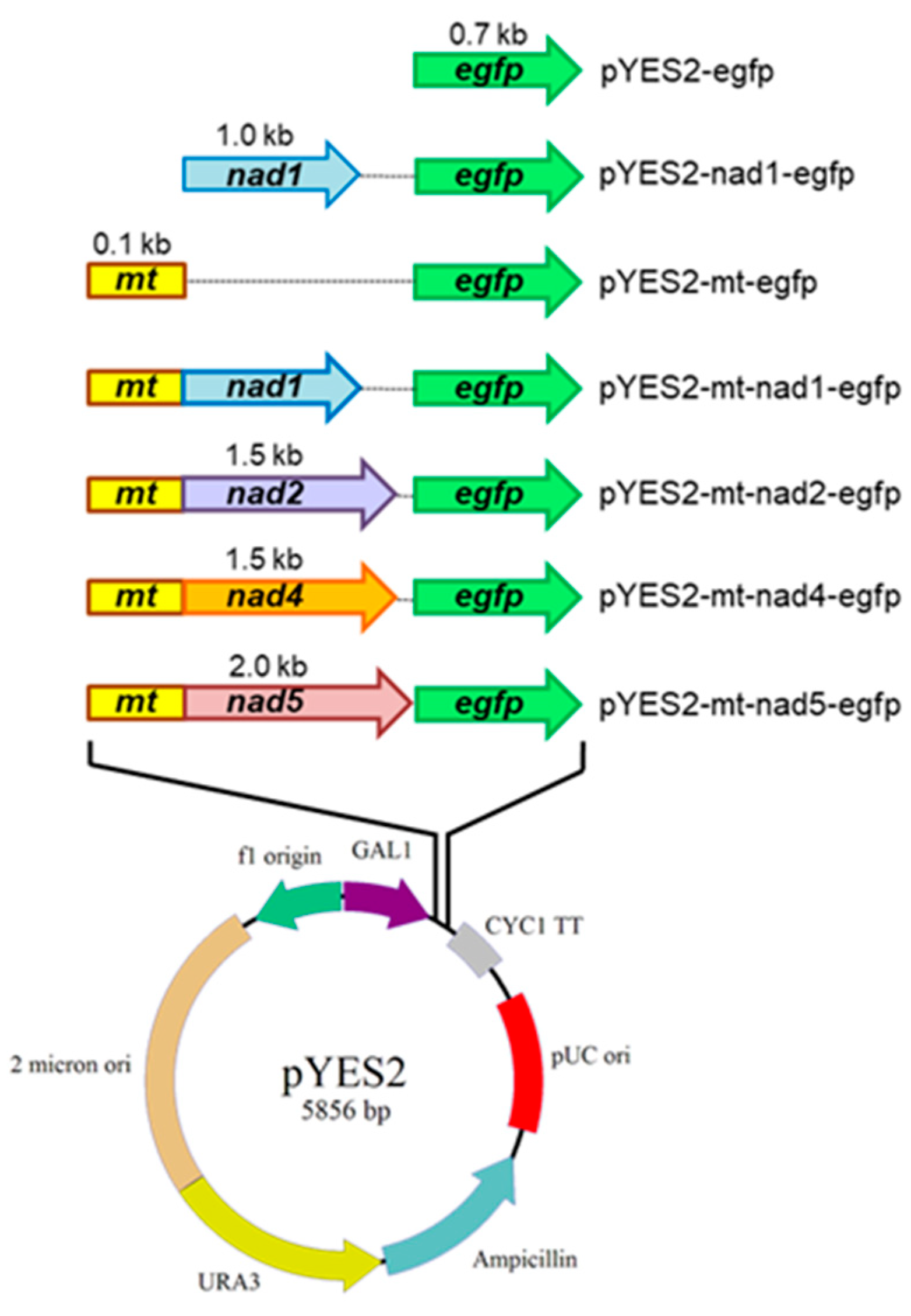
2.4. Construction of pYES2-mt-egfp Plasmid-Carrying nad1, nad2, nad4, or nad5 Genes
2.5. Yeast Transformants
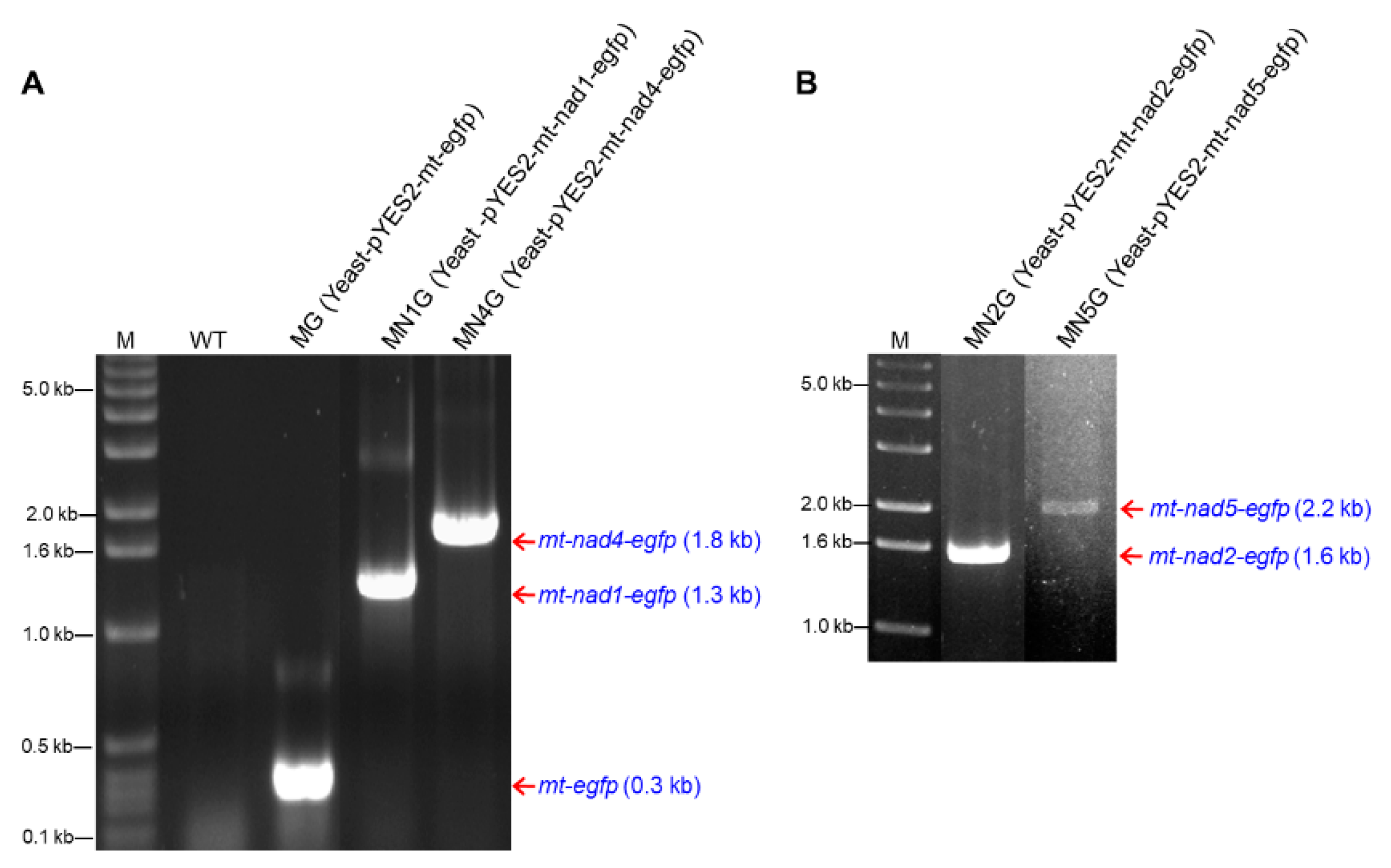
2.5.1. Yeast Transformants Containing pYES2-mt-nad1-egfp, pYES2-mt-nad2-egfp, pYES2-mt-nad4-egfp, or pYES2-mt-nad5-egfp
2.5.2. RNA Transcription of Fusion Genes in Yeast Transformants
2.5.3. Expression of MT-Nad1-EGFP, MT-Nad2-EGFP, MT-Nad4-EGFP, or MT-Nad5-EGFP Fusion Proteins in Yeast Transformants
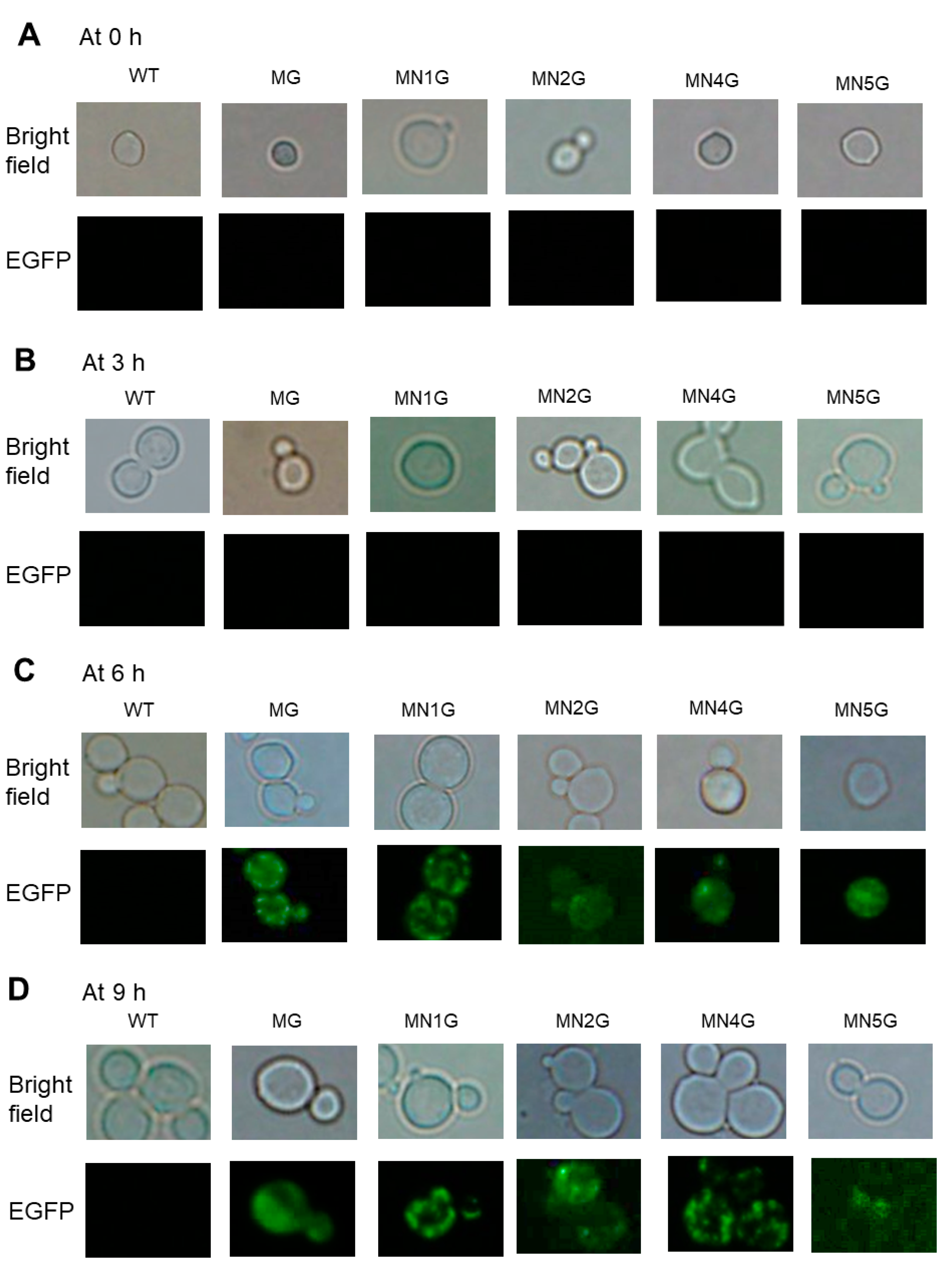
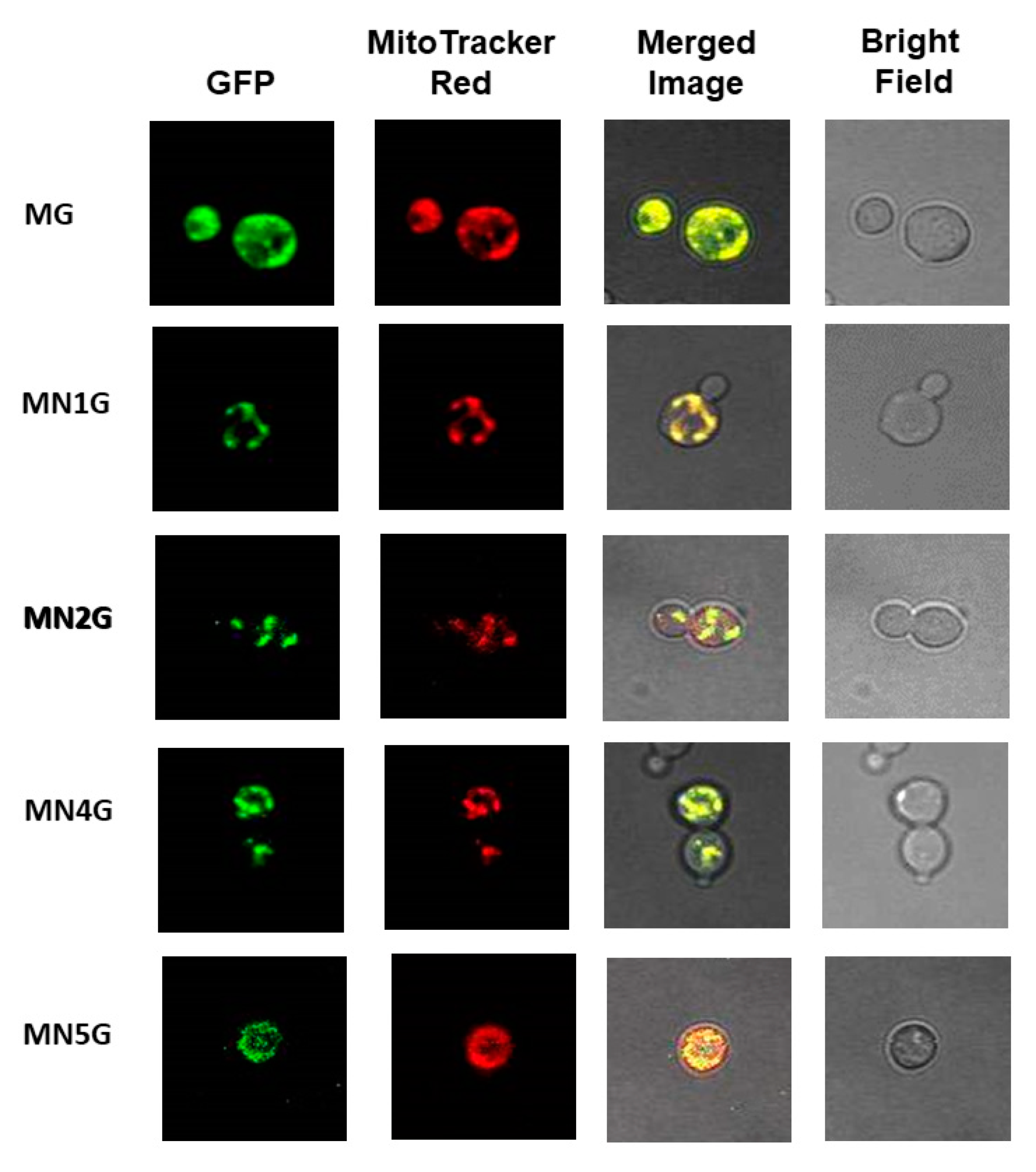
2.5.4. Subcellular Localization of MT-NAD1-EGFP, MT-NAD2-EGFP, MT-NAD4-EGFP, or MT-NAD5-EGFP Fusion Proteins in Yeast Observed Using EGFP Fluorescence
2.6. K+, Na+, or H+-Dependent Growth Phenotypes of Yeast Transformants
3. Discussion
3.1. Highly Conserved Nad1, Nad2, Nad4, and Nad5 in Plants
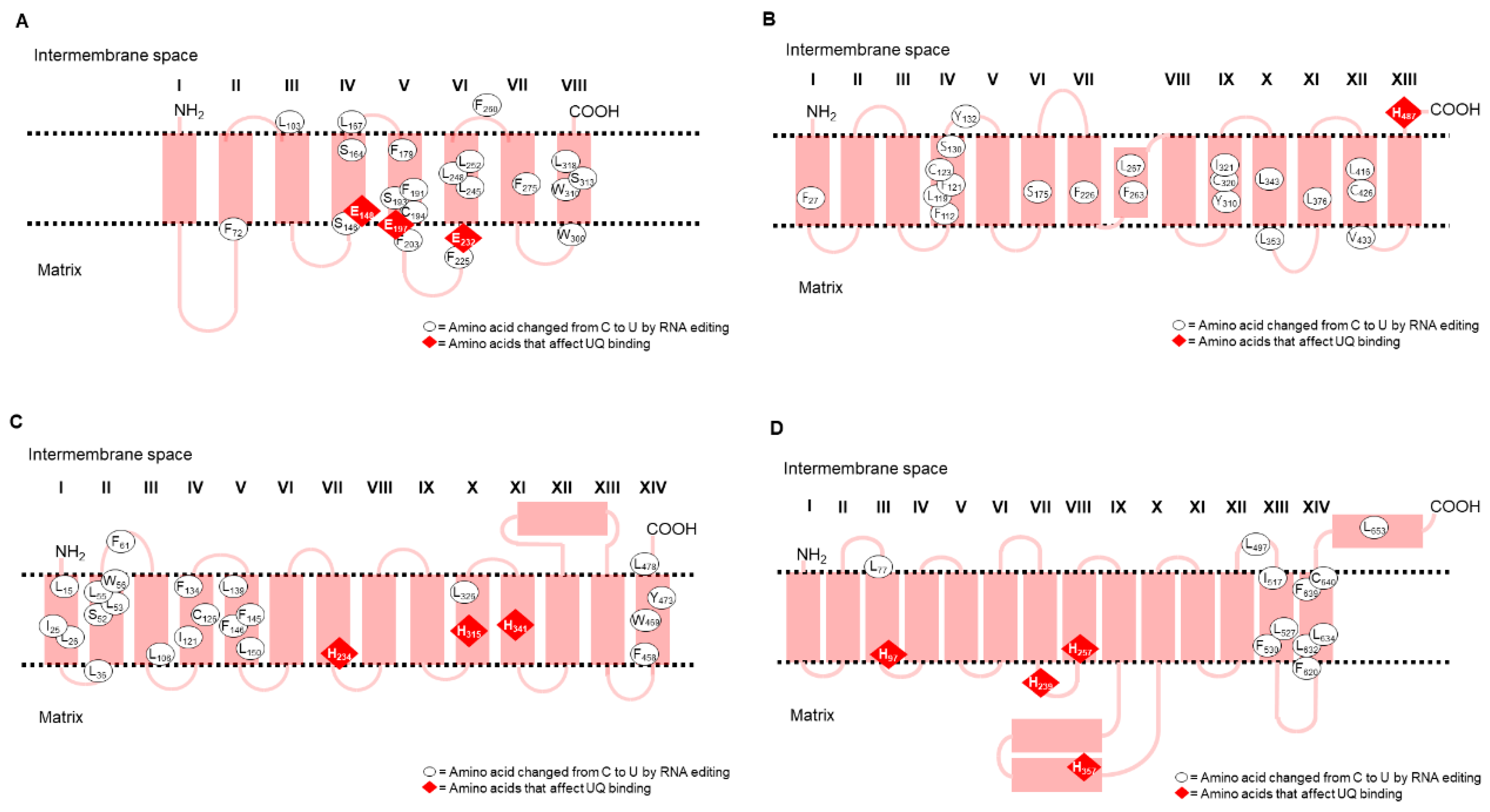
3.2. Transmembrane Domain Topology of Nad1, Nad2, Nad4, and Nad5
3.3. Allotopic Expression and Localization of Bamboo Nad1, Nad2, Nad4, and Nad5 in Yeast
3.4. Functional Characterization of the Bamboo Nad1, Nad2, Nad4, and Nad5 Subunits in Yeast Mitochondria
4. Materials and Methods
4.1. Plant Materials and Microbes
4.2. Preparation of Mitochondria from Young Bamboo Shoots
4.3. Isolation of Total Bamboo Mitochondrial RNA (mtRNA)
4.4. cDNA Synthesis by the Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
4.5. Sequencing and Alignment
4.6. Plasmid Construction
4.7. Yeast Transformation
4.8. Induction of MT-EGFP, MT-Nad1-EGFP, MT-Nad2-EGFP, MT-Nad4-EGFP, and MT-Nad5-EGFP in Yeast
4.9. Isolation of Total RNA from Yeast
4.10. Extraction of Yeast Total Proteins, Soluble Proteins, and Membrane Proteins
4.11. Measurement of the Protein Concentration
4.12. Fluorescence Detection
4.13. Western Blot Analysis
4.14. Isolation of Yeast Mitochondria
4.15. Influence of Salt or pH on the Growth of S. cerevisiae
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wikström, M. Two protons are pumped from the mitochondrial matrix per electron transferred between NADH and ubiquinone. FEBS Lett. 1984, 169, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Brandt, U. Energy converting NADH: Quinone oxidoreductase (complex I). Annu. Rev. Biochem. 2006, 75, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Yagi, T.; Matsuno-Yagi, A. The proton-translocating NADH-quinone oxidoreductase in the respiratory chain: The secret unlocked. Biochemistry 2003, 42, 2266–2274. [Google Scholar] [CrossRef] [PubMed]
- Vogel, R.O.; Smeitink, J.A.M.; Nijtmans, L.G.J. Human mitochondrial complex I assembly: A dynamic and versatile process. Biochim. Biophys. Acta 2007, 1767, 1215–1227. [Google Scholar] [CrossRef] [Green Version]
- Klodmann, J.; Sunderhaus, S.; Nimtz, M.; Jänsch, L.; Braun, H.-P. Internal architecture of mitochondrial complex I from Arabidopsis thaliana. Plant Cell 2010, 22, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subrahmanian, N.; Remacle, C.; Hamel, P.P. Plant mitochondrial Complex I composition and assembly: A review. Biochim. Biophys. Acta Bioenerg. 2016, 1857, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Senkler, J.; Senkler, M.; Braun, H.-P. Structure and function of complex I in animals and plants—A comparative view. Physiol. Plant 2017, 161, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Padavannil, A.; Zhou, L.; Guo, F.; Letts, J.A. Atomic structure of a mitochondrial complex I intermediate from vascular plants. eLife 2020, 9, e56664. [Google Scholar] [CrossRef] [PubMed]
- Grigorieff, N. Structure of the respiratory NADH:ubiquinone oxidoreductase (complex I). Curr. Opin. Struct. Biol. 1999, 9, 476–483. [Google Scholar] [CrossRef]
- Dudkina, N.V.; Eubel, H.; Keegstra, W.; Boekema, E.J.; Braun, H.-P. Structure of a mitochondrial supercomplex formed by respiratory chain complexes I and III. Proc. Natl. Acad. Sci. USA 2005, 102, 3225–3229. [Google Scholar] [CrossRef] [Green Version]
- Braun, H.-P.; Binder, S.; Brennicke, A.; Eubel, H.; Fernie, A.R.; Finkemeier, I.; Klodmann, J.; König, A.-C.; Kühn, K.; Meyer, E.; et al. The life of plant mitochondrial complex I. Mitochondrion 2014, 19, 295–313. [Google Scholar] [CrossRef] [PubMed]
- Soufari, H.; Parrot, C.; Kuhn, L.; Waltz, F.; Hashem, Y. Specific features and assembly of the plant mitochondrial complex I revealed by cryo-EM. Nat. Commun. 2020, 11, 5195. [Google Scholar] [CrossRef] [PubMed]
- Michel, J.; DeLeon-Rangel, J.; Zhu, S.; van Ree, K.; Vik, S.B. Mutagenesis of the L, M, and N subunits of complex I from Escherichia coli indicates a common role in function. PLoS ONE 2011, 6, e17420. [Google Scholar] [CrossRef]
- Röpke, M.; Saura, P.; Riepl, D.; Röverlein, M.C.; Kaila, V.R.I. Functional water wires catalyze long-range proton pumping in the mammalian respiratory complex I. J. Am. Chem. Soc. 2020, 142, 21758–21766. [Google Scholar] [CrossRef]
- Ogihara, Y.; Yamazaki, Y.; Murai, K.; Kanno, A.; Terachi, T.; Shiina, T.; Miyashita, N.; Nasuda, S.; Nakamura, C.; Mori, N.; et al. Structural dynamics of cereal mitochondrial genomes as revealed by complete nucleotide sequencing of the wheat mitochondrial genome. Nucleic Acids Res. 2005, 33, 6235–6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackenzie, S.A. Chapter 2 The unique biology of mitochondrial genome instability in plants. In Plant Mitochondria; Logan, D.C., Ed.; Annual Plant Reviews; Wiley-Blackwell: Oxford, UK, 2007; Volume 31, pp. 36–49. [Google Scholar]
- Huang, S.; Millar, A.H.; Taylor, N.L. Chapter 9 The plant mitochondrial proteome composition and stress response: Conservation and divergence between monocots and dicots. In Plant Mitochondria; Kempken, F., Ed.; Springer: Dordrecht, Netherlands, 2010; Volume 1, pp. 207–239. [Google Scholar]
- Notsu, Y.; Masood, S.; Nishikawa, T.; Kubo, N.; Akiduki, G.; Nakazono, M.; Hirai, A.; Kadowaki, K. The complete sequence of the rice (Oryza sativa L.) mitochondrial genome: Frequent DNA sequence acquisition and loss during the evolution of flowering plants. Mol. Genet. Genom. 2002, 268, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Clifton, S.W.; Minx, P.; Fauron, C.M.; Gibson, M.; Allen, J.O.; Sun, H.; Thompson, M.; Barbazuk, W.B.; Kanuganti, S.; Tayloe, C.; et al. Sequence and comparative analysis of the maize NB mitochondrial genome. Plant Physiol. 2004, 136, 3486–3503. [Google Scholar] [CrossRef] [Green Version]
- Lin, N.S.; Hu, C.C.; Chien, L.F.; Hsu, Y.H. A Mitochondrial Genome Sequence of Bamboo and Comparison to Those of Cereals. 2007. Available online: https://www.ncbi.nlm.nih.gov/nuccore/164422210/ (accessed on 4 February 2022).
- Gott, J.M.; Emeson, R.B. Functions and mechanisms of RNA editing. Annu. Rev. Genet. 2000, 34, 499–531. [Google Scholar] [CrossRef]
- Gray, M.W. RNA editing in plant mitochondria: 20 years later. IUBMB Life 2009, 61, 1101–1104. [Google Scholar] [CrossRef]
- Maldonado, M.; Abe, K.M.; Letts, J.A. A structural perspective on the RNA editing of plant respiratory complexes. Int. J. Mol. Sci. 2022, 23, 684. [Google Scholar] [CrossRef]
- Howell, N.; Bindoff, L.A.; McCullough, D.A.; Kubacka, I.; Poulton, J.; Mackey, D.; Taylor, L.; Turnbull, D.M. Leber hereditary optic neuropathy: Identification of the same mitochondrial ND1 mutation in six pedigrees. Am. J. Hum. Genet. 1991, 49, 939–950. [Google Scholar] [PubMed]
- Lertrit, P.; Noer, A.S.; Jean-Francois, M.J.; Kapsa, R.; Dennett, X.; Thyagarajan, D.; Lethlean, K.; Byrne, E.; Marzuki, S. A new disease-related mutation for mitochondrial encephalopathy lactic acidosis and strokelike episodes (MELAS) syndrome affects the ND4 subunit of the respiratory complex I. Am. J. Hum. Genet. 1992, 51, 457–468. [Google Scholar] [PubMed]
- Houštek, J.; Hejzlarová, K.; Vrbacký, M.; Drahota, Z.; Landa, V.; Zídek, V.; Mlejnek, P.; Šimáková, M.; Šilhavy, J.; Mikšík, I.; et al. Nonsynonymous variants in mt-Nd2, mt-Nd4, and mt-Nd5 are linked to effects on oxidative phosphorylation and insulin sensitivity in rat conplastic strains. Physiol. Genom. 2012, 44, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Longevialle, A.F.; Meyer, E.H.; Andres, C.; Taylor, N.L.; Lurin, C.; Millar, A.H.; Small, I.D. The pentatricopeptide repeat gene OTP43 is required for trans-splicing of the mitochondrial nad1 intron 1 in Arabidopsis thaliana. Plant Cell 2007, 19, 3256–3265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vgenopoulou, I.; Gemperli, A.C.; Steuber, J. Specific modification of a Na+ binding site in NADH:quinone oxidoreductase from Klebsiella pneumoniae with dicyclohexylcarbodiimide. J. Bacteriol. 2006, 188, 3264–3272. [Google Scholar] [CrossRef] [Green Version]
- Yoga, E.G.; Schiller, J.; Zickermann, V. Ubiquinone binding and reduction by complex I—Open questions and mechanistic implications. Front. Chem. 2021, 9, 672851. [Google Scholar] [CrossRef]
- Sato, M.; Sinha, P.K.; Torres-Bacete, J.; Matsuno-Yagi, A.; Yagi, T. Energy transducing roles of antiporter-like subunits in Escherichia coli NDH-1 with main focus on subunit NuoN (ND2). J. Biol. Chem. 2013, 288, 24705–24716. [Google Scholar] [CrossRef] [Green Version]
- Brangeon, J.; Sabar, M.; Gutierres, S.; Combettes, B.; Bove, J.; Gendy, C.; Chetrit, P.; Des Francs-Small, C.C.; Pla, M.; Vedel, F.; et al. Defective splicing of the first nad4 intron is associated with lack of several complex I subunits in the Nicotiana sylvestris NMS1 nuclear mutant. Plant J. 2000, 21, 269–280. [Google Scholar] [CrossRef]
- Sabar, M.; de Paepe, R.; de Kouchkovsky, Y. Complex I impairment, respiratory compensations, and photosynthetic decrease in nuclear and mitochondrial male sterile mutants of Nicotiana sylvestris. Plant Physiol. 2000, 124, 1239–1250. [Google Scholar] [CrossRef] [Green Version]
- Mathiesen, C.; Hägerhäll, C. Transmembrane topology of the NuoL, M and N subunits of NADH:quinone oxidoreductase and their homologues among membrane-bound hydrogenases and bona fide antiporters. Biochim. Biophys. Acta 2002, 1556, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Torres-Bacete, J.; Sinha, P.K.; Matsuno-Yagi, A.; Yagi, T. Structural contribution of the C-terminal segments of NuoL (ND5) and NuoM (ND4) subunits of complex I from Escherichia coli. J. Biol. Chem. 2011, 286, 34007–34014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steimle, S.; Bajzath, C.; Dorner, K.; Schulte, M.; Bothe, V.; Friedrich, T. Role of subunit NuoL for proton translocation by respiratory complex I. Biochemistry 2011, 50, 3386–3393. [Google Scholar] [CrossRef]
- Omura, T. Mitochondria-targeting sequence, a multi-role sorting sequence recognized at all steps of protein import into mitochondria. J. Biochem. 1998, 123, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Roise, D.; Schatz, G. Mitochondrial presequences. J. Biol. Chem. 1988, 263, 4509–4511. [Google Scholar] [CrossRef]
- Giraud, M.F.; Velours, J. ATP synthase of yeast mitochondria. Isolation of the F1 delta subunit, sequence and disruption of the structural gene. Eur. J. Biochem. 1994, 222, 851–859. [Google Scholar] [CrossRef]
- Steffen, W.; Gemperli, A.C.; Cvetesic, N.; Steuber, J. Organelle-specific expression of subunit ND5 of human complex I (NADH dehydrogenase) alters cation homeostasis in Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 648–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosek, J.; Fukuhara, H. NADH dehydrogenase subunit genes in the mitochondrial DNA of yeasts. J. Bacteriol. 1994, 176, 5622–5630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, S.; Grivell, L.A. Purification and characterization of a rotenone-insensitive NADH:Q6 oxidoreductase from mitochondria of Saccharomyces cerevisiae. Eur. J. Biochem. 1988, 176, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Luttik, M.A.; Overkamp, K.M.; Kötter, P.; de Vries, S.; van Dijken, J.P.; Pronk, J.T. The Saccharomyces cerevisiae NDE1 and NDE2 genes encode separate mitochondrial NADH dehydrogenases catalyzing the oxidation of cytosolic NADH. J. Biol. Chem. 1998, 273, 24529–24534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, B.M.; Overkamp, K.M.; van Maris, A.J.A.; Kötter, P.; Luttik, M.A.; van Dijken, J.P.; Pronk, J.T. Stoichiometry and compartmentation of NADH metabolism in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.M.; Bandeiras, T.M.; Teixeira, M. New insights into type II NAD(P)H:quinone oxidoreductases. Microbiol. Mol. Biol. Rev. 2004, 68, 603–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foury, F.; Roganti, T.; Lecrenier, N.; Purnelle, B. The complete sequence of the mitochondrial genome of Saccharomyces cerevisiae. FEBS Lett. 1998, 440, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, B.; Hennig, B.; Köhler, H.; Neupert, W. Transport of the precursor to neurospora ATPase subunit 9 into yeast mitochondria. Implications on the diversity of the transport mechanism. J. Biol. Chem. 1983, 258, 4687–4689. [Google Scholar] [CrossRef]
- Chien, L.F.; Wu, Y.C.; Chen, H.P. Mitochondrial energy metabolism in young bamboo rhizomes from Bambusa oldhamii and Phyllostachys edulis during shooting stage. Plant Physiol. Biochem. 2011, 49, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Kurki, S.; Zickermann, V.; Kervinen, M.; Hassinen, I.; Finel, M. Mutagenesis of three conserved Glu residues in a bacterial homologue of the ND1 subunit of complex I affects ubiquinone reduction kinetics but not inhibition by dicyclohexylcarbodiimide. Biochemistry 2000, 39, 13496–13502. [Google Scholar] [CrossRef]
- Torres-Bacete, J.; Sinha, P.K.; Castro-Guerrero, N.; Matsuno-Yagi, A.; Yagi, T. Features of subunit NuoM (ND4) in Escherichia coli NDH-1: Topology and implication of conserved glu144 for coupling site 1. J. Biol. Chem. 2009, 284, 33062–33069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolender, N.; Sickmann, A.; Wagner, R.; Meisinger, C.; Pfanner, N. Multiple pathways for sorting mitochondrial precursor proteins. EMBO Rep. 2008, 9, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galkin, A. Water route for proton pumping in mitochondrial complex I. Nat. Struct. Mol. Biol. 2020, 27, 1–3. [Google Scholar] [CrossRef]
- Efremov, R.G.; Baradaran, R.; Sazanov, L.A. The architecture of respiratory complex I. Nature 2010, 465, 441–445. [Google Scholar] [CrossRef]
- Treberg, J.R.; Brand, M.D. A model of the proton translocation mechanism of complex I. J. Biol. Chem. 2011, 286, 17579–17584. [Google Scholar] [CrossRef] [Green Version]
- Parey, K.; Lasham, J.; Mills, D.J.; Djurabekova, A.; Haapanen, O.; Yoga, E.G.; Xie, H.; Kühlbrandt, W.; Sharma, V.; Vonck, J.; et al. High-resolution structure and dynamics of mitochondrial complex I-Insights into the proton pumping mechanism. Sci. Adv. 2021, 7, eabj3221. [Google Scholar] [CrossRef]
- Nakamaru-Ogiso, E.; Kao, M.C.; Chen, H.; Sinha, S.C.; Yagi, T.; Ohnishi, T. The membrane subunit NuoL(ND5) is involved in the indirect proton pumping mechanism of Escherichia coli complex I. J. Biol. Chem. 2010, 285, 39070–39078. [Google Scholar] [CrossRef] [Green Version]
- Cui, P.; Liu, H.; Lin, Q.; Ding, F.; Zhuo, G.; Hu, S.; Liu, D.; Yang, W.; Zhan, K.; Zhang, A.; et al. A complete mitochondrial genome of wheat (Triticum aestivum cv. Chinese Yumai), and fast evolving mitochondrial genes in higher plants. J. Genet. 2009, 88, 299–307. [Google Scholar] [CrossRef]
- Morawala-Patell, V.; Gualberto, J.M.; Lamattina, L.; Grienenberger, J.M.; Bonnard, G. Cis- and trans-splicing and RNA editing are required for the expression of nad2 in wheat mitochondria. Mol. Gen. Genet. 1998, 258, 503–511. [Google Scholar] [CrossRef]
- Christensen, A.C.; Lyznik, A.; Mohammed, S.; Elowsky, C.G.; Elo, A.; Yule, R.; Mackenzie, S.A. Dual-domain, dual-targeting organellar protein presequences in Arabidopsis can use non-AUG start codons. Plant Cell 2005, 17, 2805–2816. [Google Scholar] [CrossRef] [Green Version]
- Edqvist, J.; Burger, G.; Gray, M.W. Expression of mitochondrial protein-coding genes in Tetrahymena pyriformis. J. Mol. Biol. 2000, 297, 381–393. [Google Scholar] [CrossRef]
- Mulligan, R.M.; Chang, K.L.; Chou, C.C. Computational analysis of RNA editing sites in plant mitochondrial genomes reveals similar information content and a sporadic distribution of editing sites. Mol. Biol. Evol. 2007, 24, 1971–1981. [Google Scholar] [CrossRef] [Green Version]
- Gualberto, J.M.; Lamattina, L.; Bonnard, G.; Weil, J.H.; Grienenberger, J.M. RNA editing in wheat mitochondria results in the conservation of protein sequences. Nature 1989, 341, 660–662. [Google Scholar] [CrossRef] [PubMed]
- Herrnstadt, C.; Elson, J.L.; Fahy, E.; Preston, G.; Turnbull, D.M.; Anderson, C.; Ghosh, S.S.; Olefsky, J.M.; Beal, M.F.; Davis, R.E.; et al. Reduced-median-network analysis of complete mitochondrial DNA coding-region sequences for the major African, Asian, and European haplogroups. Am. J. Hum. Genet. 2002, 70, 1152–1171. [Google Scholar] [CrossRef] [Green Version]
- Murai, M.; Ishihara, A.; Nishioka, T.; Yagi, T.; Miyoshi, H. The ND1 subunit constructs the inhibitor binding domain in bovine heart mitochondrial complex I. Biochemistry 2007, 46, 6409–6416. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Hroudová, J.; Van Bergen, N.J.; Lopez Sanchez, M.I.; Trounce, I.A.; McKenzie, M. Loss of mitochondrial DNA-encoded protein ND1 results in disruption of complex I biogenesis during early stages of assembly. FASEB J. 2016, 30, 2236–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tursun, A.; Zhu, S.; Vik, S.B. Probing the proton channels in subunit N of Complex I from Escherichia coli through intra-subunit cross-linking. Biochim. Biophys. Acta 2016, 1857, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.F.; Guo, Z.H.; Li, D.Z. Rapid sequencing of the bamboo mitochondrial genome using Illumina technology and parallel episodic evolution of organelle genomes in grasses. PLoS ONE 2012, 7, e30297. [Google Scholar]
- Albracht, S.P.; Hedderich, R. Learning from hydrogenases: Location of a proton pump and of a second FMN in bovine NADH-ubiquinone oxidoreductase (Complex I). FEBS Lett. 2000, 485, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, T.; Böttcher, B. The gross structure of the respiratory complex I: A Lego System. Biochim. Biophys. Acta 2004, 1608, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nakamaru-Ogiso, E.; Sakamoto, K.; Matsuno-Yagi, A.; Miyoshi, H.; Yagi, T. The ND5 subunit was labeled by a photoaffinity analogue of fenpyroximate in bovine mitochondrial complex I. Biochemistry 2003, 42, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Perales-Clemente, E.; Fernández-Silva, P.; Acín-Pérez, R.; Pérez-Martos, A.; Enríquez, J.A. Allotopic expression of mitochondrial-encoded genes in mammals: Achieved goal, undemonstrated mechanism or impossible task? Nucleic Acids Res. 2011, 39, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, C.; Augustin, S.; Ellouze, S.; Bénit, P.; Bouaita, A.; Rustin, P.; Sahel, J.A.; Corral-Debrinski, M. The optimized allotopic expression of ND1 or ND4 genes restores respiratory chain complex I activity in fibroblasts harboring mutations in these genes. Biochim. Biophys. Acta. 2008, 1783, 1707–1717. [Google Scholar] [CrossRef] [Green Version]
- Ellouze, S.; Augustin, S.; Bouaita, A.; Bonnet, C.; Simonutti, M.; Forster, V.; Picaud, S.; Sahel, J.A.; Corral-Debrinski1, M. Optimized allotopic expression of the human mitochondrial ND4 prevents blindness in a rat model of mitochondrial dysfunction. Am. J. Hum. Genet. 2008, 83, 373–387. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.; Hulett, J.M.; Lithgow, T.; Whelan, J. Protein import into mitochondria: Origins and functions today (review). Mol. Membr. Biol. 2005, 22, 87–100. [Google Scholar] [CrossRef] [PubMed]
- van der Laan, M.; Hutu, D.P.; Rehling, P. On the mechanism of preprotein import by the mitochondrial presequence translocase. Biochim. Biophys. Acta 2010, 1803, 732–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaglesfield, R.; Tokatlidis, K. Targeting and insertion of membrane proteins in mitochondria. Front. Cell Dev. Biol. 2021, 9, 803205. [Google Scholar] [CrossRef] [PubMed]
- Cardol, P.; Matagne, R.F.; Remacle, C. Impact of mutations affecting ND mitochondria-encoded subunits on the activity and assembly of complex I in Chlamydomonas. Implication for the structural organization of the enzyme. J. Mol. Biol. 2002, 319, 1211–1221. [Google Scholar] [CrossRef]
- Juszczuk, I.M.; Szal, B.; Rychter, A.M. Oxidation-reduction and reactive oxygen species homeostasis in mutant plants with respiratory chain complex I dysfunction. Plant Cell Environ. 2012, 35, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Marienfeld, J.R.; Newton, K.J. The maize NCS2 abnormal growth mutant has a chimeric nad4–nad7 mitochondrial gene and is associated with reduced complex I function. Genetics 1994, 138, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Steimle, S.; Willistein, M.; Hegger, P.; Janoschke, M.; Erhardt, H.; Friedrich, T. Asp563 of the horizontal helix of subunit NuoL is involved in proton translocation by the respiratory complex I. FEBS Lett. 2012, 586, 699–704. [Google Scholar] [CrossRef]
- Ohnishi, T. Piston drives a proton pump. Nature 2010, 465, 428–429. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar]
- Gietz, D.; St Jean, A.; Woods, R.A.; Schiestl, R.H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Res. 1992, 20, 1425. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.E.; Brown, T.A.; Trumpower, B.L. A rapid and simple method for preparation of RNA from Saccharomyces cerevisiae. Nucleic Acids Res. 1990, 18, 3091–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Meisinger, C.; Pfanner, N.; Truscott, K.N. Isolation of yeast mitochondria. Methods Mol. Biol. 2006, 133, 33–39. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, H.-C.; Hsieh, C.-H.; Hsu, C.-W.; Hsu, Y.-H.; Chien, L.-F. Cloning and Organelle Expression of Bamboo Mitochondrial Complex I Subunits Nad1, Nad2, Nad4, and Nad5 in the Yeast Saccharomyces cerevisiae. Int. J. Mol. Sci. 2022, 23, 4054. https://doi.org/10.3390/ijms23074054
Tsai H-C, Hsieh C-H, Hsu C-W, Hsu Y-H, Chien L-F. Cloning and Organelle Expression of Bamboo Mitochondrial Complex I Subunits Nad1, Nad2, Nad4, and Nad5 in the Yeast Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2022; 23(7):4054. https://doi.org/10.3390/ijms23074054
Chicago/Turabian StyleTsai, Hsieh-Chin, Cheng-Hung Hsieh, Ching-Wen Hsu, Yau-Heiu Hsu, and Lee-Feng Chien. 2022. "Cloning and Organelle Expression of Bamboo Mitochondrial Complex I Subunits Nad1, Nad2, Nad4, and Nad5 in the Yeast Saccharomyces cerevisiae" International Journal of Molecular Sciences 23, no. 7: 4054. https://doi.org/10.3390/ijms23074054