
Validation and Data-Integration of Yeast-Based Assays for Functional Classification of BRCA1 Missense Variants

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
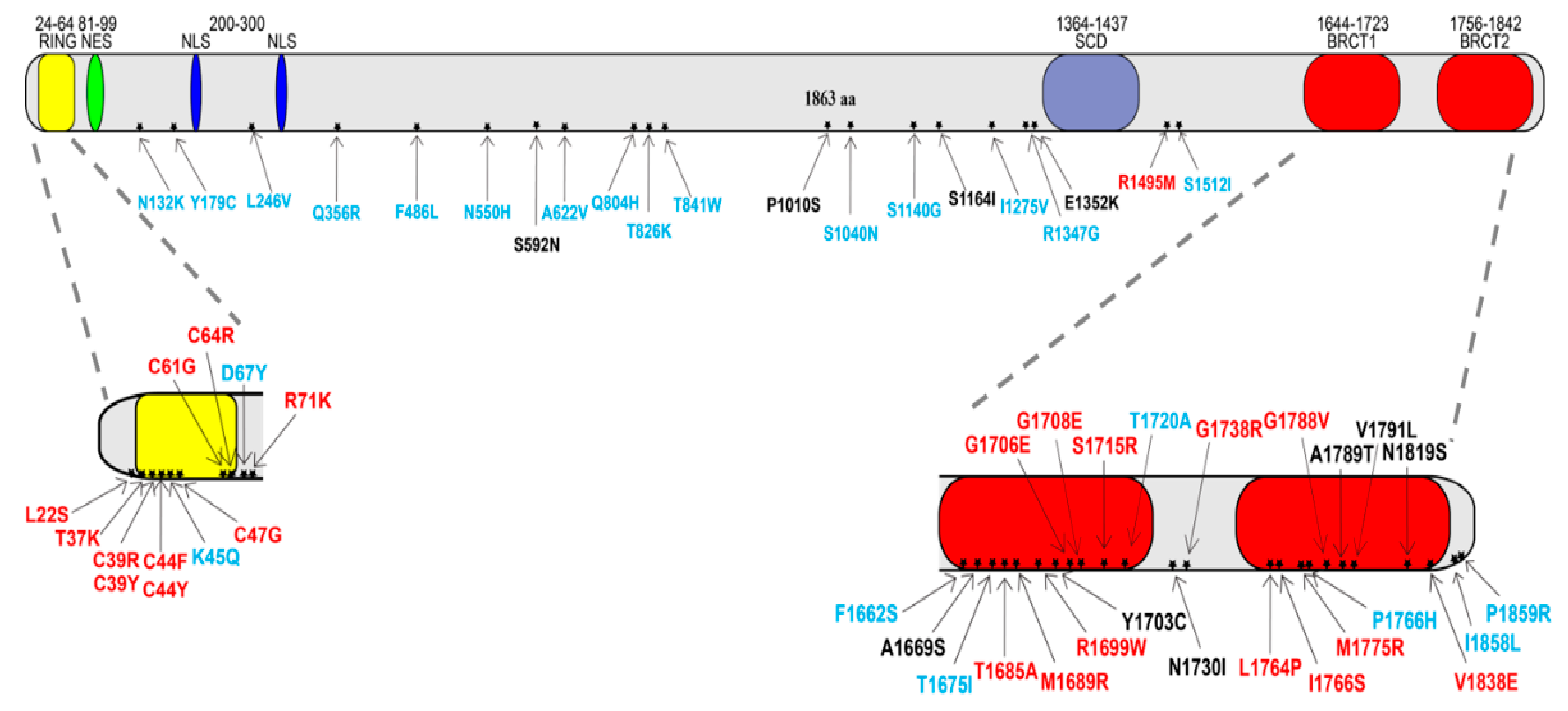
2.1. BRCA1 Missense Variants’ Classification
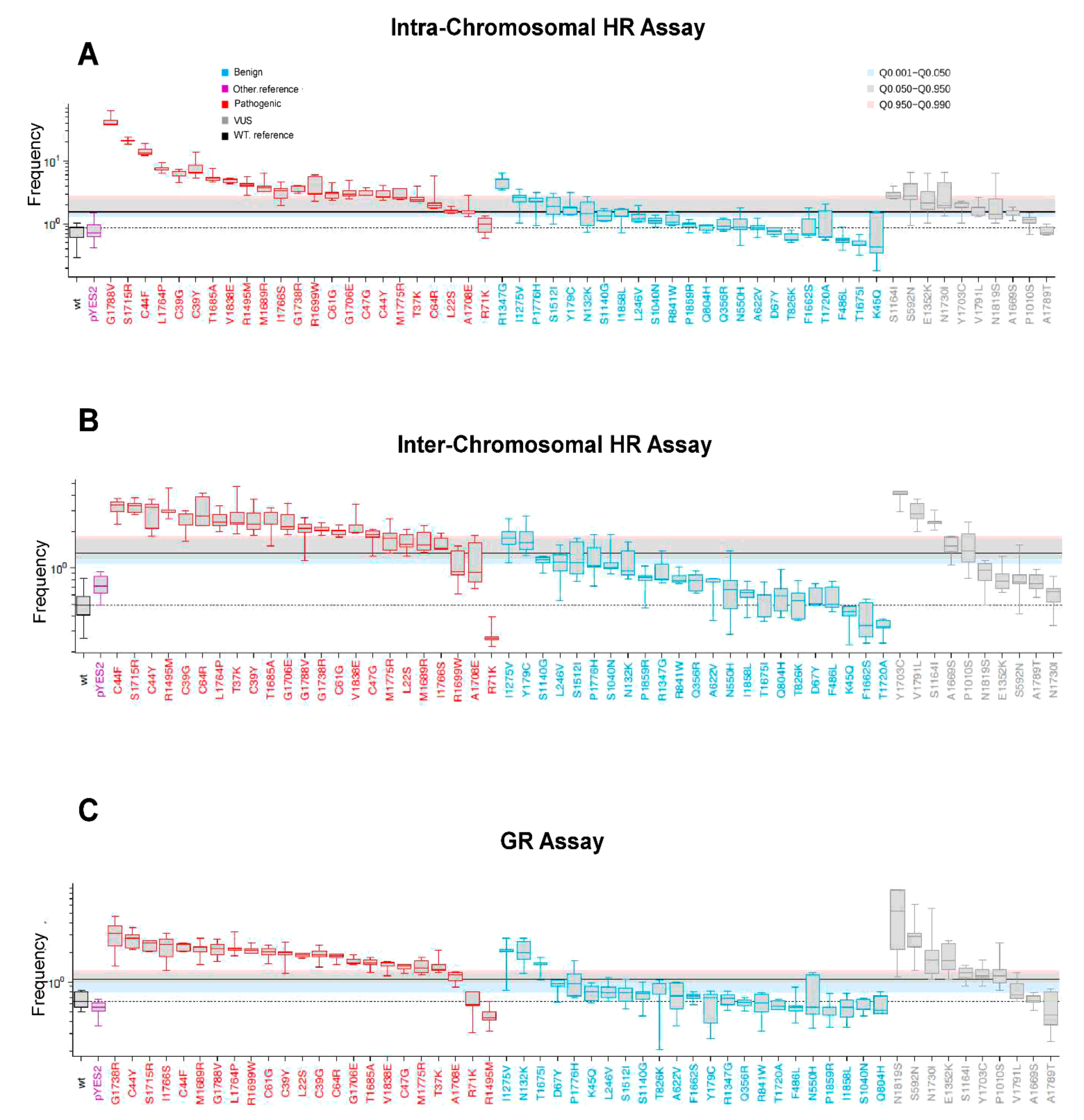
2.2. BRCA1 Pathogenic Variants Increased HR and GR in Yeast
2.3. Effect of VUS on HR and GR in Yeast
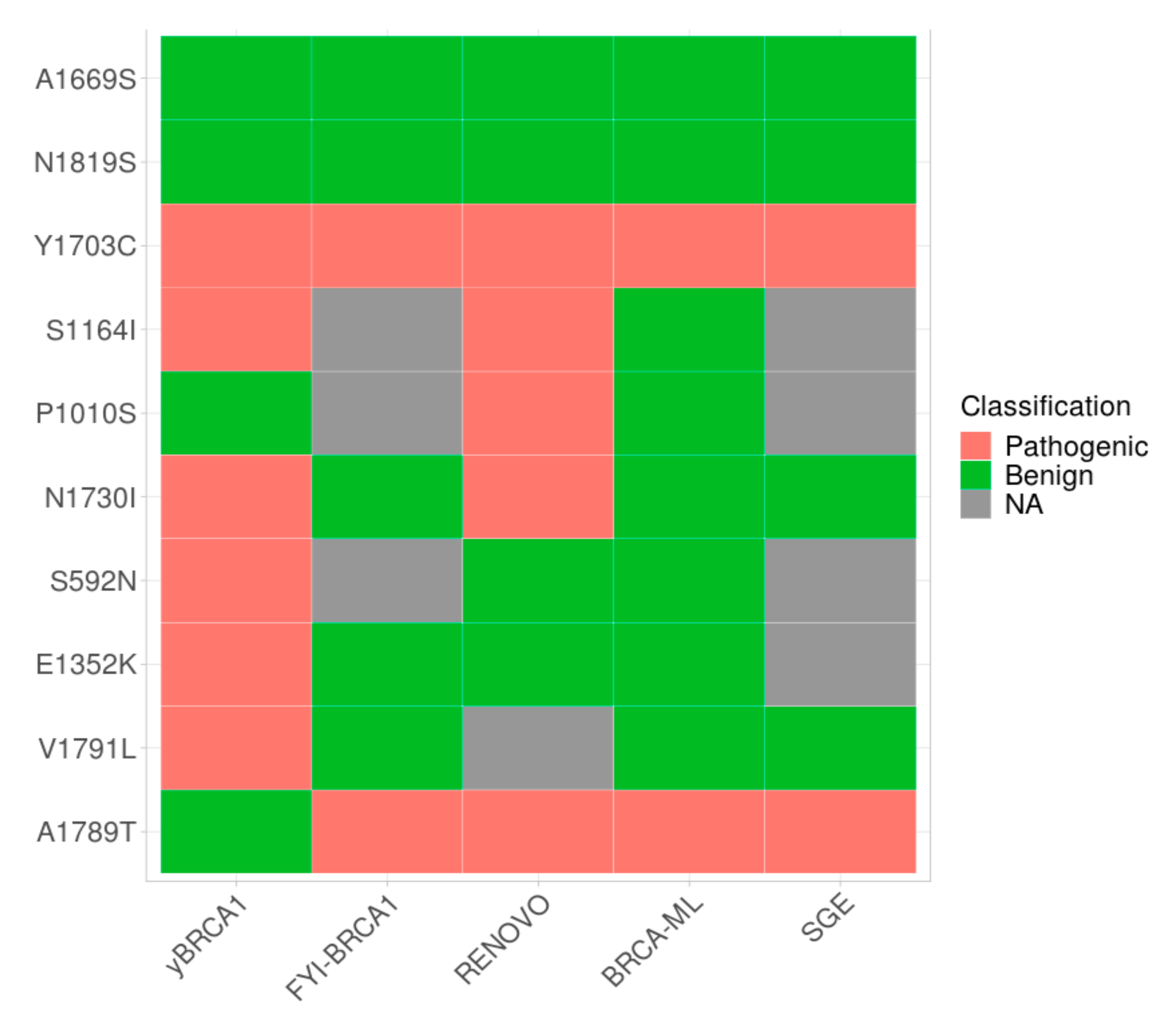
2.4. Development of yBRCA1: A Classifier Combination Approach for Functional Characterization
2.5. VUS Functional Characterization by the Classifier Combination Method yBRCA1
3. Discussion
4. Materials and Methods
4.1. BRCA1 Variant Selection
4.2. Yeast Strains
4.3. Protein Extraction and Western Blotting
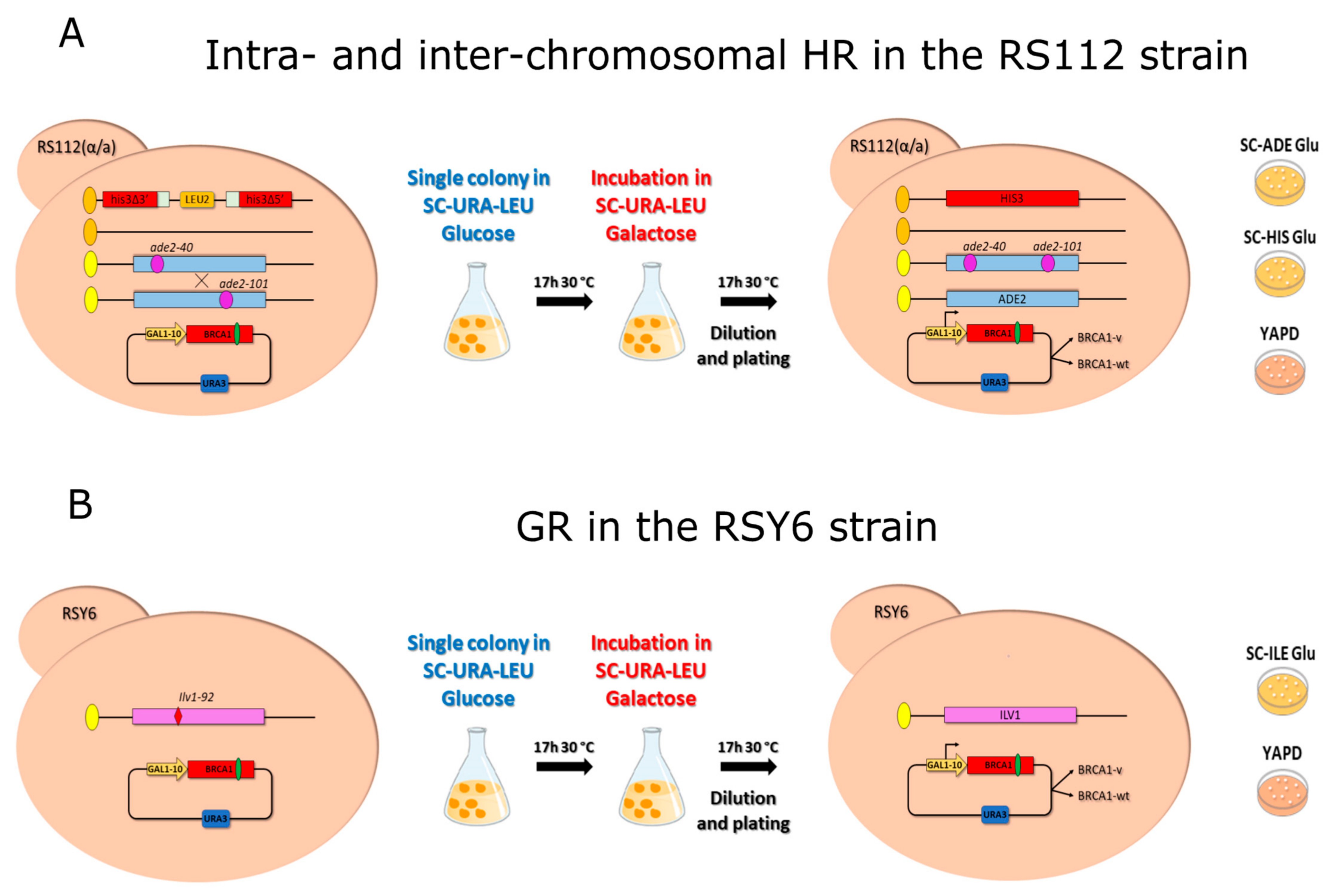
4.4. Homologous Recombination and Gene Reversion Assays
4.5. Small Colony Phenotype Assay
4.6. Best Cut-Off Calculation and Integration of Datasets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Narod, S.A.; Foulkes, W.D. BRCA1 and BRCA2: 1994 and beyond. Nat. Rev. Cancer 2004, 4, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, F.C.; van Overeem Hansen, T.; Sørensen, C.S. Hereditary breast and ovarian cancer: New genes in confined pathways. Nat. Rev. Cancer 2016, 16, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.N.; Kachnic, L.A. Roles of BRCA1 and BRCA2 in homologous recombination, DNA replication fidelity and the cellular response to ionizing radiation. Oncogene 2003, 22, 5784–5791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, M.T.; Tudini, E.; Li, H.; Hahnen, E.; Wappenschmidt, B.; Feliubadalo, L.; Aalfs, C.M.; Agata, S.; Aittomaki, K.; Alducci, E.; et al. Large scale multifactorial likelihood quantitative analysis of BRCA1 and BRCA2 variants: An ENIGMA resource to support clinical variant classification. Hum Mutat. 2019, 40, 1557–1578. [Google Scholar] [CrossRef] [Green Version]
- Starita, L.M.; Ahituv, N.; Dunham, M.J.; Kitzman, J.O.; Roth, F.P.; Seelig, G.; Shendure, J.; Fowler, D.M. Variant Interpretation: Functional Assays to the Rescue. Am. J. Hum. Genet. 2017, 101, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Cline, M.S.; Liao, R.G.; Parsons, M.T.; Paten, B.; Alquaddoomi, F.; Antoniou, A.; Baxter, S.; Brody, L.; Cook-Deegan, R.; Coffin, A.; et al. BRCA Challenge: BRCA Exchange as a global resource for variants in BRCA1 and BRCA2. PLoS Genet. 2018, 14, e1007752. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, R.; Oak, N.; Plon, S.E. Evaluation of in silico algorithms for use with ACMG/AMP clinical variant interpretation guidelines. Genome Biol. 2017, 18, 225. [Google Scholar] [CrossRef] [Green Version]
- Cubuk, C.; Garrett, A.; Choi, S.; King, L.; Loveday, C.; Torr, B.; Burghel, G.J.; Durkie, M.; Callaway, A.; Robinson, R.; et al. Clinical likelihood ratios and balanced accuracy for 44 in silico tools against multiple large-scale functional assays of cancer susceptibility genes. Genet. Med. 2021, 23, 2096–2104. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Kanavy, D.M.; McNulty, S.M.; Jairath, M.K.; Brnich, S.E.; Bizon, C.; Powell, B.C.; Berg, J.S. Comparative analysis of functional assay evidence use by ClinGen Variant Curation Expert Panels. Genome Med. 2019, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Brnich, S.E.; Abou Tayoun, A.N.; Couch, F.J.; Cutting, G.R.; Greenblatt, M.S.; Heinen, C.D.; Kanavy, D.M.; Luo, X.; McNulty, S.M.; Starita, L.M.; et al. Recommendations for application of the functional evidence PS3/BS3 criterion using the ACMG/AMP sequence variant interpretation framework. Genome Med. 2020, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.N.; Bouwman, P.; Kousholt, A.N.; Eccles, D.M.; Millot, G.A.; Masson, J.-Y.; Schmidt, M.K.; Sharan, S.K.; Scully, R.; Wiesmüller, L.; et al. Variants of uncertain clinical significance in hereditary breast and ovarian cancer genes: Best practices in functional analysis for clinical annotation. J. Med. Genet. 2020, 57, 509–518. [Google Scholar] [CrossRef]
- Millot, G.A.; Carvalho, M.A.; Caputo, S.M.; Vreeswijk, M.P.G.; Brown, M.A.; Webb, M.; Rouleau, E.; Neuhausen, S.L.; Hansen, T.v.O.; Galli, A.; et al. A guide for functional analysis of BRCA1 variants of uncertain significance. Hum. Mutat. 2012, 33, 1526–1537. [Google Scholar] [CrossRef] [Green Version]
- Findlay, G.M.; Daza, R.M.; Martin, B.; Zhang, M.D.; Leith, A.P.; Gasperini, M.; Janizek, J.D.; Huang, X.; Starita, L.M.; Shendure, J. Accurate classification of BRCA1 variants with saturation genome editing. Nature 2018, 562, 217–222. [Google Scholar] [CrossRef]
- Starita, L.M.; Islam, M.M.; Banerjee, T.; Adamovich, A.I.; Gullingsrud, J.; Fields, S.; Shendure, J.; Parvin, J.D. A Multiplex Homology-Directed DNA Repair Assay Reveals the Impact of More Than 1000 BRCA1 Missense Substitution Variants on Protein Function. Am. J. Hum. Genet. 2018, 103, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Jhuraney, A.; Velkova, A.; Johnson, R.C.; Kessing, B.; Carvalho, R.S.; Whiley, P.; Spurdle, A.B.; Vreeswijk, M.P.; Caputo, S.M.; Millot, G.A.; et al. BRCA1 Circos: A visualisation resource for functional analysis of missense variants. J. Med. Genet. 2015, 52, 224–230. [Google Scholar] [CrossRef]
- Lyra, P.C.M., Jr.; Nepomuceno, T.C.; de Souza, M.L.M.; Machado, G.F.; Veloso, M.F.; Henriques, T.B.; Dos Santos, D.Z.; Ribeiro, I.G.; Ribeiro, R.S., Jr.; Rangel, L.B.A.; et al. Integration of functional assay data results provides strong evidence for classification of hundreds of BRCA1 variants of uncertain significance. Genet. Med. 2021, 23, 306–315. [Google Scholar] [CrossRef]
- Caligo, M.A.; Bonatti, F.; Guidugli, L.; Aretini, P.; Galli, A. A yeast recombination assay to characterize human BRCA1 missense variants of unknown pathological significance. Hum. Mutat. 2009, 30, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Lodovichi, S.; Bellè, F.; Cervelli, T.; Lorenzoni, A.; Maresca, L.; Cozzani, C.; Caligo, M.A.; Galli, A. Effect of BRCA1 missense variants on gene reversion in DNA double-strand break repair mutants and cell cycle-Arrested cells of Saccharomyces cerevisiae. Mutagenesis 2020, 35, 189–195. [Google Scholar] [CrossRef]
- Maresca, L.; Lodovichi, S.; Lorenzoni, A.; Cervelli, T.; Monaco, R.; Spugnesi, L.; Tancredi, M.; Falaschi, E.; Zavaglia, K.; Landucci, E.; et al. Functional Interaction Between BRCA1 and DNA Repair in Yeast May Uncover a Role of RAD50, RAD51, MRE11A, and MSH6 Somatic Variants in Cancer Development. Front. Genet. 2018, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Maresca, L.; Spugnesi, L.; Lodovichi, S.; Cozzani, C.; Naccarato, A.G.; Tancredi, M.; Collavoli, A.; Falaschi, E.; Rossetti, E.; Aretini, P.; et al. MSH2 role in BRCA1-driven tumorigenesis: A preliminary study in yeast and in human tumors from BRCA1-VUS carriers. Eur. J. Med. Genet. 2015, 58, 531–539. [Google Scholar] [CrossRef]
- Lodovichi, S.; Vitello, M.; Cervelli, T.; Galli, A. Expression of cancer related BRCA1 missense variants decreases MMS-induced recombination in Saccharomyces cerevisiae without altering its nuclear localization. Cell Cycle 2016, 15, 2723–2731. [Google Scholar] [CrossRef] [Green Version]
- Thouvenot, P.; Ben Yamin, B.; Fourrière, L.; Lescure, A.; Boudier, T.; Del Nery, E.; Chauchereau, A.; Goldgar, D.E.; Houdayer, C.; Stoppa-Lyonnet, D.; et al. Functional Assessment of Genetic Variants with Outcomes Adapted to Clinical Decision-Making. PLoS Genet. 2016, 12, e1006096. [Google Scholar] [CrossRef] [Green Version]
- Mohandes, M.; Deriche, M.; Aliyu, S.O. Classifiers combination techniques: A comprehensive review. IEEEAccess 2018, 6, 19626–19639. [Google Scholar] [CrossRef]
- Favalli, V.; Tini, G.; Bonetti, E.; Vozza, G.; Guida, A.; Gandini, S.; Pelicci, P.G.; Mazzarella, L. Machine learning-based reclassification of germline variants of unknown significance: The RENOVO algorithm. Am. J. Hum. Genet. 2021, 108, 682–695. [Google Scholar] [CrossRef]
- Hart, S.N.; Polley, E.C.; Shimelis, H.; Yadav, S.; Couch, F.J. Prediction of the functional impact of missense variants in BRCA1 and BRCA2 with BRCA-ML. NPJ Breast Cancer 2020, 6, 13. [Google Scholar] [CrossRef]
- Poon, K.S. In silico analysis of BRCA1 and BRCA2 missense variants and the relevance in molecular genetic testing. Sci. Rep. 2021, 11, 11114. [Google Scholar] [CrossRef]
- Woods, N.T.; Baskin, R.; Golubeva, V.; Jhuraney, A.; De-Gregoriis, G.; Vaclova, T.; Goldgar, D.E.; Couch, F.J.; Carvalho, M.A.; Iversen, E.S.; et al. Functional assays provide a robust tool for the clinical annotation of genetic variants of uncertain significance. NPJ Genom. Med. 2016, 1, 16001. [Google Scholar] [CrossRef] [Green Version]
- Toland, A.E.; Andreassen, P.R. DNA repair-related functional assays for the classification of BRCA1 and BRCA2 variants: A critical review and needs assessment. J. Med. Genet. 2017, 54, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Bouwman, P.; van der Heijden, I.; van der Gulden, H.; de Bruijn, R.; Braspenning, M.E.; Moghadasi, S.; Wessels, L.F.A.; Vreeswijk, M.P.G.; Jonkers, J. Functional Categorization of BRCA1 Variants of Uncertain Clinical Significance in Homologous Recombination Repair Complementation Assays. Clin. Cancer Res. 2020, 26, 4559–4568. [Google Scholar] [CrossRef]
- Cervelli, T.; Lodovichi, S.; Belle, F.; Galli, A. Yeast-based assays for the functional characterization of cancer-associated variants of human DNA repair genes. Microb. Cell 2020, 7, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, T.; Galli, A. Yeast as a Tool to Understand the Significance of Human Disease-Associated Gene Variants. Genes 2021, 12, 1303. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.B.; Westmoreland, T.J.; Verrier, C.S.; Blanchette, C.A.; Sabin, T.L.; Phatnani, H.P.; Mishina, Y.V.; Huper, G.; Selim, A.L.; Madison, E.R.; et al. Yeast screens identify the RNA polymerase II CTD and SPT5 as relevant targets of BRCA1 interaction. PLoS ONE 2008, 3, e1448. [Google Scholar] [CrossRef] [PubMed]
- Sherman, F. Getting started with yeast. Methods Enzym. 2002, 350, 3–41. [Google Scholar] [CrossRef]
- Schiestl, R.H.; Gietz, R.D.; Mehta, R.D.; Hastings, P.J. Carcinogens induce intrachromosomal recombination in yeast. Carcinogenesis 1989, 10, 1445–1455. [Google Scholar] [CrossRef] [Green Version]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Coyne, R.S.; McDonald, H.B.; Edgemon, K.; Brody, L.C. Functional characterization of BRCA1 sequence variants using a yeast small colony phenotype assay. Cancer Biol. 2004, 3, 453–457. [Google Scholar] [CrossRef] [Green Version]
- Fluss, R.; Faraggi, D.; Reiser, B. Estimation of the Youden Index and its Associated Cutoff Point. Biom. J. 2005, 47, 458–472. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Sensitivity | Specificity | Accuracy | AUROC | YI | Best Cut-off | FP | FN |
|---|---|---|---|---|---|---|---|---|
| Intra-HR | 0.957 | 0.783 | 0.87 | 0.914 | 0.739 | 1.535 | Y179C I1275V R1347G S1512I P1776H | R71K |
| Inter-HR | 0.87 | 0.913 | 0.891 | 0.929 | 0.783 | 1.32 | Y179C I1275V | R71K R1699W A1708E |
| GR | 0.913 | 0.87 | 0.891 | 0.867 | 0.783 | 1.077 | N132K I1275V T1675I | R71K R1495M |
| SCP | 1.00 | 0.913 | 0.957 | 0.951 | 0.913 | 25.833 | I1275V S1512I |
| yBRCA1 Method | |
|---|---|
| Accuracy (CI 95%) | 0.9565 (0.8516–0.9947) |
| Sensitivity | 0.9565 |
| Specificity | 0.9565 |
| AUROC | 0.9855 |
| Cohen’s Kappa | 0.9575 |
| MCC | 0.9583 |
| Inter-HR | Intra-HR | GR | SCP | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variant | PS | FI | PS | FI | PS | FI | PS | FI | PV | PPS | yBRCA1 |
| A1669S | 0.63575 | Pathogenic | 0.00675 | Benign | 0.00075 | Benign | 0.00025 | Benign | 1 | 0.25 | Benign |
| A1789T | 0.00025 | Benign | 0.00025 | Benign | 0.00025 | Benign | 0.99975 | Pathogenic | 1 | 0.25 | Benign |
| E1352K | 0.00025 | Benign | 0.56025 | Pathogenic | 0.99925 | Pathogenic | 0.99975 | Pathogenic | 3 | 0.75 | Pathogenic |
| N1730I | 0.00025 | Benign | 0.43825 | Pathogenic | 0.99925 | Pathogenic | 0.88025 | Pathogenic | 3 | 0.75 | Pathogenic |
| N1819S | 0.00125 | Benign | 0.01725 | Benign | 0.99975 | Pathogenic | 0.93575 | Pathogenic | 2 | 0.5 | Benign |
| P1010S | 0.40675 | Pathogenic | 0.00025 | Benign | 0.54025 | Pathogenic | 0.00025 | Benign | 2 | 0.5 | Benign |
| S1164I | 0.99975 | Pathogenic | 0.90575 | Pathogenic | 0.75425 | Pathogenic | 0.00025 | Benign | 3 | 0.75 | Pathogenic |
| S592N | 0.00025 | Benign | 0.89925 | Pathogenic | 0.99975 | Pathogenic | 0.99975 | Pathogenic | 3 | 0.75 | Pathogenic |
| V1791L | 0.99975 | Pathogenic | 0.30975 | Pathogenic | 0.00275 | Benign | 0.99975 | Pathogenic | 3 | 0.75 | Pathogenic |
| Y1703C | 0.99975 | Pathogenic | 0.32425 | Pathogenic | 0.54275 | Pathogenic | 0.99975 | Pathogenic | 4 | 1 | Pathogenic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellè, F.; Mercatanti, A.; Lodovichi, S.; Congregati, C.; Guglielmi, C.; Tancredi, M.; Caligo, M.A.; Cervelli, T.; Galli, A. Validation and Data-Integration of Yeast-Based Assays for Functional Classification of BRCA1 Missense Variants. Int. J. Mol. Sci. 2022, 23, 4049. https://doi.org/10.3390/ijms23074049
Bellè F, Mercatanti A, Lodovichi S, Congregati C, Guglielmi C, Tancredi M, Caligo MA, Cervelli T, Galli A. Validation and Data-Integration of Yeast-Based Assays for Functional Classification of BRCA1 Missense Variants. International Journal of Molecular Sciences. 2022; 23(7):4049. https://doi.org/10.3390/ijms23074049
Chicago/Turabian StyleBellè, Francesca, Alberto Mercatanti, Samuele Lodovichi, Caterina Congregati, Chiara Guglielmi, Mariella Tancredi, Maria Adelaide Caligo, Tiziana Cervelli, and Alvaro Galli. 2022. "Validation and Data-Integration of Yeast-Based Assays for Functional Classification of BRCA1 Missense Variants" International Journal of Molecular Sciences 23, no. 7: 4049. https://doi.org/10.3390/ijms23074049