
Small Prokaryotic DNA-Binding Proteins Protect Genome Integrity throughout the Life Cycle


Abstract
:1. Introduction
2. Bacterial Nucleoid-Associated Proteins and Genome Protection
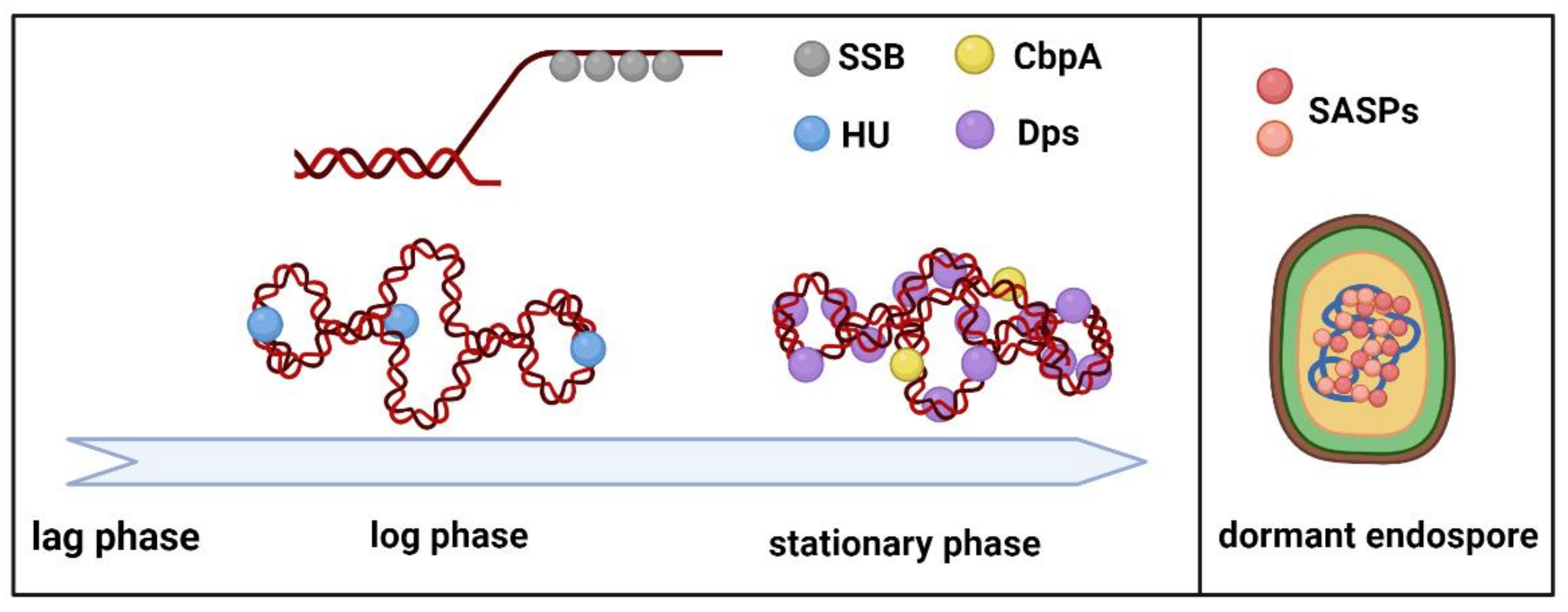
2.1. HU
2.1.1. HU Post-Translational Modifications
2.1.2. HU and Biofilms
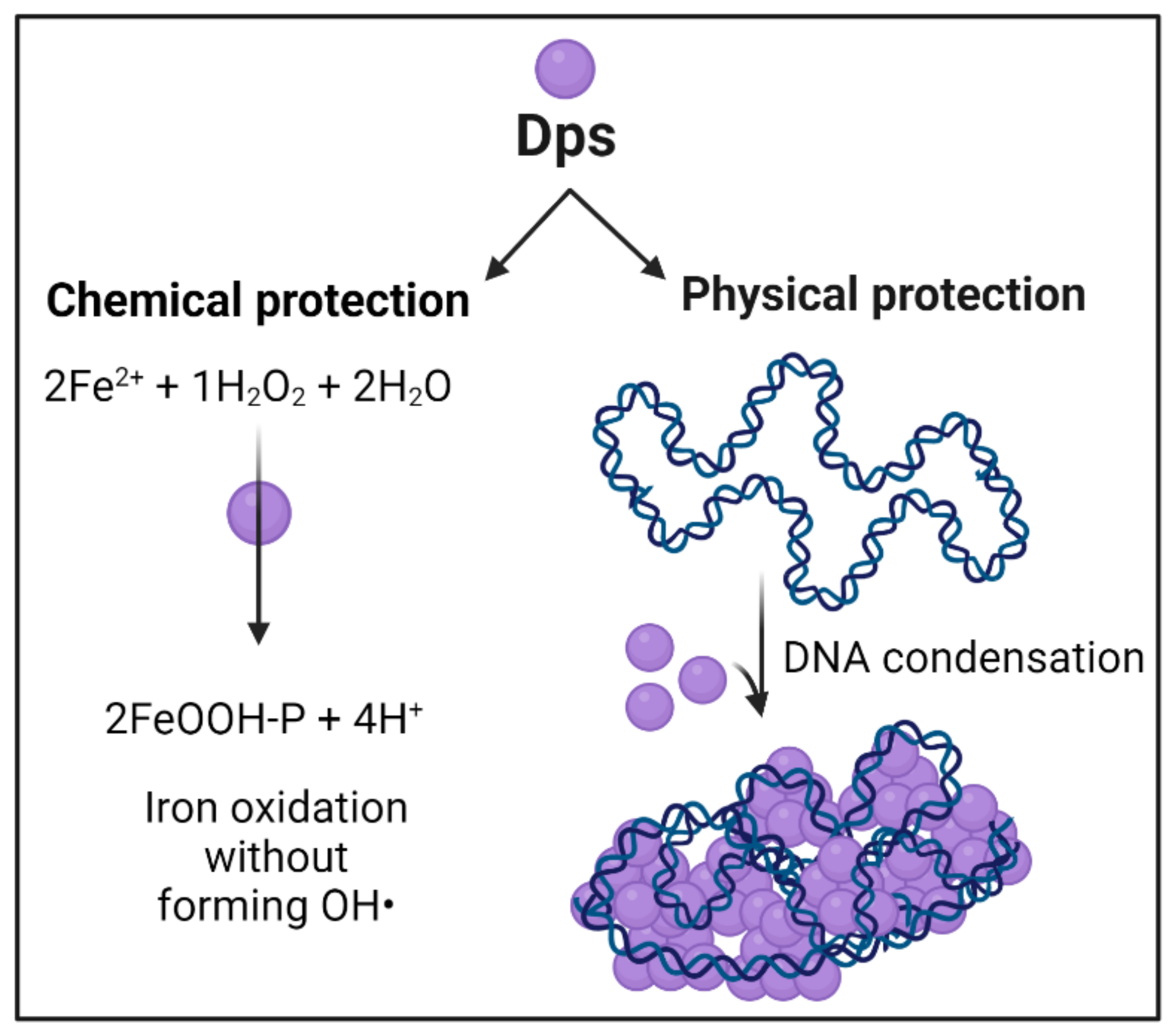
2.2. Dps
2.3. CbpA
3. Archaeal NAPs
4. Small Acid Soluble DNA Binding Proteins
5. Bacterial Single-Stranded DNA Binding Proteins
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foster, P.L. Stress-induced mutagenesis in bacteria. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 373–397. [Google Scholar] [CrossRef] [PubMed]
- Pennington, J.M.; Rosenberg, S.M. Spontaneous DNA breakage in single living Escherichia coli cells. Nat. Genet. 2007, 39, 797–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGlynn, P.; Savery, N.J.; Dillingham, M.S. The conflict between DNA replication and transcription. Mol. Microbiol. 2012, 85, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Butala, M.; Žgur-Bertok, D.; Busby, S.J.W. The bacterial LexA transcriptional repressor. Cell. Mol. Life Sci. 2009, 66, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Maslowska, K.H.; Makiela-Dzbenska, K.; Fijalkowska, I.J. The SOS system: A complex and tightly regulated response to DNA damage. Environ. Mol. Mutagen. 2019, 60, 368–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moeller, R.; Setlow, P.; Reitz, G.; Nicholson, W.L. Roles of small, acid-soluble spore proteins and core water content in survival of Bacillus subtilis spores exposed to environmental solar UV radiation. Appl. Environ. Microbiol. 2009, 75, 5202–5208. [Google Scholar] [CrossRef] [Green Version]
- Marceau, A.H. Functions of single-strand DNA-binding proteins in DNA replication, recombination, and repair. In Single-Stranded DNA Binding Proteins; Methods in Molecular Biology (Methods and Protocols); Keck, J., Ed.; Humana Press: Totowa, NJ, USA, 2012; Volume 922. [Google Scholar]
- Dillon, S.; Dorman, C. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef]
- Verma, S.C.; Qian, Z.; Adhya, S.L. Architecture of the Escherichia coli nucleoid. PLoS Genet. 2019, 15, e1008456. [Google Scholar] [CrossRef] [Green Version]
- Hołówka, J.; Zakrzewska-Czerwińska, J. Nucleoid associated proteins: The small organizers that help to cope with stress. Front. Microbiol. 2020, 11, 590. [Google Scholar] [CrossRef]
- Azam, T.A.; Ishihama, A. Twelve species of the nucleoid-associated protein from Escherichia coli. Sequence recognition specificity and DNA binding affinity. J. Biol. Chem. 1999, 274, 33105–33113. [Google Scholar] [CrossRef] [Green Version]
- Azam, T.A.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojkova, P.; Spidlova, P.; Stulik, J. Nucleoid-associated protein HU: A Lilliputian in gene regulation of bacterial virulence. Front. Cell. Infect. Microbiol. 2019, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Rouvière-Yaniv, J.; Gros, F.; Haselkorn, R.; Reiss, C. Histone-like Proteins in Prokaryotic Organisms and Their Interaction with DNA; Academic Press: New York, NY, USA, 1977; pp. 211–231. [Google Scholar]
- Boubrik, F.; Rouviere-Yaniv, J. Increased sensitivity to gamma irradiation in bacteria lacking protein HU. Proc. Natl. Acad. Sci. USA 1995, 92, 3958–3962. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Waters, R. Escherichia coli strains lacking protein HU are UV sensitive due to a role for HU in homologous recombination. J. Bacteriol. 1998, 180, 3750–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamashev, D.; Rouviere-Yaniv, J. The histone-like protein HU binds specifically to DNA recombination and repair intermediates. EMBO J. 2000, 19, 6527–6535. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Lo, L.F.; Maier, R.J. A histone-like protein of Helicobacter pylori protects DNA from stress damage and aids host colonization. DNA Repair 2012, 11, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preobrajenskaya, O.; Boullard, A.; Boubrik, F.; Schnarr, M.; Rouvière-Yaniv, J. The protein HU can displace the LexA repressor from its DNA-binding sites. Mol. Microbiol. 1994, 13, 459–467. [Google Scholar] [CrossRef]
- Pettijohn, D.E. Histone-like proteins and bacterial chromosome structure. J. Biol. Chem. 1988, 263, 12793–12796. [Google Scholar] [CrossRef]
- Hammel, M.; Amlanjyoti, D.; Reyes, F.E.; Chen, J.-H.; Parpana, R.; Tang, H.Y.H.; Larabell, C.A.; Tainer, J.A.; Adhya, S. HU multimerization shift controls nucleoid compaction. Sci. Adv. 2016, 2, e1600650. [Google Scholar] [CrossRef] [Green Version]
- Claret, L.; Rouviere-Yaniv, J. Variation in HU composition during growth of Escherichia coli: The heterodimer is required for long term survival. J. Mol. Biol. 1997, 273, 93–104. [Google Scholar] [CrossRef]
- Bhowmick, T.; Ghosh, S.; Dixit, K.; Ganesan, V.; Ramagopal, U.A.; Dey, D.; Sarma, S.P.; Ramakumar, S.; Nagaraja, V. Targeting Mycobacterium tuberculosis nucleoid-associated protein HU with structure-based inhibitors. Nat. Commun. 2014, 5, 4124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiva, A.M.O.; Friggen, A.H.; Qin, L.; Douwes, R.; Dame, R.T.; Smits, W.K. The bacterial chromatin protein HupA can remodel DNA and associates with the nucleoid in Clostridium difficile. J. Mol. Biol. 2019, 431, 653–672. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T. The role of nucleoid-associated proteins in the organization and compaction of bacterial chromatin. Mol. Microbiol. 2005, 56, 858–870. [Google Scholar] [CrossRef]
- Dame, R.T.; Kalmykowa, O.J.; Grainger, D.C. Chromosomal macrodomains and associated proteins: Implications for DNA organization and replication in gram negative bacteria. PLoS Genet. 2011, 7, e1002123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dame, R.T.; Rashid, F.-Z.M.; Grainger, D.C. Chromosome organization in bacteria: Mechanistic insights into genome structure and function. Nat. Rev. Genet. 2019, 21, 227–242. [Google Scholar] [CrossRef]
- Vis, H.; Mariani, M.; Vorgias, C.E.; Wilson, K.S.; Kaptein, R.; Boelens, R. Solution structure of the HU protein from Bacillus stearothermophilus. J. Mol. Biol. 1995, 254, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Swinger, K.K.; Lemberg, K.M.; Zhang, Y.; Rice, P.A. Flexible DNA bending in HU-DNA cocrystal structures. EMBO J. 2003, 22, 3749–3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Noort, J.; Verbrugge, S.; Goosen, N.; Dekker, C.; Dame, R.T. Dual architectural roles of HU: Formation of flexible hinges and rigid filaments. Proc. Natl. Acad. Sci. USA 2004, 101, 6969–6974. [Google Scholar] [CrossRef] [Green Version]
- Bettridge, K.; Verma, S.; Weng, X.; Adhya, S.; Xiao, J. Single-molecule tracking reveals that the nucleoid-associated protein HU plays a dual role in maintaining proper nucleoid volume through differential interactions with chromosomal DNA. Mol. Microbiol. 2021, 115, 12–27. [Google Scholar] [CrossRef]
- Makarova, K.S.; Aravind, L.; Wolf, Y.I.; Tatusov, R.L.; Minton, K.W.; Koonin, E.V.; Daly, M.J. Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics. Microbiol. Mol. Biol. Rev. 2001, 65, 44–79. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.H.; de la Tour, C.B.; Toueille, M.; Vannier, F.; Sommer, S.; Servant, P. The essential histone-like protein HU plays a major role in Deinococcus radiodurans nucleoid compaction. Mol. Microbiol. 2009, 73, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Floc’H, K.; Lacroix, F.; Servant, P.; Wong, Y.-S.; Kleman, J.-P.; Bourgeois, D.; Timmins, J. Cell morphology and nucleoid dynamics in dividing Deinococcus radiodurans. Nat. Commun. 2019, 10, 3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.; Sajid, A.; Sharma, K.; Ghosh, S.; Arora, G.; Singh, R.; Nagaraja, V.; Tandon, V.; Singh, Y. HupB, a nucleoid-associated protein of Mycobacterium tuberculosis, is modified by serine/threonine protein kinases in vivo. J. Bacteriol. 2014, 196, 2646–2657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Padmanabhan, B.; Anand, C.; Nagaraja, V. Lysine acetylation of the Mycobacterium tuberculosis HU protein modulates its DNA binding and genome organization. Mol. Microbiol. 2016, 100, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Anand, C.; Garg, R.; Ghosh, S.; Nagaraja, V. A Sir2 family protein Rv1151c deacetylates HU to alter its DNA binding mode in Mycobacterium tuberculosis. Biochem. Biophys. Res. Commun. 2017, 493, 1204–1209. [Google Scholar] [CrossRef]
- Sakatos, A.; Babunovic, G.H.; Chase, M.R.; Dills, A.; Leszyk, J.; Rosebrock, T.; Bryson, B.; Fortune, S.M. Posttranslational modification of a histone-like protein regulates phenotypic resistance to isoniazid in mycobacteria. Sci. Adv. 2018, 4, eaao1478. [Google Scholar] [CrossRef] [Green Version]
- Weinert, B.T.; Schölz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Thakur, B.; Arora, K.; Gupta, A.; Guptasarma, P. The DNA-binding protein HU is a molecular glue that attaches bacteria to extracellular DNA in biofilms. J. Biol. Chem. 2021, 296, 100532. [Google Scholar] [CrossRef]
- Kolter, R.; Siegele, D.A.; Tormo, A. The stationary phase of the bacterial life cycle. Annu. Rev. Microbiol. 1993, 47, 855–874. [Google Scholar] [CrossRef]
- Almiron, M.; Link, A.J.; Furlong, D.; Kolter, R. A novel DNA-binding protein with regulatory and protective roles in starved Escherichia coli. Genes Dev. 1992, 6, 2646–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talukder, A.A.; Ishihama, A. Dps is a stationary phase-specific protein of Escherichia coli nucleoid. Adv. Microbiol. 2014, 4, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Calhoun, L.N.; Kwon, Y.M. Structure, function and regulation of the DNA-binding protein Dps and its role in acid and oxidative stress resistance in Escherichia coli: A review. J. Appl. Microbiol. 2011, 110, 375–386. [Google Scholar] [CrossRef] [PubMed]
- De la Garza-García, J.A.; Ouahrani-Bettache, S.; Lyonnais, S.; Ornelas-Eusebio, E.; Freddi, L.; Al Dahouk, S.; Occhialini, A.; Köhler, S. Comparative genome-wide transcriptome analysis of Brucella suis and Brucella microti under acid stress at pH 4.5: Cold shock protein CspA and Dps are associated with acid resistance of B. microti. Front. Microbiol. 2021, 12, 794535. [Google Scholar] [CrossRef]
- Algu, K.; Choi, V.S.C.; Dhami, R.S.; Duncan, D.A.K. Dps confers protection of DNA sequence integrity in UV irradiated Escherichia coli. J. Exp. Microbiol. Immunol. 2007, 11, 60–65. [Google Scholar]
- Karas, V.O.; Westerlaken, I.; Meyer, A.S. The DNA-binding protein from starved cells (Dps) utilizes dual functions to defend cells against multiple stresses. J. Bacteriol. 2015, 197, 3206–3215. [Google Scholar] [CrossRef] [Green Version]
- Grant, R.A.; Filman, D.; Finkel, S.E.; Kolter, R.; Hogle, J.M. The crystal structure of Dps, a ferritin homolog that binds and protects DNA. Nat. Struct. Biol. 1998, 5, 294–303. [Google Scholar] [CrossRef]
- Wolf, S.; Frenkiel, D.; Arad, T.; Finkel, S.E.; Kolter, R.; Minsky, A. DNA protection by stress-induced biocrystallization. Nature 1999, 400, 83–85. [Google Scholar] [CrossRef]
- Antipov, S.; Turishchev, S.; Purtov, Y.; Shvyreva, U.; Sinelnikov, A.; Semov, Y.; Preobrazhenskaya, E.; Berezhnoy, A.; Shusharina, N.; Novolokina, N.; et al. The oligomeric form of the Escherichia coli Dps protein depends on the availability of iron ions. Molecules 2017, 22, 1904. [Google Scholar] [CrossRef] [Green Version]
- Ceci, P.; Cellai, S.; Falvo, E.; Rivetti, C.; Rossi, G.L.; Chiancone, E. DNA condensation and self-aggregation of Escherichia coli Dps are coupled phenomena related to the properties of the N-terminus. Nucleic Acids Res. 2004, 32, 5935–5944. [Google Scholar] [CrossRef] [Green Version]
- Chiancone, E.; Ceci, P. The multifaceted capacity of Dps proteins to combat bacterial stress conditions: Detoxification of iron and hydrogen peroxide and DNA binding. Biochim. Biophys. Acta 2010, 1800, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Melekhov, V.V.; Shvyreva, U.S.; Timchenko, A.A.; Tutukina, M.; Preobrazhenskaya, E.V.; Burkova, D.V.; Artiukhov, V.G.; Ozoline, O.; Antipov, S. Modes of Escherichia coli Dps interaction with DNA as revealed by atomic force microscopy. PLoS ONE 2015, 10, e0126504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrovin, E.V.; Dadinova, L.A.; Petoukhov, M.V.; Soshinskaya, E.Y.; Mozhaev, A.A.; Klinov, D.V.; Schäffer, T.E.; Shtykova, E.V.; Batishchev, O.V. Spatial organization of Dps and DNA–Dps complexes. J. Mol. Biol. 2021, 433, 166930. [Google Scholar] [CrossRef] [PubMed]
- Bessonova, T.A.; Shumeiko, S.A.; Purtov, Y.A.; Antipov, S.S.; Preobrazhenskaya, E.V.; Tutukina, M.N.; Ozoline, O.N. Hexuronates affect oligomeric form of a structural protein of bacterial nucleoid Dps and its ability to bind linear DNA fragments. Biophysics 2016, 61, 1059–1067. [Google Scholar] [CrossRef]
- Dadinova, L.; Kamyshinsky, R.; Chesnokov, Y.; Mozhaev, A.; Matveev, V.; Gruzinov, A.; Vasiliev, A.; Shtykova, E. Structural rearrangement of Dps-DNA complex caused by divalent Mg and Fe cations. Int. J. Mol. Sci. 2021, 22, 6056. [Google Scholar] [CrossRef]
- Papinutto, E.; Dundon, W.G.; Pitulis, N.; Battistutta, R.; Montecucco, C.; Zanotti, G. Structure of two iron-binding proteins from Bacillus anthracis. J. Biol. Chem. 2002, 277, 15093–15098. [Google Scholar] [CrossRef] [Green Version]
- Ceci, P.; Mangiarotti, L.; Rivetti, C.; Chiancone, E. The neutrophil-activating Dps protein of Helicobacter pylori, HP-NAP, adopts a mechanism different from Escherichia coli Dps to bind and condense DNA. Nucleic Acids Res. 2007, 35, 2247–2256. [Google Scholar] [CrossRef]
- Roy, S.; Saraswathi, R.; Chatterji, D.; Vijayan, M. Structural studies on the second Mycobacterium smegmatis Dps: Invariant and variable features of structure, assembly and function. J. Mol. Biol. 2008, 375, 948–959. [Google Scholar] [CrossRef]
- Chowdhury, R.P.; Saraswathi, R.; Chatterji, D. Mycobacterial stress regulation: The Dps “twin sister” defense mechanism and structure-function relationship. IUBMB Life 2010, 62, 67–77. [Google Scholar] [CrossRef]
- Ghatak, P.; Karmakar, K.; Kasetty, S.; Chatterji, D. Unveiling the role of Dps in the organization of mycobacterial nucleoid. PLoS ONE 2011, 6, e16019. [Google Scholar] [CrossRef]
- Santos, S.P.; Mitchell, E.P.; Franquelim, H.G.; Castanho, M.A.; Abreu, I.A.; Romão, C.V. Dps from Deinococcus radiodurans: Oligomeric forms of Dps1 with distinct cellular functions and Dps2 involved in metal storage. FEBS J. 2015, 282, 4307–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, C.; Ho, F.; Nenninger, A.; Raleiras, P.; Stensjö, K. Differential biochemical properties of three canonical Dps proteins from the cyanobacterium Nostoc punctiforme suggest distinct cellular functions. J. Biol. Chem. 2018, 293, 16635–16646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antipov, S.S.; Tutukina, M.N.; Preobrazhenskaya, E.V.; Kondrashov, F.A.; Patrushev, M.V.; Toshchakov, S.V.; Dominova, I.; Shvyreva, U.S.; Vrublevskaya, V.V.; Morenkov, O.S.; et al. The nucleoid protein Dps binds genomic DNA of Escherichia coli in a non-random manner. PLoS ONE 2017, 12, e0182800. [Google Scholar] [CrossRef] [PubMed]
- Janissen, R.; Arens, M.M.A.; Vtyurina, N.N.; Rivai, Z.; Sunday, N.D.; Eslami-Mossallam, B.; Gritsenko, A.A.; Laan, L.; de Ridder, D.; Artsimovitch, I.; et al. Global DNA compaction in stationary-phase bacteria does not affect transcription. Cell 2018, 174, 1188–1199.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrea, D.M.; Kriwacki, R.W. Phase separation in biology; functional organization of a higher order. Cell Commun. Signal. 2016, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Loiko, N.; Danilova, Y.; Moiseenko, A.; Kovalenko, V.; Tereshkina, K.; Tutukina, M.; El-Registan, G.; Sokolova, O.; Krupyanskii, Y. Morphological peculiarities of the DNA-protein complexes in starved Escherichia coli cells. PLoS ONE 2020, 15, e0231562. [Google Scholar] [CrossRef]
- Minsky, A.; Shimoni, E.; Frenkiel-Krispin, D. Stress, order and survival. Nat. Rev. Mol. Cell Biol. 2002, 3, 50–60. [Google Scholar] [CrossRef]
- Moiseenko, A.; Loiko, N.; Tereshkina, K.; Danilova, Y.; Kovalenko, V.; Chertkov, O.; Feofanov, A.V.; Krupyanskii, Y.F.; Sokolova, O.S. Projection structures reveal the position of the DNA within DNA-Dps Co-crystals. Biochem. Biophys. Res. Commun. 2019, 517, 463–469. [Google Scholar] [CrossRef]
- Theoret, J.R.; Cooper, K.K.; Zekarias, B.; Roland, K.L.; Law, B.F.; Curtiss, R.; Joens, L.A. The Campylobacter jejuni Dps homologue is important for in vitro biofilm formation and cecal colonization of poultry and may serve as a protective antigen for vaccination. Clin. Vaccine Immunol. 2012, 19, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- Lacqua, A.; Wanner, O.; Colangelo, T.; Martinotti, M.G.; Landini, P. Emergence of biofilm-forming subpopulations upon exposure of Escherichia coli to environmental bacteriophages. Appl. Environ. Microbiol. 2006, 72, 956–959. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Muramatsu, S.; Mizuno, T. An Escherichia coli protein that preferentially binds to sharply curved DNA. J. Biochem. 1990, 108, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.S.; Grainger, D.C. The Escherichia coli nucleoid in stationary phase. Adv. Appl. Microbiol. 2013, 83, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Bird, J.G.; Sharma, S.; Roshwalb, S.C.; Hoskins, J.R.; Wickner, S. Functional analysis of CbpA, a DnaJ homolog and nucleoid-associated DNA-binding protein. J. Biol. Chem. 2006, 281, 34349–34356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgriff, S.; Chintakayala, K.; Chim, Y.T.A.; Chen, X.; Allen, S.; Lovering, A.L.; Grainger, D.C. Dimerization and DNA-dependent aggregation of the Escherichia coli nucleoid protein and chaperone CbpA. Mol. Microbiol. 2010, 77, 1289–1300. [Google Scholar] [CrossRef] [Green Version]
- Chintakayala, K.; Sellars, L.E.; Singh, S.S.; Shahapure, R.; Westerlaken, I.; Meyer, A.S.; Dame, R.T.; Grainger, D.C. DNA recognition by Escherichia coli CbpA protein requires a conserved arginine-minor-groove interaction. Nucleic Acids Res. 2015, 43, 2282–2292. [Google Scholar] [CrossRef] [Green Version]
- Baker, B.J.; De Anda, V.; Seitz, K.W.; Dombrowski, N.; Santoro, A.E.; Lloyd, K.G. Diversity, ecology and evolution of Archaea. Nat. Microbiol. 2020, 5, 887–900. [Google Scholar] [CrossRef]
- Adam, P.S.; Borrel, G.; Brochier-Armanet, C.; Gribaldo, S. The growing tree of Archaea: New perspectives on their diversity, evolution and ecology. ISME J. 2017, 11, 2407–2425. [Google Scholar] [CrossRef]
- Peeters, E.; Driessen, R.P.; Werner, F.; Dame, R.T. The interplay between nucleoid organization and transcription in archaeal genomes. Nat. Rev. Microbiol. 2015, 13, 333–341. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, M.; Chen, Y.; Wang, L.; Liu, Q.; Dong, Y.; Gong, Y.; Huang, L. Architectural roles of Cren7 in folding crenarchaeal chromatin filament. Mol. Microbiol. 2019, 111, 556–569. [Google Scholar] [CrossRef]
- Laursen, S.P.; Bowerman, S.; Luger, K. Archaea: The final frontier of chromatin. J. Mol. Biol. 2021, 433, 166791. [Google Scholar] [CrossRef]
- Loth, K.; Largillière, J.; Coste, F.; Culard, F.; Landon, C.; Castaing, B.; Delmas, A.F.; Paquet, F. New protein-DNA complexes in archaea: A small monomeric protein induces a sharp V-turn DNA structure. Sci. Rep. 2019, 9, 14253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocher, A.; Rojec, M.; Swadling, J.B.; Esin, A.; Warnecke, T. The DNA-binding protein HTa from Thermoplasma acidophilum is an archaeal histone analog. eLife 2019, 8, e52542. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Xie, T.; Hu, Q.; Deng, C.; Zheng, R.; Chen, W. Genome-wide comparison of ferritin family from Archaea, Bacteria, Eukarya, and Viruses: Its distribution, characteristic motif, and phylogenetic relationship. Naturwissenschaften 2015, 102, 64. [Google Scholar] [CrossRef]
- Jones, D.L.; Baxter, B.K. DNA repair and photoprotection: Mechanisms of overcoming environmental ultraviolet radiation exposure in halophilic archaea. Front. Microbiol. 2017, 8, 1882. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.J.; Santangelo, T.J. Archaeal DNA repair mechanisms. Biomolecules 2020, 10, 1472. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Spore resistance properties. Microbiol. Spectr. 2014, 2, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Li, L. Spore photoproduct lyase: The known, the controversial, and the unknown. J. Biol. Chem. 2015, 290, 4003–4009. [Google Scholar] [CrossRef] [Green Version]
- Djouiai, B.; Thwaite, J.E.; Laws, T.R.; Commichau, F.M.; Setlow, B.; Setlow, P.; Moeller, R. Role of DNA repair and protective components in Bacillus subtilis spore resistance to inactivation by 400-nm-wavelength blue light. Appl. Environ. Microbiol. 2018, 84, e01604. [Google Scholar] [CrossRef] [Green Version]
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile infection. Nat. Rev. Dis. Primers 2016, 2, 16020. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Bioterrorism Agents/Diseases A to Z by Category: Category A; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2012. Available online: https://emergency.cdc.gov/agent/agentlist-category.asp (accessed on 26 March 2022).
- Talukdar, P.K.; Olguín-Araneda, V.; Alnoman, M.; Paredes-Sabja, D.; Sarker, M.R. Updates on the sporulation process in Clostridium species. Res. Microbiol. 2015, 166, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Hoch, J.A. Regulation of the phosphorelay and the initiation of sporulation in Bacillus subtilis. Annu. Rev. Microbiol. 1993, 47, 441–465. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Bumbaca, D.; Kosman, J.; Setlow, P.; Jedrzejas, M.J. Structure of a protein–DNA complex essential for DNA protection in spores of Bacillus species. Proc. Natl. Acad. Sci. USA 2008, 105, 2806–2811. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Mechanisms for the prevention of damage to DNA in spores of Bacillus species. Annu. Rev. Microbiol. 1995, 49, 29–54. [Google Scholar] [CrossRef]
- Tennen, R.; Setlow, B.; Davis, K.L.; Loshon, C.A.; Setlow, P. Mechanisms of killing of spores of Bacillus subtilis by iodine, glutaraldehyde and nitrous acid. J. Appl. Microbiol. 2000, 89, 330–338. [Google Scholar] [CrossRef]
- Meaney, C.A.; Cartman, S.T.; McClure, P.J.; Minton, N.P. The role of small acid-soluble proteins (SASPs) in protection of spores of Clostridium botulinum against nitrous acid. Int. J. Food Microbiol. 2016, 216, 25–30. [Google Scholar] [CrossRef]
- Raju, D.; Waters, M.; Setlow, P.; Sarker, M.R. Investigating the role of small, acid-soluble spore proteins (SASPs) in the resistance of Clostridium perfringens spores to heat. BMC Microbiol. 2006, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Raju, D.; Setlow, P.; Sarker, M.R. Antisense-RNA-mediated decreased synthesis of small, acid-soluble spore proteins leads to decreased resistance of Clostridium perfringens spores to moist heat and UV radiation. Appl. Environ. Microbiol. 2007, 73, 2048–2053. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; McClane, B.A. A novel small acid soluble protein variant is important for spore resistance of most Clostridium perfringens food poisoning isolates. PLoS Pathog. 2008, 4, e1000056. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Paredes-Sabja, D.; Sarker, M.R.; McClane, B.A. Further characterization of Clostridium perfringens small acid soluble protein-4 (Ssp4) properties and expression. PLoS ONE 2009, 4, e6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes-Sabja, D.; Raju, D.; Torres, J.A.; Sarker, M.R. Role of small, acid-soluble spore proteins in the resistance of Clostridium perfringens spores to chemicals. Int. J. Food Microbiol. 2008, 122, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Nerber, H.N.; Sorg, J.A. The small acid-soluble proteins of Clostridioides difficile are important for UV resistance and serve as a check point for sporulation. PLoS Pathog. 2021, 17, e1009516. [Google Scholar] [CrossRef] [PubMed]
- Greipel, J.; Urbanke, C.; Maass, G. The single-stranded DNA binding protein of Escherichia coli: Physicochemical properties and biological functions. In Protein-Nucleic Acid Interaction; Saenger, W., Heinemann, U., Eds.; Macmillan: London, UK, 1989; pp. 61–86. [Google Scholar] [CrossRef]
- Shereda, R.D.; Kozlov, A.G.; Lohman, T.M.; Cox, M.M.; Keck, J.L. SSB as an organizer/mobilizer of genome maintenance complexes. Crit. Rev. Biochem. Mol. Biol. 2009, 43, 289–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.T.; Ciesielski, G.L. The essential, ubiquitous single-stranded DNA-binding proteins. In Single Stranded DNA Binding Proteins; Methods in Molecular Biology; Oliveira, M.T., Ed.; Humana: New York, NY, USA, 2021; Volume 2281, pp. 1–21. [Google Scholar] [CrossRef]
- Raghunathan, S.; Ricard, C.S.; Lohman, T.M.; Waksman, G. Crystal structure of the homo-tetrameric DNA binding domain of Escherichia coli single-stranded DNA-binding protein determined by multiwavelength x-ray diffraction on the selenomethionyl protein at 2.9-Å resolution. Proc. Natl. Acad. Sci. USA 1997, 94, 6652–6657. [Google Scholar] [CrossRef] [Green Version]
- Filipkowski, P.; Koziatek, M.; Kur, J. A highly thermostable, homodimeric single-stranded DNA-binding protein from Deinococcus radiopugnans. Extremophiles 2006, 10, 607–614. [Google Scholar] [CrossRef]
- Bernstein, D.A.; Eggington, J.M.; Killoran, M.P.; Misic, A.M.; Cox, M.M.; Keck, J.L. Crystal structure of the Deinococcus radiodurans single-stranded DNA-binding protein suggests a mechanism for coping with DNA damage. Proc. Natl. Acad. Sci. USA 2004, 101, 8575–8580. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, J.S.; DeVeaux, L.C. The essential role of the Deinococcus radiodurans ssb gene in cell survival and radiation tolerance. PLoS ONE 2013, 8, e71651. [Google Scholar] [CrossRef]
- George, N.P.; Ngo, K.V.; Chitteni-Pattu, S.; Norais, C.A.; Battista, J.R.; Cox, M.M.; Keck, J.L. Structure and cellular dynamics of Deinococcus radiodurans single-stranded DNA (ssDNA)-binding protein (SSB)-DNA complexes. J. Biol. Chem. 2012, 287, 22123–22132. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, R.; Inoue, J.; Mikawa, T.; Ha, T. Single molecule analysis of Thermus thermophilus SSB protein dynamics on single-stranded DNA. Nucleic Acids Res. 2014, 42, 3821–3832. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| NAPs | Structure | Protection | DNA Binding | Function | Identified in |
|---|---|---|---|---|---|
| HU | α2, β2, αβ | physical | chromosomal dsDNA, non-specific, weak; binding to AT-rich and specific DNA structures | nucleoid compaction, maintaining proper nucleoid volume | bacteria |
| Dps | α12 | physical, chemical | chromosome/DNA, some sequence or structural selectivity | iron oxidation in ferroxidase centre; DNA condensation, co-crystallization with DNA | in ˃ 1000 species of bacteria and archaea |
| CbpA | monomer or dimer | physical | AT-rich DNA, non-specific | forms DNA aggregates, co-chaperone | γ-proteobacteria, highly conserved |
| NAPs | Structure | DNA Binding | Function | Identified in |
|---|---|---|---|---|
| Alba | dimer | DNA/RNA binding | genome packaging and organization | all archea except Methanomicrobia, Haloarchaea |
| MC1 | monomer | non-specific binding; prefers bent DNA | compacts DNA induces a V-turn on DNA; protects against thermal denaturation and radiolysis | Methanomicrobia, Haloarchaea |
| HTa (HU homolog) | histone-like oligomerization | GC-rich DNA | wraps DNA, required for DNA folding | Thermoplasmatales |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molan, K.; Žgur Bertok, D. Small Prokaryotic DNA-Binding Proteins Protect Genome Integrity throughout the Life Cycle. Int. J. Mol. Sci. 2022, 23, 4008. https://doi.org/10.3390/ijms23074008
Molan K, Žgur Bertok D. Small Prokaryotic DNA-Binding Proteins Protect Genome Integrity throughout the Life Cycle. International Journal of Molecular Sciences. 2022; 23(7):4008. https://doi.org/10.3390/ijms23074008
Chicago/Turabian StyleMolan, Katja, and Darja Žgur Bertok. 2022. "Small Prokaryotic DNA-Binding Proteins Protect Genome Integrity throughout the Life Cycle" International Journal of Molecular Sciences 23, no. 7: 4008. https://doi.org/10.3390/ijms23074008