
The Methyltransferase HemK Regulates the Virulence and Nutrient Utilization of the Phytopathogenic Bacterium Xanthomonas citri Subsp. citri

 ,
,
Abstract
:1. Introduction
2. Results
2.1. HemK Influences the Cell Motility, Biofilm Formation, Extracellular Polysaccharide, and Enzyme Production in Xcc jx-6
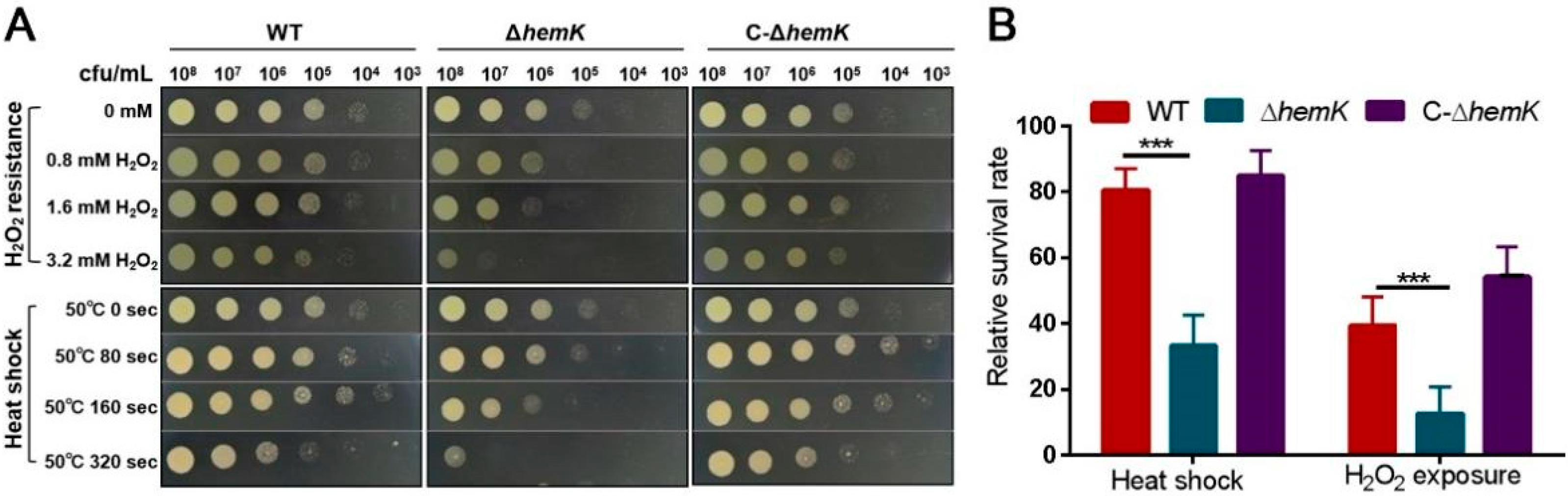
2.2. HemK Contributes to Bacterial Stress Tolerance of Oxidative Stress and Heat Shock in Xcc jx-6
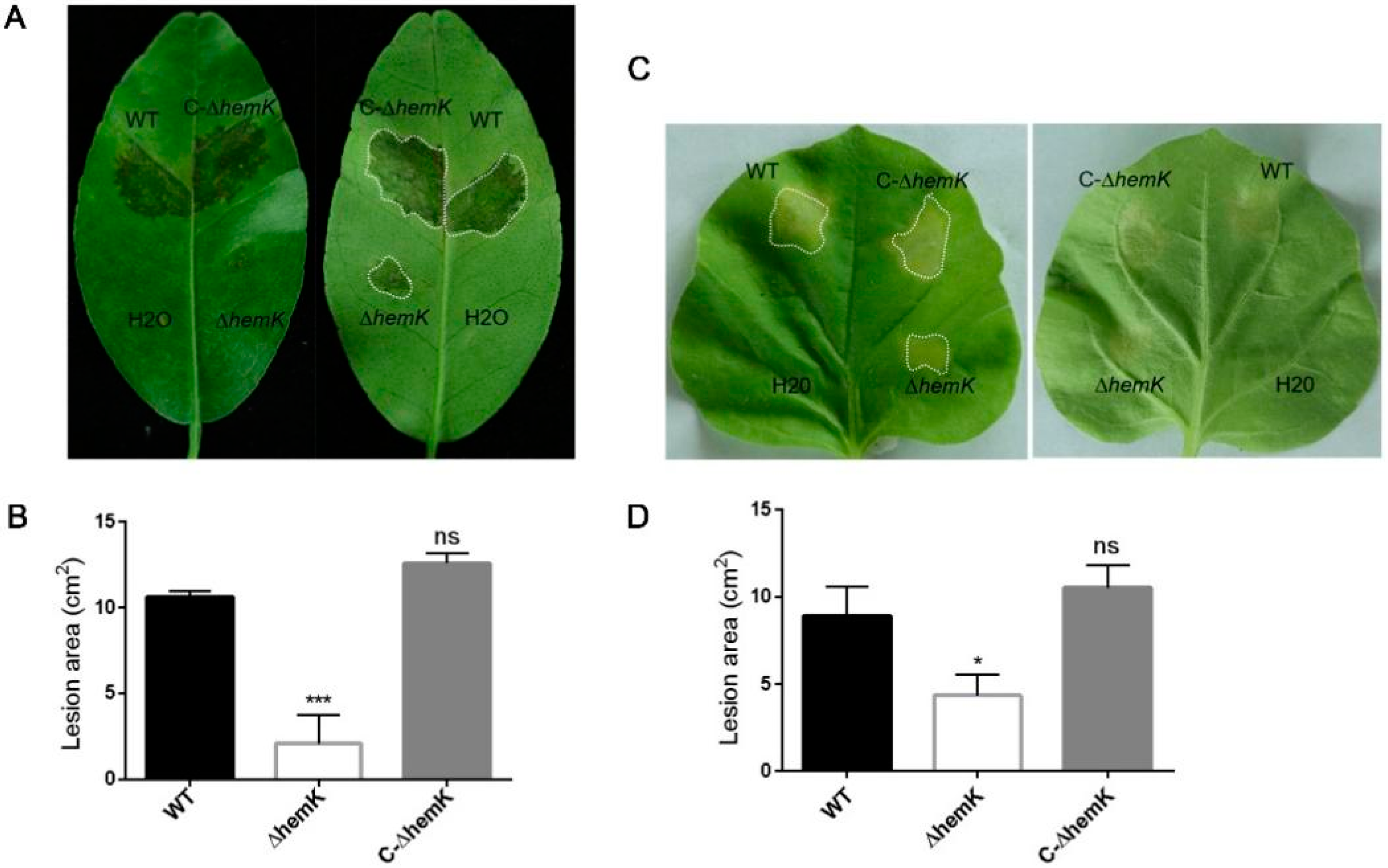
2.3. Mutation of hemK Impaired Activation of Virulence on Citrus and Hypersensitive Response to Nicotiana benthamiana
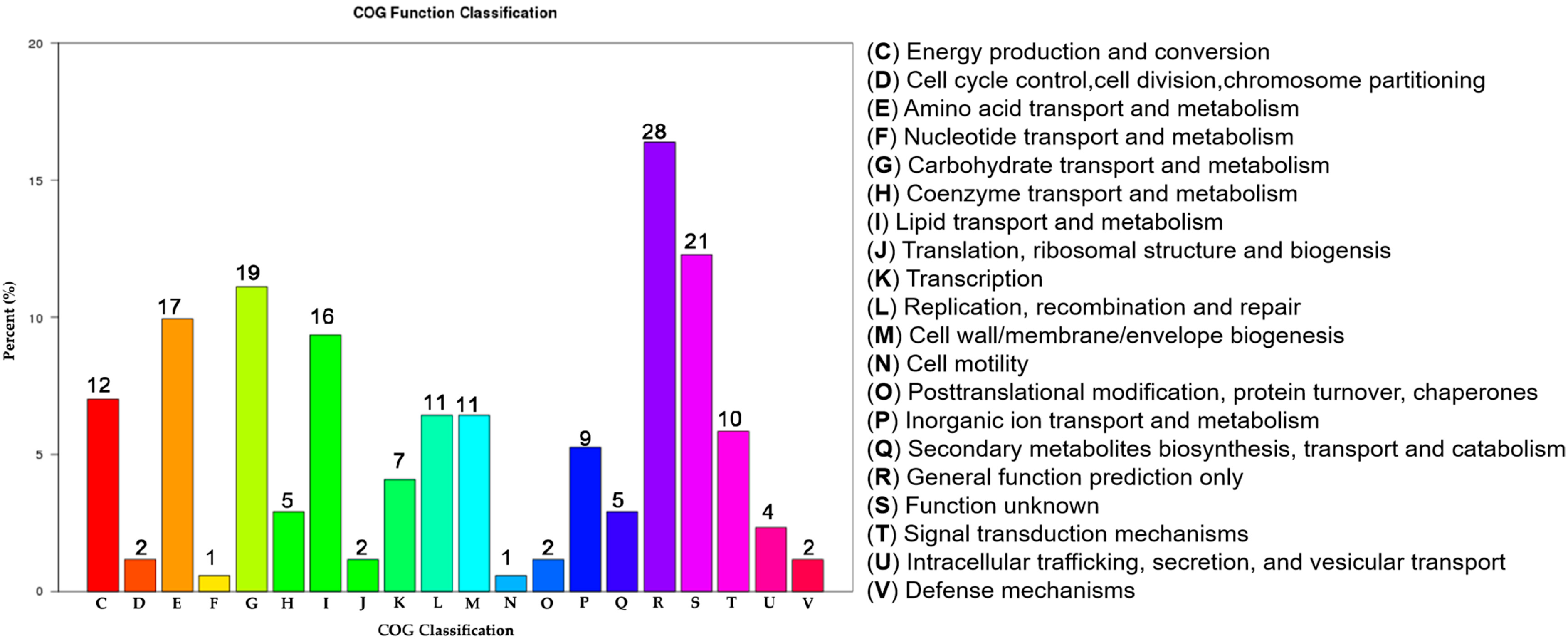
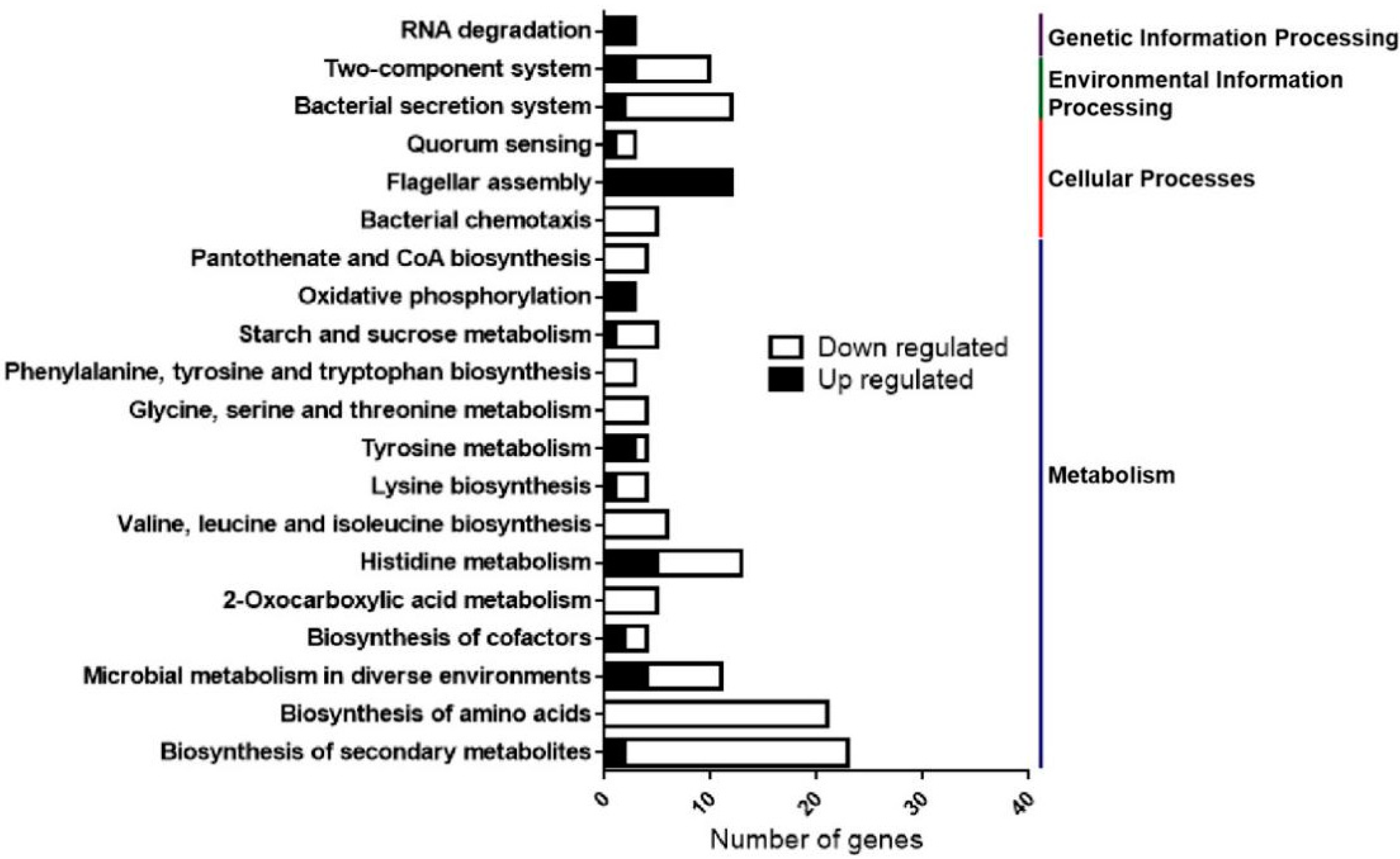
2.4. Transcriptome RNA Sequencing (RNA-Seq) Analysis Reveals Multiple Physiological Processes Regulated by HemK in Xcc
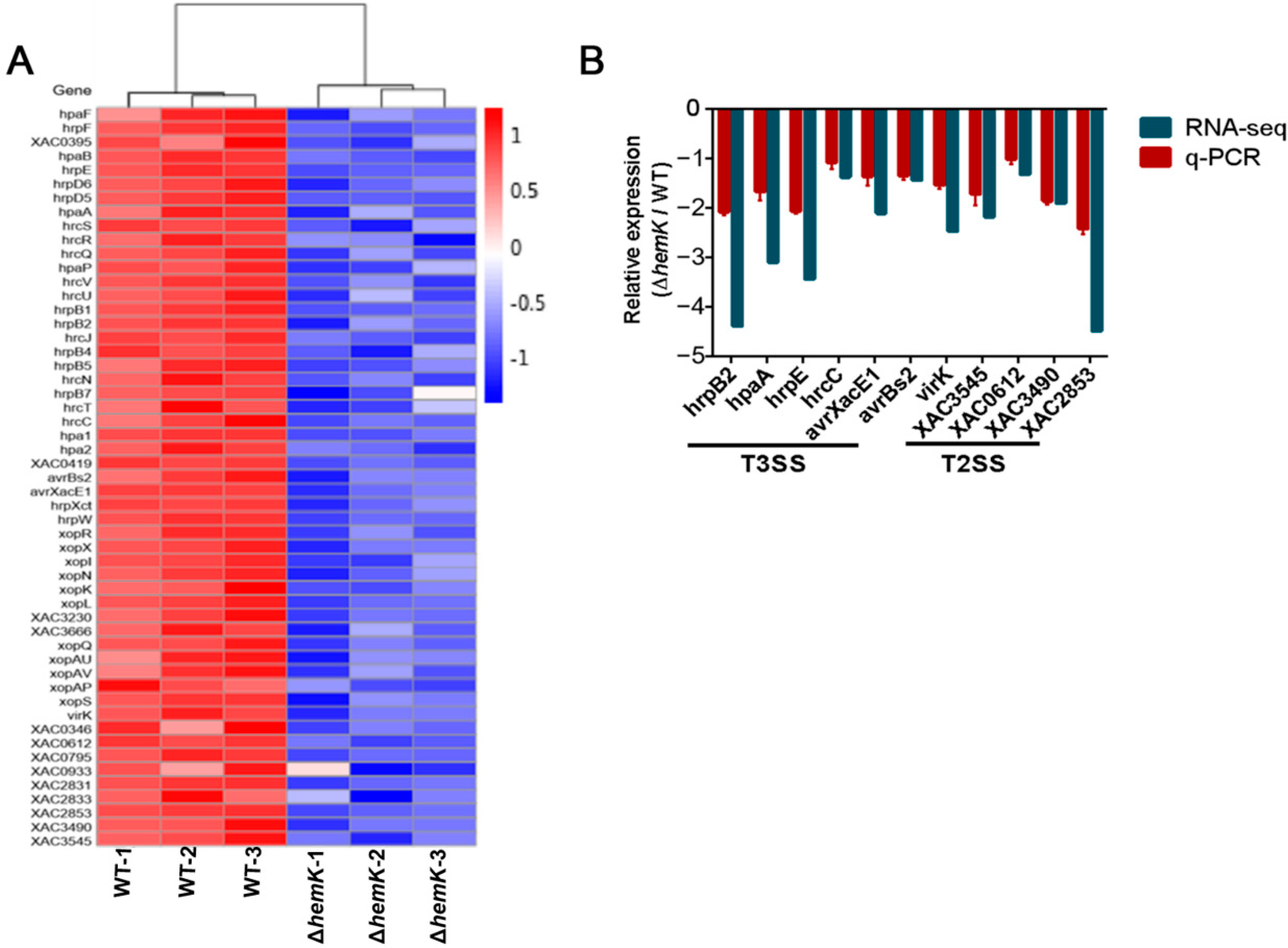
2.5. HemK Controls the Expression of T3SS and Its Associated Effectors, as Well as Many Extracellular Enzymes Secreted by T2SS
2.6. HemK Is Implicated in the Regulation of the Expression of Genes Involved in Diffusible Signal Factor (DSF) Mediated Quorum Sensing of Xcc
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Culture Media, and Culture Conditions
4.2. Plasmid Construction and Primers Used
4.3. Construction of the ∆hemK Mutant and Its Complementary Strain in Xcc jx-6
4.4. Bacterial Growth Curve
4.5. Bacterial Motility Assays
4.6. Bacterial Biofilm Formation Assay
4.7. Extracellular Enzymes Activity Assay
4.8. Extracellular Polysaccharides Production Assays
4.9. Stress Tolerance Assay
4.10. Virulence and Hypersensitive Response Assays
4.11. High-Throughput RNA Sequencing (RNA-Seq) and Data Analysis
4.12. Quantitative Real-Time PCR (qRT-PCR) Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Brunings, A.M.; Gabriel, D.W. Xanthomonas citri: Breaking the surface. Mol. Plant Pathol. 2003, 4, 141–157. [Google Scholar] [CrossRef]
- Vojnov, A.A.; Do, A.A.; Dow, J.M.; Castagnaro, A.P.; Marano, M.R. Bacteria causing important diseases of citrus utilise distinct modes of pathogenesis to attack a common host. Appl. Microbiol. Biotechnol. 2010, 87, 467–477. [Google Scholar] [CrossRef]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant-pathogen interactions. Nat. Rev. Microbiol 2020, 18, 415–427. [Google Scholar] [CrossRef]
- An, S.Q.; Potnis, N.; Dow, M.; Vorholter, F.J.; He, Y.Q.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Rev. 2020, 44, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Ference, C.M.; Gochez, A.M.; Behlau, F.; Wang, N.; Graham, J.H.; Jones, J.B. Recent advances in the understanding of Xanthomonas citri ssp. citri pathogenesis and citrus canker disease management. Mol. Plant Pathol. 2018, 19, 1302–1318. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.J.; Wu, T.L.; Zheng, P.X.; Ou, J.Y.; Ni, H.F.; Lin, Y.C. Comparative Genomic Analysis Uncovered Evolution of Pathogenicity Factors, Horizontal Gene Transfer Events, and Heavy Metal Resistance Traits in Citrus Canker Bacterium Xanthomonas citri subsp. citri. Front. Microbiol. 2021, 12, 731711. [Google Scholar] [CrossRef]
- Da, S.A.; Ferro, J.A.; Reinach, F.C.; Farah, C.S.; Furlan, L.R.; Quaggio, R.B.; Monteiro-Vitorello, C.B.; Van Sluys, M.A.; Almeida, N.F.; Alves, L.M.; et al. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 2002, 417, 459–463. [Google Scholar] [CrossRef]
- Zhang, Y.; Teper, D.; Xu, J.; Wang, N. Stringent response regulators (p)ppGpp and DksA positively regulate virulence and host adaptation of Xanthomonas citri. Mol. Plant Pathol. 2019, 20, 1550–1565. [Google Scholar] [CrossRef] [Green Version]
- Braun, S.G.; Meyer, A.; Holst, O.; Puhler, A.; Niehaus, K. Characterization of the Xanthomonas campestris pv. campestris lipopolysaccharide substructures essential for elicitation of an oxidative burst in tobacco cells. Mol. Plant-Microbe Interact. 2005, 18, 674–681. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, N. The wxacO gene of Xanthomonas citri ssp. citri encodes a protein with a role in lipopolysaccharide biosynthesis, biofilm formation, stress tolerance and virulence. Mol. Plant Pathol. 2011, 12, 381–396. [Google Scholar] [CrossRef]
- Rigano, L.A.; Siciliano, F.; Enrique, R.; Sendin, L.; Filippone, P.; Torres, P.S.; Questa, J.; Dow, J.M.; Castagnaro, A.P.; Vojnov, A.A.; et al. Biofilm formation, epiphytic fitness, and canker development in Xanthomonas axonopodis pv. citri. Mol. Plant-Microbe Interact. 2007, 20, 1222–1230. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Guo, J.; Ma, W.X.; Ji, Z.Y.; Zou, L.F.; Chen, G.Y.; Zou, H.S. Identification of seven novel virulence genes from Xanthomonas citri subsp. citri by Tn5-based random mutagenesis. J. Microbiol. 2015, 53, 330–336. [Google Scholar] [CrossRef]
- Xia, T.; Li, Y.; Sun, D.; Zhuo, T.; Fan, X.; Zou, H. Identification of an Extracellular Endoglucanase That Is Required for Full Virulence in Xanthomonas citri subsp. citri. PLoS ONE 2016, 11, e151017. [Google Scholar] [CrossRef] [Green Version]
- Buttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Figueiredo, F.; Jones, J.; Wang, N. HrpG and HrpX play global roles in coordinating different virulence traits of Xanthomonas axonopodis pv. citri. Mol. Plant-Microbe Interact. 2011, 24, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Hu, X.; Li, J.; Wang, N. A Novel Periplasmic Protein, VrpA, Contributes to Efficient Protein Secretion by the Type III Secretion System in Xanthomonas spp. Mol. Plant-Microbe Interact. 2015, 28, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Hu, X.; Wang, N. The novel virulence-related gene nlxA in the lipopolysaccharide cluster of Xanthomonas citri ssp. citri is involved in the production of lipopolysaccharide and extracellular polysaccharide, motility, biofilm formation and stress resistance. Mol. Plant Pathol. 2012, 13, 923–934. [Google Scholar] [CrossRef]
- Baptista, J.C.; Machado, M.A.; Homem, R.A.; Torres, P.S.; Vojnov, A.A.; Do, A.A. Mutation in the xpsD gene of Xanthomonas axonopodis pv. citri affects cellulose degradation and virulence. Genet. Mol. Biol. 2010, 33, 146–153. [Google Scholar] [CrossRef]
- Li, R.F.; Lu, G.T.; Li, L.; Su, H.Z.; Feng, G.F.; Chen, Y.; He, Y.Q.; Jiang, B.L.; Tang, D.J.; Tang, J.L. Identification of a putative cognate sensor kinase for the two-component response regulator HrpG, a key regulator controlling the expression of the hrp genes in Xanthomonas campestris pv. campestris. Environ. Microbiol. 2014, 16, 2053–2071. [Google Scholar] [CrossRef]
- Zhou, X.; Yan, Q.; Wang, N. Deciphering the regulon of a GntR family regulator via transcriptome and ChIP-exo analyses and its contribution to virulence in Xanthomonas citri. Mol. Plant Pathol. 2017, 18, 249–262. [Google Scholar] [CrossRef]
- Nakahigashi, K.; Kubo, N.; Narita, S.; Shimaoka, T.; Goto, S.; Oshima, T.; Mori, H.; Maeda, M.; Wada, C.; Inokuchi, H. HemK, a class of protein methyl transferase with similarity to DNA methyl transferases, methylates polypeptide chain release factors, and hemK knockout induces defects in translational termination. Proc. Natl. Acad. Sci. USA 2002, 99, 1473–1478. [Google Scholar] [CrossRef] [Green Version]
- Polevoda, B.; Span, L.; Sherman, F. The yeast translation release factors Mrf1p and Sup45p (eRF1) are methylated, respectively, by the methyltransferases Mtq1p and Mtq2p. J. Biol. Chem. 2006, 281, 2562–2571. [Google Scholar] [CrossRef] [Green Version]
- Heurgue-Hamard, V.; Champ, S.; Mora, L.; Merkulova-Rainon, T.; Kisselev, L.L.; Buckingham, R.H. The glutamine residue of the conserved GGQ motif in Saccharomyces cerevisiae release factor eRF1 is methylated by the product of the YDR140w gene. J. Biol. Chem. 2005, 280, 2439–2445. [Google Scholar] [CrossRef] [Green Version]
- Figaro, S.; Scrima, N.; Buckingham, R.H.; Heurgue-Hamard, V. HemK2 protein, encoded on human chromosome 21, methylates translation termination factor eRF1. FEBS Lett. 2008, 582, 2352–2356. [Google Scholar] [CrossRef] [Green Version]
- Kailasam, S.; Singh, S.; Liu, M.J.; Lin, C.C.; Yeh, K.C. A HemK class glutamine-methyltransferase is involved in the termination of translation and essential for iron homeostasis in Arabidopsis. New Phytol. 2020, 226, 1361–1374. [Google Scholar] [CrossRef]
- Zhou, X.; Cooke, P.; Li, L. Eukaryotic release factor 1-2 affects Arabidopsis responses to glucose and phytohormones during germination and early seedling development. J. Exp. Bot. 2010, 61, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Meurer, J.; Lezhneva, L.; Amann, K.; Godel, M.; Bezhani, S.; Sherameti, I.; Oelmuller, R. A peptide chain release factor 2 affects the stability of UGA-containing transcripts in Arabidopsis chloroplasts. Plant. Cell 2002, 14, 3255–3269. [Google Scholar] [CrossRef] [Green Version]
- Pustelny, C.; Brouwer, S.; Musken, M.; Bielecka, A.; Dotsch, A.; Nimtz, M.; Haussler, S. The peptide chain release factor methyltransferase PrmC is essential for pathogenicity and environmental adaptation of Pseudomonas aeruginosa PA14. Environ. Microbiol. 2013, 15, 597–609. [Google Scholar] [CrossRef]
- Garbom, S.; Olofsson, M.; Bjornfot, A.C.; Srivastava, M.K.; Robinson, V.L.; Oyston, P.; Titball, R.W.; Wolf-Watz, H. Phenotypic characterization of a virulence-associated protein, VagH, of Yersinia pseudotuberculosis reveals a tight link between VagH and the type III secretion system. Microbiology 2007, 153, 1464–1473. [Google Scholar] [CrossRef] [Green Version]
- Heurgue-Hamard, V.; Champ, S.; Engstrom, A.; Ehrenberg, M.; Buckingham, R.H. The hemK gene in Escherichia coli encodes the N(5)-glutamine methyltransferase that modifies peptide release factors. EMBO J. 2002, 21, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.E.; O’Toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybak, M.; Minsavage, G.V.; Stall, R.E.; Jones, J.B. Identification of Xanthomonas citri ssp. citri host specificity genes in a heterologous expression host. Mol. Plant Pathol. 2009, 10, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Teper, D.; Pandey, S.S.; Wang, N. The HrpG/HrpX Regulon of Xanthomonads-An Insight to the Complexity of Regulation of Virulence Traits in Phytopathogenic Bacteria. Microorganisms 2021, 9, 187. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Zhang, Y.; Wang, N. Diffusible signal factor (DSF)-mediated quorum sensing modulates expression of diverse traits in Xanthomonas citri and responses of citrus plants to promote disease. BMC Genom. 2019, 20, 55. [Google Scholar] [CrossRef]
- Samal, B.; Chatterjee, S. New insight into bacterial social communication in natural host: Evidence for interplay of heterogeneous and unison quorum response. PLoS Genet. 2019, 15, e1008395. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Nie, S.; Li, B.; Yang, Z.Q.; Xu, Z.M.; Fei, J.; Lin, C.; Zeng, R.; Xu, G.L. Deficiency in a glutamine-specific methyltransferase for release factor causes mouse embryonic lethality. Mol. Cell. Biol. 2010, 30, 4245–4253. [Google Scholar] [CrossRef] [Green Version]
- Matilla, M.A.; Krell, T. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 2018, 42, fux052. [Google Scholar] [CrossRef] [Green Version]
- Antunez-Lamas, M.; Cabrera-Ordonez, E.; Lopez-Solanilla, E.; Raposo, R.; Trelles-Salazar, O.; Rodriguez-Moreno, A.; Rodriguez-Palenzuela, P. Role of motility and chemotaxis in the pathogenesis of Dickeya dadantii 3937 (ex Erwinia chrysanthemi 3937). Microbiology 2009, 155, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Sgro, G.G.; Ficarra, F.A.; Dunger, G.; Scarpeci, T.E.; Valle, E.M.; Cortadi, A.; Orellano, E.G.; Gottig, N.; Ottado, J. Contribution of a harpin protein from Xanthomonas axonopodis pv. citri to pathogen virulence. Mol. Plant Pathol. 2012, 13, 1047–1059. [Google Scholar] [CrossRef]
- Galan, J.E.; Collmer, A. Type III secretion machines: Bacterial devices for protein delivery into host cells. Science 1999, 284, 1322–1328. [Google Scholar] [CrossRef] [Green Version]
- Dunger, G.; Garofalo, C.G.; Gottig, N.; Garavaglia, B.S.; Rosa, M.C.; Farah, C.S.; Orellano, E.G.; Ottado, J. Analysis of three Xanthomonas axonopodis pv. citri effector proteins in pathogenicity and their interactions with host plant proteins. Mol. Plant Pathol. 2012, 13, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Wengelnik, K.; Van den Ackerveken, G.; Bonas, U. HrpG, a key hrp regulatory protein of Xanthomonas campestris pv. vesicatoria is homologous to two-component response regulators. Mol. Plant-Microbe Interact. 1996, 9, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Li, R.F.; Wang, X.X.; Wu, L.; Huang, L.; Qin, Q.J.; Yao, J.L.; Lu, G.T.; Tang, J.L. Xanthomonas campestris sensor kinase HpaS co-opts the orphan response regulator VemR to form a branched two-component system that regulates motility. Mol. Plant Pathol. 2020, 21, 360–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Amaral, A.M.D.; Toledo, C.P.; Baptista, J.C.; Machado, M.A. Transformation of Xanthomonas axonopodis pv. citri by electroporation. Fitopatol. Bras. 2005, 30, 292–294. [Google Scholar] [CrossRef]
- Malamud, F.; Conforte, V.P.; Rigano, L.A.; Castagnaro, A.P.; Marano, M.R.; Morais, D.A.A.; Vojnov, A.A. HrpM is involved in glucan biosynthesis, biofilm formation and pathogenicity in Xanthomonas citri ssp. citri. Mol. Plant Pathol. 2012, 13, 1010–1018. [Google Scholar] [CrossRef]
- Medina, A.; Lambert, R.J.; Magan, N. Rapid throughput analysis of filamentous fungal growth using turbidimetric measurements with the Bioscreen C: A tool for screening antifungal compounds. Fungal Biol. 2012, 116, 161–169. [Google Scholar] [CrossRef]
- Li, R.F.; Cui, P.; Wei, P.Z.; Liu, X.Y.; Tang, J.L.; Lu, G.T. HprKXcc is a serine kinase that regulates virulence in the Gram-negative phytopathogen Xanthomonas campestris. Environ. Microbiol. 2019, 21, 4504–4520. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Wang, N. The ColR/ColS two-component system plays multiple roles in the pathogenicity of the citrus canker pathogen Xanthomonas citri subsp. citri. J. Bacteriol. 2011, 193, 1590–1599. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.L.; Liu, Y.N.; Barber, C.E.; Dow, J.M.; Wootton, J.C.; Daniels, M.J. Genetic and molecular analysis of a cluster of rpf genes involved in positive regulation of synthesis of extracellular enzymes and polysaccharide in Xanthomonas campestris pathovar campestris. Mol. Gen. Genet. 1991, 226, 409–417. [Google Scholar] [CrossRef]
- Slater, H.; Alvarez-Morales, A.; Barber, C.E.; Daniels, M.J.; Dow, J.M. A two-component system involving an HD-GYP domain protein links cell-cell signalling to pathogenicity gene expression in Xanthomonas campestris. Mol. Microbiol. 2000, 38, 986–1003. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Mao, W.; Han, Y.; Zhang, X.; Yang, C.; Chen, Y.; Chen, Y.; Xu, J.; Li, H.; Qi, X.; et al. Structural characteristics and antioxidant activities of the extracellular polysaccharides produced by marine bacterium Edwardsiella tarda. Bioresour. Technol. 2010, 101, 4729–4732. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, N. The gpsX gene encoding a glycosyltransferase is important for polysaccharide production and required for full virulence in Xanthomonas citri subsp. citri. BMC Microbiol. 2012, 12, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.; Ding, T.; Chang, C.; Yu, C.; Li, X.; Liu, Q. Global Regulator PhoP is Necessary for Motility, Biofilm Formation, Exoenzyme Production and Virulence of Xanthomonas citri subsp. citri on Citrus Plants. Genes 2019, 10, 340. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Tag | Gene Name | Log2Fold Change | Protein Function |
|---|---|---|---|
| XAC2992 | XAC2992 | −1.5 | endoproteinase Arg-C, degrading host defense proteins |
| XAC0612 | engXCA | −1.3 | cellulase |
| XAC3120 | glk | 1.3 | glucose kinase |
| XAC3921 | ugt | −1.2 | glucosyltransferase |
| XAC1556 | fucP | −1.3 | glucose-galactose transporter |
| XAC1557 | scrK | −1.2 | fructokinase |
| XAC3489 | fyuA | −2.3 | TonB-dependent receptor |
| XAC3490 | XAC3490 | −1.9 | amylosucrase or alpha amylase |
| XAC0465 | XAC0465 | −1.0 | metalloproteinase |
| XAC4327 | uahA | −1.8 | urea amidolyase |
| XAC1820 | metL | −3.3 | aspartokinase |
| XAC1821 | thrB | −3.4 | homoserine kinase |
| XAC1823 | thrC | −3.6 | threonine synthase |
| XAC1828 | hisG | −2.4 | ATP phosphoribosyltransferase |
| XAC1829 | hisD | −2.3 | histidinol dehydrogenase |
| XAC1830 | hisC | −2.4 | histidinol-phosphate aminotransferase |
| XAC1831 | hisB | −2.5 | Imidazole glycerol phosphate dehydratase/histidinol-phosphate phosphatase bifunctional enzyme |
| XAC1832 | hisH | −1.9 | amidotransferase |
| XAC1833 | hisA | −2.2 | phosphoribosylformimino-5-aminoimidazole carboxam |
| XAC1834 | hisF | −2.3 | cyclase |
| XAC1835 | hisI | −2.4 | phosphoribosyl-AMP cyclohydrolase/phosphoribosyl—ATP pyrophosphatase bifunctional enzyme |
| XAC3451 | ilvC | −2.7 | ketol-acid reductoisomerase |
| XAC3452 | ilvG | −2.2 | acetolactate synthase isozyme II large subunit |
| XAC3453 | ilvM | −2.5 | acetolactate synthase isozyme II large subunit |
| XAC3454 | tdcB | −2.1 | threonine dehydratase catabolic |
| XAC3455 | leuA | −1.3 | 2-isopropylmalate synthase |
| XAC0999 | cirA | −1.1 | colicin I receptor |
| XAC3546 | xadA | −1.7 | autotransporter adhesion protein |
| XAC1471 | XAC1471 | −1.1 | glycine zipper 2TM domain containing protein |
| XAC1827 | XAC1827 | −3.2 | Trp repressor protein |
| XAC3085 | XAC3085 | −1.1 | putative type III secretion system effector protein |
| XAC3754 | XAC3754 | 1.4 | putative chemotaxis membrane protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Yang, X.; Ye, X.; Feng, J.; Cheng, T.; Zhou, X.; Liu, D.X.; Xu, L.; Wang, J. The Methyltransferase HemK Regulates the Virulence and Nutrient Utilization of the Phytopathogenic Bacterium Xanthomonas citri Subsp. citri. Int. J. Mol. Sci. 2022, 23, 3931. https://doi.org/10.3390/ijms23073931
Shi Y, Yang X, Ye X, Feng J, Cheng T, Zhou X, Liu DX, Xu L, Wang J. The Methyltransferase HemK Regulates the Virulence and Nutrient Utilization of the Phytopathogenic Bacterium Xanthomonas citri Subsp. citri. International Journal of Molecular Sciences. 2022; 23(7):3931. https://doi.org/10.3390/ijms23073931
Chicago/Turabian StyleShi, Yu, Xiaobei Yang, Xiaoxin Ye, Jiaying Feng, Tianfang Cheng, Xiaofan Zhou, Ding Xiang Liu, Linghui Xu, and Junxia Wang. 2022. "The Methyltransferase HemK Regulates the Virulence and Nutrient Utilization of the Phytopathogenic Bacterium Xanthomonas citri Subsp. citri" International Journal of Molecular Sciences 23, no. 7: 3931. https://doi.org/10.3390/ijms23073931