
Role of Glucocorticoid Signaling and HDAC4 Activation in Diaphragm and Gastrocnemius Proteolytic Activity in Septic Rats

 , , , and
, , , and
Abstract
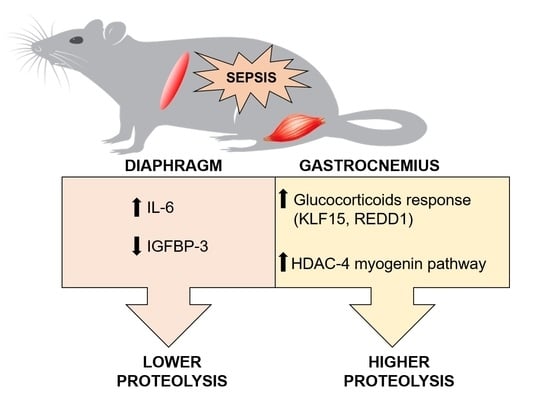
:
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Protocol
4.2. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.3. Protein Analysis by Western Blot
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lakshmikanth, C.L.; Jacob, S.P.; Chaithra, V.H.; de Castro-Faria-Neto, H.C.; Marathe, G.K. Sepsis: In search of cure. Inflamm. Res. 2016, 65, 587–602. [Google Scholar] [CrossRef]
- Dumitru, A.; Radu, B.M.; Radu, M.; Cretoiu, S.M. Muscle Changes During Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 73–92. [Google Scholar] [CrossRef] [PubMed]
- Klaude, M.; Mori, M.; Tjader, I.; Gustafsson, T.; Wernerman, J.; Rooyackers, O. Protein metabolism and gene expression in skeletal muscle of critically ill patients with sepsis. Clin. Sci. 2012, 122, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Morel, J.; Palao, J.C.; Castells, J.; Desgeorges, M.; Busso, T.; Molliex, S.; Jahnke, V.; Del Carmine, P.; Gondin, J.; Arnould, D.; et al. Regulation of Akt-mTOR, ubiquitin-proteasome and autophagy-lysosome pathways in locomotor and respiratory muscles during experimental sepsis in mice. Sci. Rep. 2017, 7, 10866. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, M.; Muthny, T.; Sispera, L.; Holecek, M. Effects of beta-hydroxy-beta-methylbutyrate treatment in different types of skeletal muscle of intact and septic rats. J. Physiol. Biochem. 2010, 66, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Stana, F.; Vujovic, M.; Mayaki, D.; Leduc-Gaudet, J.P.; Leblanc, P.; Huck, L.; Hussain, S.N.A. Differential Regulation of the Autophagy and Proteasome Pathways in Skeletal Muscles in Sepsis. Crit. Care Med. 2017, 45, e971–e979. [Google Scholar] [CrossRef] [PubMed]
- Moarbes, V.; Mayaki, D.; Huck, L.; Leblanc, P.; Vassilakopoulos, T.; Petrof, B.J.; Hussain, S.N.A. Differential regulation of myofibrillar proteins in skeletal muscles of septic mice. Physiol. Rep. 2019, 7, e14248. [Google Scholar] [CrossRef]
- Owen, A.M.; Patel, S.P.; Smith, J.D.; Balasuriya, B.K.; Mori, S.F.; Hawk, G.S.; Stromberg, A.J.; Kuriyama, N.; Kaneki, M.; Rabchevsky, A.G.; et al. Chronic muscle weakness and mitochondrial dysfunction in the absence of sustained atrophy in a preclinical sepsis model. Elife 2019, 8, e49920. [Google Scholar] [CrossRef] [PubMed]
- Yende, S.; Austin, S.; Rhodes, A.; Finfer, S.; Opal, S.; Thompson, T.; Bozza, F.A.; LaRosa, S.P.; Ranieri, V.M.; Angus, D.C. Long-Term Quality of Life Among Survivors of Severe Sepsis: Analyses of Two International Trials. Crit. Care Med. 2016, 44, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, C.E.; Bersten, A.D. Alterations in Respiratory and Limb Muscle Strength and Size in Patients With Sepsis Who Are Mechanically Ventilated. Phys. Ther. 2014, 94, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, E.; Paraskevas, T.; Velissaris, D. Sepsis and the muscle tissue. A narrative review. Rom. J. Intern. Med. 2021, 59, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.C.; Pai, M.H.; Wu, J.M.; Yang, P.J.; Lee, P.C.; Chen, K.Y.; Yeh, S.L.; Lin, M.T. Protective Effects of Glutamine and Leucine Supplementation on Sepsis-Induced Skeletal Muscle Injuries. Int. J. Mol. Sci. 2021, 22, 13003. [Google Scholar] [CrossRef] [PubMed]
- Jude, B.; Tissier, F.; Dubourg, A.; Droguet, M.; Castel, T.; Leon, K.; Giroux-Metges, M.A.; Pennec, J.P. TGF-beta Pathway Inhibition Protects the Diaphragm From Sepsis-Induced Wasting and Weakness in Rat. Shock 2020, 53, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Londhe, P.; Guttridge, D.C. Inflammation induced loss of skeletal muscle. Bone 2015, 80, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.I.; Priego, T.; Lopez-Calderon, A. Hormones and Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 207–233. [Google Scholar] [CrossRef]
- Soto, L.; Martin, A.I.; Millan, S.; Vara, E.; Lopez-Calderon, A. Effects of endotoxin lipopolysaccharide administration on the somatotropic axis. J. Endocrinol. 1998, 159, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.I.; Gomez-SanMiguel, A.B.; Priego, T.; Lopez-Calderon, A. Formoterol treatment prevents the effects of endotoxin on muscle TNF/NF-kB, Akt/mTOR, and proteolytic pathways in a rat model. Role of IGF-I and miRNA 29b. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E705–E714. [Google Scholar] [CrossRef]
- Martin, A.I.; Priego, T.; Moreno-Ruperez, A.; Gonzalez-Hedstrom, D.; Granado, M.; Lopez-Calderon, A. IGF-1 and IGFBP-3 in Inflammatory Cachexia. Int. J. Mol. Sci. 2021, 22, 9469. [Google Scholar] [CrossRef]
- Gomez-SanMiguel, A.B.; Villanua, M.A.; Martin, A.I.; Lopez-Calderon, A. D-TRP(8)-gammaMSH Prevents the Effects of Endotoxin in Rat Skeletal Muscle Cells through TNFalpha/NF-KB Signalling Pathway. PLoS ONE 2016, 11, e0155645. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Schakman, O.; Kalista, S.; Barbe, C.; Loumaye, A.; Thissen, J.P. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Minetto, M.A.; Lanfranco, F.; Motta, G.; Allasia, S.; Arvat, E.; D’Antona, G. Steroid myopathy: Some unresolved issues. J. Endocrinol. Investig. 2011, 34, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.E.; Van Remmen, H. Emerging roles for histone deacetylases in age-related muscle atrophy. Nutr. Healthy Aging 2016, 4, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Martin, S.C.; Parkington, J.; Cadena, S.M.; Zhu, J.; Ibebunjo, C.; Summermatter, S.; Londraville, N.; Patora-Komisarska, K.; Widler, L.; et al. HDAC4 Controls Muscle Homeostasis through Deacetylation of Myosin Heavy Chain, PGC-1alpha, and Hsc70. Cell Rep. 2019, 29, 749–763.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Hedstrom, D.; Priego, T.; Lopez-Calderon, A.; Amor, S.; de la Fuente-Fernandez, M.; Inarejos-Garcia, A.M.; Garcia-Villalon, A.L.; Martin, A.I.; Granado, M. Beneficial Effects of a Mixture of Algae and Extra Virgin Olive Oils on the Age-Induced Alterations of Rodent Skeletal Muscle: Role of HDAC-4. Nutrients 2020, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Mielcarek, M.; Zielonka, D.; Carnemolla, A.; Marcinkowski, J.T.; Guidez, F. HDAC4 as a potential therapeutic target in neurodegenerative diseases: A summary of recent achievements. Front. Cell. Neurosci. 2015, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bricceno, K.V.; Sampognaro, P.J.; Van Meerbeke, J.P.; Sumner, C.J.; Fischbeck, K.H.; Burnett, B.G. Histone deacetylase inhibition suppresses myogenin-dependent atrogene activation in spinal muscular atrophy mice. Hum. Mol. Genet 2012, 21, 4448–4459. [Google Scholar] [CrossRef] [Green Version]
- Hotchkiss, R.S.; Karl, I.E. Reevaluation of the role of cellular hypoxia and bioenergetic failure in sepsis. JAMA 1992, 267, 1503–1510. [Google Scholar] [CrossRef]
- Levy, B.; Gibot, S.; Franck, P.; Cravoisy, A.; Bollaert, P.E. Relation between muscle Na+K+ ATPase activity and raised lactate concentrations in septic shock: A prospective study. Lancet 2005, 365, 871–875. [Google Scholar] [CrossRef]
- Garcia-Alvarez, M.; Marik, P.; Bellomo, R. Sepsis-associated hyperlactatemia. Crit. Care 2014, 18, 503. [Google Scholar] [CrossRef] [Green Version]
- de Morais, H.; de Fatima Silva, F.; da Silva, F.G.; Silva, M.O.; Graciano, M.F.R.; Martins, M.I.L.; Carpinelli, Â.R.; Mazucco, T.L.; Bazotte, R.B.; de Souza, H.M. Insulin, not glutamine dipeptide, reduces lipases expression and prevents fat wasting and weight loss in Walker 256 tumor-bearing rats. Eur. J. Pharmacol. 2017, 806, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Mofarrahi, M.; Sigala, I.; Guo, Y.; Godin, R.; Davis, E.C.; Petrof, B.; Sandri, M.; Burelle, Y.; Hussain, S.N. Autophagy and skeletal muscles in sepsis. PLoS ONE 2012, 7, e47265. [Google Scholar] [CrossRef] [PubMed]
- Goodman, M.N. Interleukin-6 induces skeletal muscle protein breakdown in rats. Proc. Soc. Exp. Biol. Med. 1994, 205, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Sun, J.; Li, M.; Qian, L.; Zhang, L.; Huang, Z.; Shen, Y.; Law, B.Y.; Liu, L.; Gu, X. Transcriptome Analysis of Immune Receptor Activation and Energy Metabolism Reduction as the Underlying Mechanisms in Interleukin-6-Induced Skeletal Muscle Atrophy. Front. Immunol. 2021, 12, 730070. [Google Scholar] [CrossRef]
- Fischer, C.P. Interleukin-6 in acute exercise and training: What is the biological relevance? Exerc. Immunol. Rev. 2006, 12, 6–33. [Google Scholar]
- Bruells, C.S.; Duschner, P.; Marx, G.; Gayan-Ramirez, G.; Frank, N.; Breuer, T.; Krenkel, O.; Tacke, F.; Mossanen, J.C. Acute liver injury following acetaminophen administration does not activate atrophic pathways in the mouse diaphragm. Sci. Rep. 2021, 11, 6302. [Google Scholar] [CrossRef]
- Garcia-Martinez, C.; Lopez-Soriano, F.J.; Argiles, J.M. Interleukin-6 does not activate protein breakdown in rat skeletal muscle. Cancer Lett. 1994, 76, 1–4. [Google Scholar] [CrossRef]
- Gao, S.; Durstine, J.L.; Koh, H.J.; Carver, W.E.; Frizzell, N.; Carson, J.A. Acute myotube protein synthesis regulation by IL-6-related cytokines. Am. J. Physiol. Cell Physiol. 2017, 313, C487–C500. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Zhou, Y.; Bao, W.; Fu, Q.; Hao, H.; Han, L.; Zhang, X.; Tian, X.; Zhang, M. STAT3 and IL-6 Contribute to Corticosteroid Resistance in an OVA and Ozone-induced Asthma Model with Neutrophil Infiltration. Front. Mol. Biosci. 2021, 8, 717962. [Google Scholar] [CrossRef]
- Baetz, A.; Frey, M.; Heeg, K.; Dalpke, A.H. Suppressor of cytokine signaling (SOCS) proteins indirectly regulate toll-like receptor signaling in innate immune cells. J. Biol. Chem. 2004, 279, 54708–54715. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Liu, W.; Dai, Y. SOCS3 and its role in associated diseases. Hum. Immunol. 2015, 76, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.P.; Grossberg, A.J.; Krasnow, S.M.; Levasseur, P.R.; Szumowski, M.; Zhu, X.X.; Maxson, J.E.; Knoll, J.G.; Barnes, A.P.; Marks, D.L. Cancer- and endotoxin-induced cachexia require intact glucocorticoid signaling in skeletal muscle. FASEB J. 2013, 27, 3572–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surmachevska, N.; Tiwari, V. Corticosteroid Induced Myopathy. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Shimizu, N.; Yoshikawa, N.; Ito, N.; Maruyama, T.; Suzuki, Y.; Takeda, S.; Nakae, J.; Tagata, Y.; Nishitani, S.; Takehana, K.; et al. Crosstalk between glucocorticoid receptor and nutritional sensor mTOR in skeletal muscle. Cell Metab. 2011, 13, 170–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, S.; Wang, B.; Orihuela, Y.; Hong, E.G.; Fisch, S.; Haldar, S.; Cline, G.W.; Kim, J.K.; Peroni, O.D.; Kahn, B.B.; et al. Regulation of gluconeogenesis by Kruppel-like factor 15. Cell Metab. 2007, 5, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britto, F.A.; Dumas, K.; Giorgetti-Peraldi, S.; Ollendorff, V.; Favier, F.B. Is REDD1 a metabolic double agent? Lessons from physiology and pathology. Am. J. Physiol. Cell Physiol. 2020, 319, C807–C824. [Google Scholar] [CrossRef]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model. Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Britto, F.A.; Begue, G.; Rossano, B.; Docquier, A.; Vernus, B.; Sar, C.; Ferry, A.; Bonnieu, A.; Ollendorff, V.; Favier, F.B. REDD1 deletion prevents dexamethasone-induced skeletal muscle atrophy. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E983–E993. [Google Scholar] [CrossRef] [Green Version]
- Schulze, P.C.; Gielen, S.; Adams, V.; Linke, A.; Mobius-Winkler, S.; Erbs, S.; Kratzsch, J.; Hambrecht, R.; Schuler, G. Muscular levels of proinflammatory cytokines correlate with a reduced expression of insulinlike growth factor-I in chronic heart failure. Basic Res. Cardiol. 2003, 98, 267–274. [Google Scholar] [CrossRef]
- Costelli, P.; Muscaritoli, M.; Bossola, M.; Penna, F.; Reffo, P.; Bonetto, A.; Busquets, S.; Bonelli, G.; Lopez-Soriano, F.J.; Doglietto, G.B.; et al. IGF-1 is downregulated in experimental cancer cachexia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R674–R683. [Google Scholar] [CrossRef] [Green Version]
- Castillero, E.; Martin, A.I.; Lopez-Menduina, M.; Granado, M.; Villanua, M.A.; Lopez-Calderon, A. IGF-I system, atrogenes and myogenic regulatory factors in arthritis induced muscle wasting. Mol. Cell. Endocrinol. 2009, 309, 8–16. [Google Scholar] [CrossRef]
- Nystrom, G.; Pruznak, A.; Huber, D.; Frost, R.A.; Lang, C.H. Local insulin-like growth factor I prevents sepsis-induced muscle atrophy. Metabolism 2009, 58, 787–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Celemin, L.; Pasko, N.; Blomart, V.; Thissen, J.P. Inhibition of muscle insulin-like growth factor I expression by tumor necrosis factor-alpha. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E1279–E1290. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Huang, Z.L.; Yang, J.H.; Xu, Y.H.; Sun, J.S.; Zheng, Q.; Wei, C.Y.; Song, W.; Yuan, Z. Pancreatic cancer cell-derived IGFBP-3 contributes to muscle wasting. J. Exp. Clin. Cancer Res. 2016, 35, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakovenko, A.; Cameron, M.; Trevino, J.G. Molecular therapeutic strategies targeting pancreatic cancer induced cachexia. World J. Gastrointest. Surg. 2018, 10, 95–106. [Google Scholar] [CrossRef]
- Lopez-Menduina, M.; Martin, A.I.; Castillero, E.; Villanua, M.A.; Lopez-Calderon, A. Systemic IGF-I administration attenuates the inhibitory effect of chronic arthritis on gastrocnemius mass and decreases atrogin-1 and IGFBP-3. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R541–R551. [Google Scholar] [CrossRef] [Green Version]
- Cole, C.L.; Bachman, J.F.; Ye, J.; Murphy, J.; Gerber, S.A.; Beck, C.A.; Boyce, B.F.; Muthukrishnan, G.; Chakkalakal, J.V.; Schwarz, E.M.; et al. Increased myocellular lipid and IGFBP-3 expression in a pre-clinical model of pancreatic cancer-related skeletal muscle wasting. J. Cachexia Sarcopenia Muscle 2021, 12, 731–745. [Google Scholar] [CrossRef]
- Puig-Vilanova, E.; Aguilo, R.; Rodriguez-Fuster, A.; Martinez-Llorens, J.; Gea, J.; Barreiro, E. Epigenetic mechanisms in respiratory muscle dysfunction of patients with chronic obstructive pulmonary disease. PLoS ONE 2014, 9, e111514. [Google Scholar] [CrossRef]
- Ha, Z.L.; Yu, Z.Y. Downregulation of miR-29b-3p aggravates podocyte injury by targeting HDAC4 in LPS-induced acute kidney injury. Kaohsiung J. Med. Sci. 2021, 37, 1069–1076. [Google Scholar] [CrossRef]
- Zhang, W.; Guan, Y.J.; Bayliss, G.; Zhuang, S.G. Class IIa HDAC inhibitor TMP195 alleviates lipopolysaccharide-induced acute kidney injury. Am. J. Physiol.-Ren. Physiol. 2020, 319, F1015–F1026. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, C.; Qi, J. Activation of HDAC4 and GR signaling contributes to stress-induced hyperalgesia in the medial prefrontal cortex of rats. Brain Res. 2020, 1747, 147051. [Google Scholar] [CrossRef]
- Ko, J.Y.; Chuang, P.C.; Chen, M.W.; Ke, H.C.; Wu, S.L.; Chang, Y.H.; Chen, Y.S.; Wang, F.S. MicroRNA-29a ameliorates glucocorticoid-induced suppression of osteoblast differentiation by regulating beta-catenin acetylation. Bone 2013, 57, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Gao, Y.; Hu, N.; Wu, L.; Chen, Q. miR-365 Ameliorates Dexamethasone-Induced Suppression of Osteogenesis in MC3T3-E1 Cells by Targeting HDAC4. Int. J. Mol. Sci. 2017, 18, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielcarek, M.; Toczek, M.; Smeets, C.J.; Franklin, S.A.; Bondulich, M.K.; Jolinon, N.; Muller, T.; Ahmed, M.; Dick, J.R.; Piotrowska, I.; et al. HDAC4-myogenin axis as an important marker of HD-related skeletal muscle atrophy. PLoS Genet 2015, 11, e1005021. [Google Scholar] [CrossRef]
- Moresi, V.; Williams, A.H.; Meadows, E.; Flynn, J.M.; Potthoff, M.J.; McAnally, J.; Shelton, J.M.; Backs, J.; Klein, W.H.; Richardson, J.A.; et al. Myogenin and class II HDACs control neurogenic muscle atrophy by inducing E3 ubiquitin ligases. Cell 2010, 143, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holecek, M. Muscle wasting in animal models of severe illness. Int. J. Exp. Pathol. 2012, 93, 157–171. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control, n = 9 | LPS, n = 8 | |
|---|---|---|
| Body weight gain (g/24 h.) | −2.0 ± 0.9 | −21.9 ± 3.6 ** |
| Diaphragm (mg/100 g b.w.) | 206 ± 15 | 232 ± 17 |
| Gastrocnemius (mg/100 g b.w.) | 520 ± 20 | 517 ± 10 |
| Urea (mg/dL) | 22.1 ± 1.7 | 49.8 ± 5.9 ** |
| Creatinine (mg/dL) | 0.208 ± 0.008 | 0.331 ± 0.03 ** |
| Glucose (mg/dL) | 234 ± 8 | 180 ± 16 ** |
| Lactate (mmol/L) | 1.6 ± 0.2 | 5.6 ± 0.8 ** |
| HCO3- (mmol/L) | 26.0 ± 0.6 | 18.0 ± 2.2 ** |
| PaCO2 (mmHg) | 42.7 ± 3.9 | 37.8 ± 5.6 |
| PaO2 (mmHg) | 86.0 ± 6.1 | 88.0 ± 6.6 |
| SO2% (mmol/L) | 95.0 ± 1.3 | 95 ± 0.9 |
| pH | 7.39 ± 0.03 | 7.30 ± 0.04 |
| Gene | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| 18 S | GGTGCATGGCCGTTCTTA | TCGTTCGTTATCGGAATTAACC |
| IL10 | AGTGGAGCAGGTGAAGAATGA | TCATGGCCTTGTAGACACCTT |
| IL6 | GGAAGTTGGGGTAGGAAGGA | CCTGGAGTTTGTGAAGAACAACT |
| TNFα | TGAACTTCGGGGTGATCG | GGGCTTGTCACTCGAGTTTT |
| SOCS3 | CCTCCAGCATCTTTGTCGGAAGAC | CATTCGGGAGTTCCTGGACCAGTA |
| MuRF1 | TGTCTGGAGGTCGTTTCCG | AAGTGATCATGGACCGGCAT |
| Atrogin-1 | GAACAGCAAAACCAAAACTCAGTA | GCTCCTTAGTACTCCCTTTGTGAA |
| IGF-1 Ea | GCTATGGCTCCAGCATTCG | GGATGAGTGTTGCTTCCGGA |
| IGFBP3 | GGAAAGACGACGTGCATTG | GCGTATTTGAGCTCCACGTT |
| IGF1R | GCCTCCAACTTTGTCTTTGC | TCACTGGGCCAGGAATGT |
| HDAC4 | CACACCTCTTGGAGGGTACAA | AGCCCATCAGCTGTTTTGTC |
| Myogenin | CCTTGCTCAGCTCCCTCA | TGGGAGTTGCATTCACTGG |
| LC3b | CAGGTTGCCTAGCAGAGGTC | TGTCCTATACACCTGACCTGTTTC |
| GR | AAGAGCAGTGGAAGGACAGC | GCTGGGCAGTTTTTCCTTCG |
| KLF15 | TTGTGGGCCAGAAGTTCC | TGCATTTGTGCATTTTGAGAA |
| REDD1 | CCAGAGAAGAGGGCCTTGA | CCATCCAGGTATGAGGAGTCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Rupérez, Á.; Priego, T.; González-Nicolás, M.Á.; López-Calderón, A.; Lázaro, A.; Martín, A.I. Role of Glucocorticoid Signaling and HDAC4 Activation in Diaphragm and Gastrocnemius Proteolytic Activity in Septic Rats. Int. J. Mol. Sci. 2022, 23, 3641. https://doi.org/10.3390/ijms23073641
Moreno-Rupérez Á, Priego T, González-Nicolás MÁ, López-Calderón A, Lázaro A, Martín AI. Role of Glucocorticoid Signaling and HDAC4 Activation in Diaphragm and Gastrocnemius Proteolytic Activity in Septic Rats. International Journal of Molecular Sciences. 2022; 23(7):3641. https://doi.org/10.3390/ijms23073641
Chicago/Turabian StyleMoreno-Rupérez, Álvaro, Teresa Priego, María Ángeles González-Nicolás, Asunción López-Calderón, Alberto Lázaro, and Ana Isabel Martín. 2022. "Role of Glucocorticoid Signaling and HDAC4 Activation in Diaphragm and Gastrocnemius Proteolytic Activity in Septic Rats" International Journal of Molecular Sciences 23, no. 7: 3641. https://doi.org/10.3390/ijms23073641