
Fascinating Molecular and Immune Escape Mechanisms in the Treatment of STIs (Syphilis, Gonorrhea, Chlamydia, and Herpes Simplex)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Host Interactions
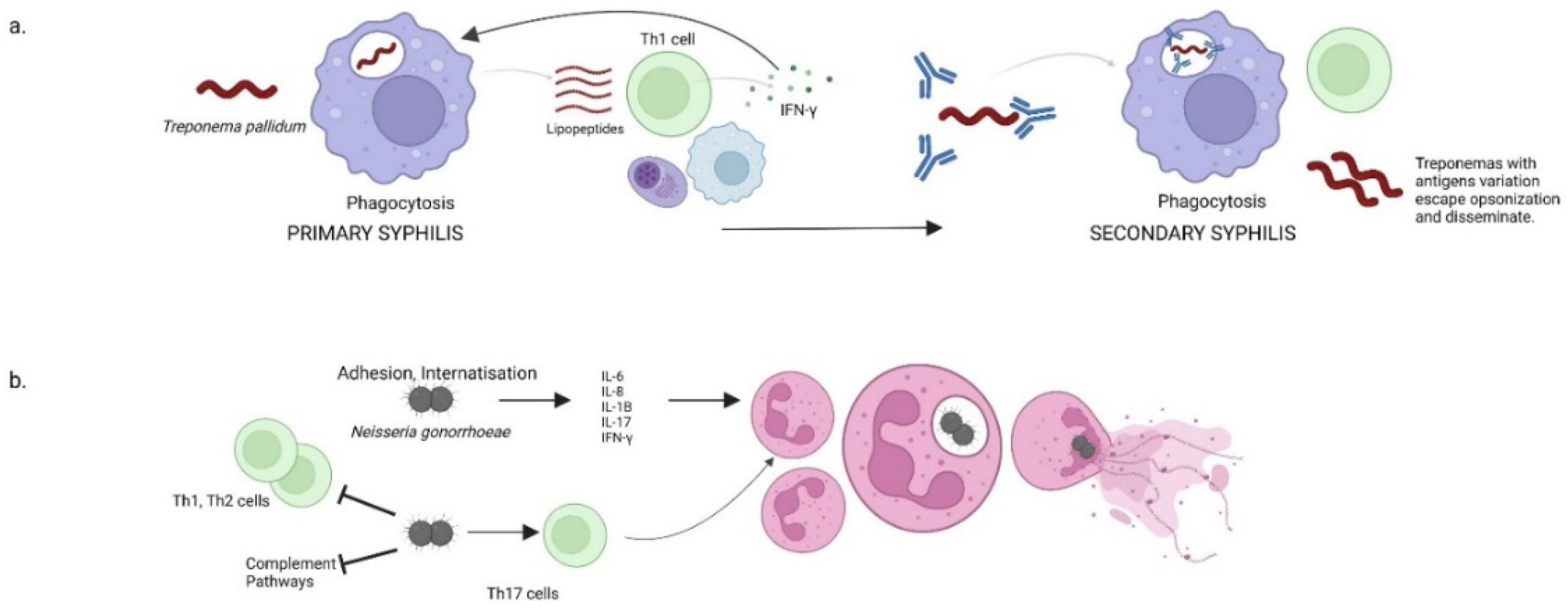
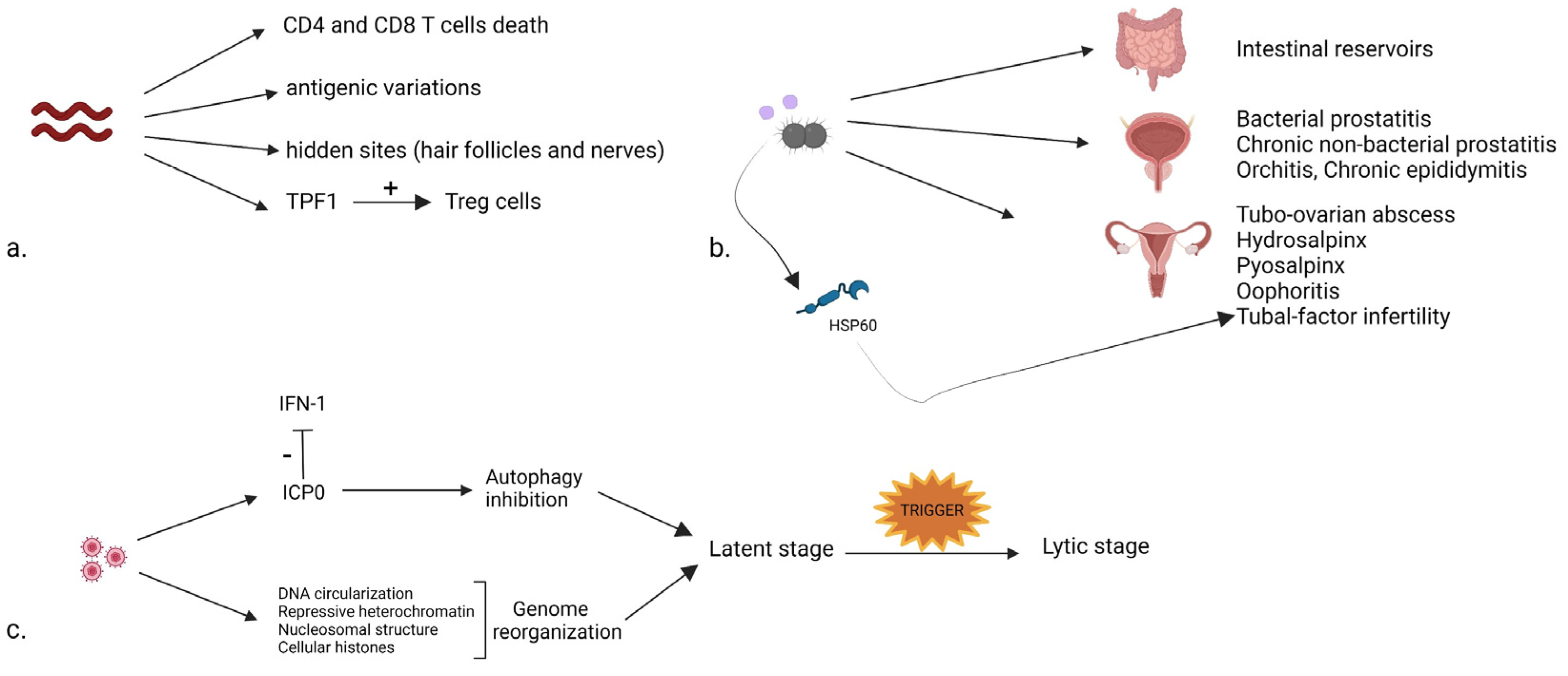
2.1. Syphilis
2.2. Gonorrhea
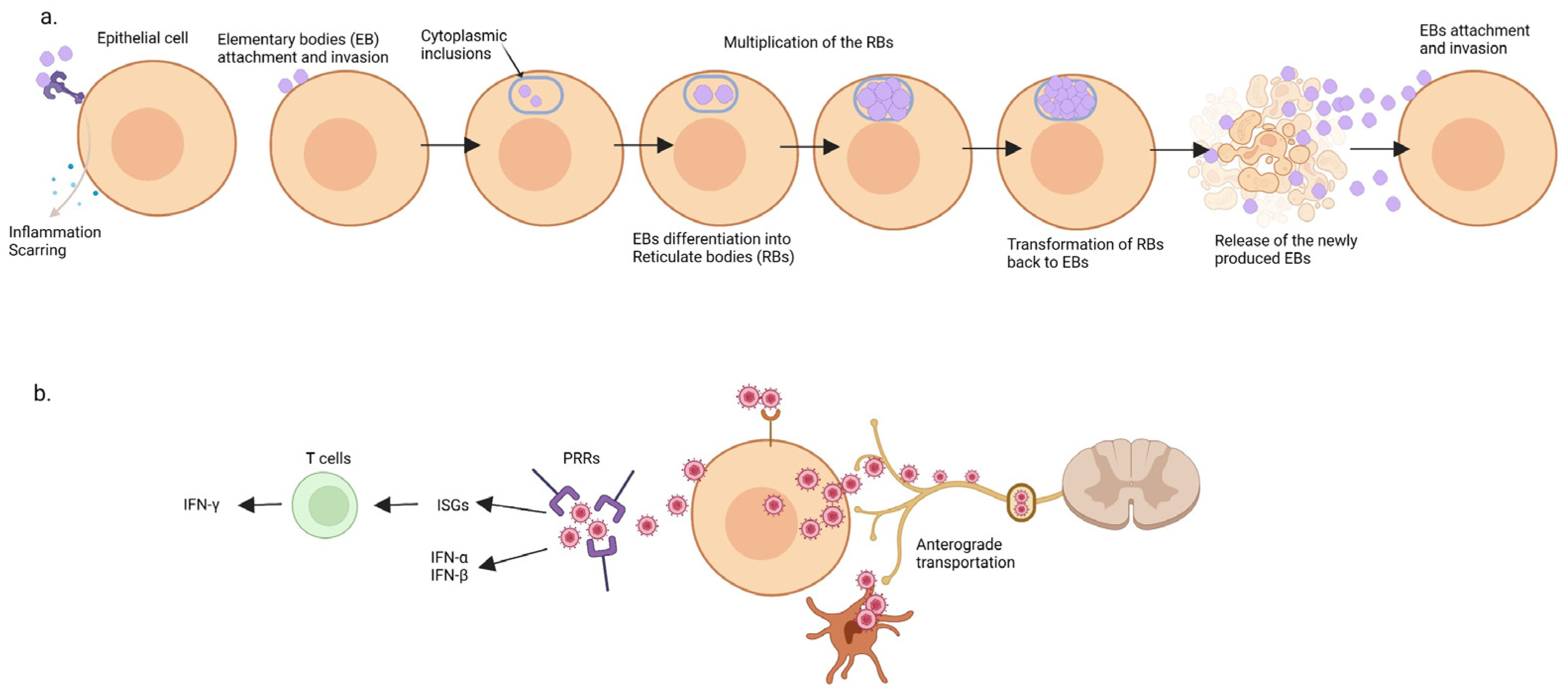
2.3. Chlamydia trachomatis
2.4. Herpes Simplex Virus
3. Treatment Guidelines and Resistant Pathogens
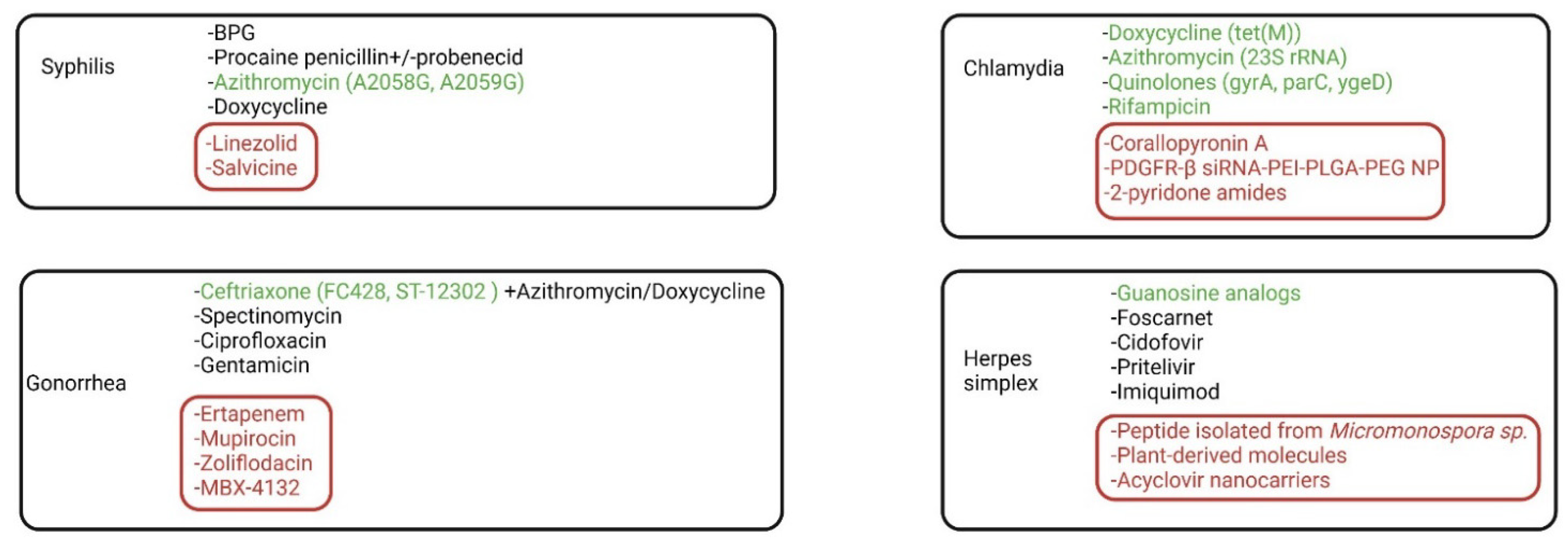
3.1. Syphilis
3.2. Gonorrhea and Chlamydia
3.3. Herpes Simplex
4. Novel Drugs and Nano Molecules
4.1. Syphilis
4.2. Gonorrhea
4.3. Chlamydia
4.4. Herpes Simplex
5. Conclusions and Future Research Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Eisinger, R.W.; Erbelding, E.; Fauci, A.S. Refocusing Research on Sexually Transmitted Infections. J. Infect. Dis. 2020, 222, 1432–1434. [Google Scholar] [CrossRef] [PubMed]
- Sexually Transmitted Disease Surveillance, Centers for Disease Control and Prevention. 2017. Available online: https://www.cdc.gov/std/stats17/2017-STD-Surveillance-Report_CDC-clearance-9.10.18.pdf (accessed on 15 November 2021).
- James, C.; Harfouche, M.; Welton, N.J.; Turner, K.M.E.; Abu-Raddad, L.J.; Gottlieb, S.L.; Looker, K.J. Herpes simplex virus: Global infection prevalence and incidence estimates, 2016. Bull. World Health Organ. 2020, 98, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Tien, V.; Punjabi, C.; Holubar, M.K. Antimicrobial resistance in sexually transmitted infections. J. Travel Med. 2020, 27, 101. [Google Scholar] [CrossRef]
- LaFond, R.E.; Molini, B.J.; Van Voorhis, W.C.; Lukehart, S.A. Antigenic Variation of TprK V Regions Abrogates Specific Antibody Binding in Syphilis. Infect. Immun. 2006, 74, 6244–6251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekelesidis, T. The Cross-Talk between Spirochetal Lipoproteins and Immunity. Front. Immunol. 2014, 5, 310. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A. The Gonococcus fights back: Is this time a knock out? Sex. Transm. Infect. 2010, 86, 415–421. [Google Scholar] [CrossRef]
- Unemo, M.; Lahra, M.M.; Cole, M.; Galarza, P.; Ndowa, F.; Martin, I.; Dillon, J.-A.R.; Ramon-Pardo, P.; Bolan, G.; Wi, T. World Health Organization Global Gonococcal Antimicrobial Surveillance Program (WHO GASP): Review of new data and evidence to inform international collaborative actions and research efforts. Sex. Health 2019, 16, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Aitolo, G.L.; Adeyemi, O.S.; Afolabi, B.L.; Owolabi, A.O. Neisseria gonorrhoeae Antimicrobial Resistance: Past to Present to Future. Curr. Microbiol. 2021, 78, 867–878. [Google Scholar] [CrossRef]
- Mestrovic, T.; Ljubin-Sternak, S. Molecular mechanisms of Chlamydia trachomatis resistance to antimicrobial drugs. Front. Biosci. 2018, 23, 656–670. [Google Scholar]
- Cossé, M.M.; Hayward, R.D.; Subtil, A. One Face of Chlamydia trachomatis: The Infectious Elementary Body. Curr. Top. Microbiol. Immunol. 2018, 412, 35–58. [Google Scholar]
- Whitley, R.; Baines, J. Clinical management of herpes simplex virus infections: Past, present, and future. F1000Research 2018, 7, 1726. [Google Scholar] [CrossRef] [Green Version]
- Bacon, T.H.; Levin, M.J.; Leary, J.J.; Sarisky, R.T.; Sutton, D. Herpes Simplex Virus Resistance to Acyclovir and Penciclovir after Two Decades of Antiviral Therapy. Clin. Microbiol. Rev. 2003, 16, 114–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lolis, M.S.; González, L.; Cohen, P.J.; A Schwartz, R. Drug-resistant herpes simplex virus in HIV infected patients. Acta Dermatovenerol. Croat. 2008, 16, 204–212. [Google Scholar] [PubMed]
- Osias, E.; Hung, P.; Giacani, L.; Stafylis, C.; Konda, K.A.; Vargas, S.K.; Reyes-Díaz, E.M.; Comulada, W.S.; Haake, D.A.; Haynes, A.M.; et al. Investigation of syphilis immunology and Treponema pallidum subsp. pallidum biology to improve clinical management and design a broadly protective vaccine: Study protocol. BMC Infect. Dis. 2020, 20, 444. [Google Scholar] [CrossRef]
- Fraser, C.M.; Norris, S.J.; Weinstock, G.M.; White, O.; Sutton, G.G.; Dodson, R.; Gwinn, M.; Hickey, E.K.; Clayton, R.; Ketchum, K.A.; et al. Complete Genome Sequence of Treponema pallidum, the Syphilis Spirochete. Science 1998, 281, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Radolf, J.D.; Lukehart, S.A. Immunology of Syphilis. In Pathogenic Treponemes: Cellular and Molecular Biology; Caister Academic Press: Norfolk, UK, 2006; pp. 285–322. [Google Scholar]
- Tomson, F.L.; Conley, P.G.; Norgard, M.V.; Hagman, K.E. Assessment of cell-surface exposure and vaccinogenic potentials of Treponema pallidum candidate outer membrane proteins. Microbes Infect. 2007, 9, 1267–1275. [Google Scholar] [CrossRef] [Green Version]
- Radolf, J.D.; Tramont, E.C.; Salazar, J.C. Mandell, Douglas and Bennett’s Principles and Practice of Infectious Diseases; Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; Churchill Livingtone Elsevier: Philadelphia, PA, USA, 2014; pp. 2684–2709. [Google Scholar]
- Grosenbaugh, D.A.; Rissi, D.R.; Krimer, P.M. Demonstration of the ability of a canine Lyme vaccine to reduce the incidence of histological synovial lesions following experimentally-induced canine Lyme borreliosis. Veter. Immunol. Immunopathol. 2016, 180, 29–33. [Google Scholar] [CrossRef]
- Edmondson, D.G.; Norris, S.J. In Vitro Cultivation of the Syphilis Spirochete Treponema pallidum. Curr. Protoc. 2021, 1, e44. [Google Scholar] [CrossRef]
- Radolf, J.D.; Deka, R.K.; Anand, A.; Šmajs, D.; Norgard, M.V.; Yang, X.F. Treponema pallidum, the syphilis spirochete: Making a living as a stealth pathogen. Nat. Rev. Microbiol. 2016, 14, 744–759. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.W.; Cruz, A.R.; LaVake, C.J.; Marzo, A.L.; Eggers, C.H.; Salazar, J.C.; Radolf, J.D. Phagocytosis of Borrelia burgdorferi and Treponema pallidum Potentiates Innate Immune Activation and Induces Gamma Interferon Production. Infect. Immun. 2007, 75, 2046–2062. [Google Scholar] [CrossRef] [Green Version]
- Leader, B.T.; Godornes, C.; VanVoorhis, W.C.; Lukehart, S.A. CD4+ Lymphocytes and Gamma Interferon Predominate in Local Immune Responses in Early Experimental Syphilis. Infect. Immun. 2007, 75, 3021–3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sell, S.; Salman, J.; Norris, S.J. Reinfection of chancre-immune rabbits with Treponema pallidum. I. Light and immunofluorescence studies. Am. J. Pathol. 1985, 118, 248–255. [Google Scholar] [PubMed]
- Ho, E.L.; Lukehart, S.A. Syphilis: Using modern approaches to understand an old disease. J. Clin. Investig. 2011, 121, 4584–4592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christodoulides, M. Preparation of Lipooligosaccharide (LOS) from Neisseria gonorrhoeae. Methods Mol. Biol. 2019, 1997, 87–96. [Google Scholar] [PubMed]
- Gulati, S.; Shaughnessy, J.; Ram, S.; Rice, P.A. Targeting Lipooligosaccharide (LOS) for a Gonococcal Vaccine. Front. Immunol. 2019, 10, 321. [Google Scholar] [CrossRef]
- Schneider, H.; Hale, T.L.; Zollinger, W.D.; Seid, R.C.; A Hammack, C.; Griffiss, J.M. Heterogeneity of molecular size and antigenic expression within lipooligosaccharides of individual strains of Neisseria gonorrhoeae and Neisseria meningitidis. Infect. Immun. 1984, 45, 544–549. [Google Scholar] [CrossRef] [Green Version]
- Criss, A.K.; Seifert, H.S. A bacterial siren song: Intimate interactions between Neisseria and neutrophils. Nat. Rev. Microbiol. 2012, 10, 178–190. [Google Scholar] [CrossRef]
- Calton, C.M.; Wade, L.K.; So, M. Upregulation of ATF3 inhibits expression of the pro-inflammatory cytokine IL-6 duringNeisseria gonorrhoeaeinfection. Cell. Microbiol. 2013, 15, 1837–1850. [Google Scholar] [CrossRef] [Green Version]
- Jayasundara, P.; Regan, D.G.; Seib, K.L.; Jayasundara, D.; Wood, J.G. Modelling the in-host dynamics of Neisseria gonorrhoeae infection. Pathog Dis. 2019, 77, ftz008. [Google Scholar] [CrossRef] [PubMed]
- Feinen, B.; Russell, M.W. Contrasting Roles of IL-22 and IL-17 in Murine Genital Tract Infection by Neisseria gonorrhoeae. Front. Immunol. 2012, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Handing, J.W.; Ragland, S.A.; Bharathan, U.V.; Criss, A.K. The MtrCDE Efflux Pump Contributes to Survival of Neisseria gonorrhoeae From Human Neutrophils and Their Antimicrobial Components. Front. Microbiol. 2018, 9, 2688. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.L.; Apicella, M.A. The Molecular Mechanisms Used by Neisseria gonorrhoeae To Initiate Infection Differ between Men and Women. Clin. Microbiol. Rev. 2004, 17, 965–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.O.; Wan, Y.Y.; Sanjabi, S.; Robertson, A.K.; Flavell, R.A. Transforming growth factor-beta regulation of immune responses. Annu. Rev. Immunol. 2006, 24, 99–146. [Google Scholar] [PubMed]
- Liu, Y.; Russell, M.W. Diversion of the Immune Response to Neisseria gonorrhoeae from Th17 to Th1/Th2 by Treatment with Anti-Transforming Growth Factor β Antibody Generates Immunological Memory and Protective Immunity. mBio 2011, 2, e00095-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghilardi, N.; Ouyang, W. Targeting the development and effector functions of TH17 cells. Semin. Immunol. 2007, 19, 383–393. [Google Scholar] [CrossRef]
- Zhu, W.; Ventevogel, M.S.; Knilans, K.J.; Anderson, J.E.; Oldach, L.M.; McKinnon, K.P.; Hobbs, M.M.; Sempowski, G.D.; Duncan, J.A. Neisseria gonorrhoeae Suppresses Dendritic Cell-Induced, Antigen-Dependent CD4 T Cell Proliferation. PLoS ONE 2012, 7, e41260. [Google Scholar] [CrossRef] [Green Version]
- Escobar, A.; Candia, E.; Reyes-Cerpa, S.; Villegas-Valdes, B.; Neira, T.; Lopez, M.; Maisey, K.; Tempio, F.; Ríos, M.; Acuña-Castillo, C.; et al. Neisseria gonorrhoeaeInduces a Tolerogenic Phenotype in Macrophages to Modulate Host Immunity. Mediat. Inflamm. 2013, 2013, 127017. [Google Scholar] [CrossRef] [Green Version]
- Stamm, W.E.; Batteiger, B.E. Chlamydia trachomatis trachoma, perinatal infections, lymphogranuloma venereum, and other genital infections. Princ. Pract. Infect. Dis. 2010, 72, 2443–2461. [Google Scholar]
- Carey, A.J.; Beagley, K. Chlamydia trachomatis, a Hidden Epidemic: Effects on Female Reproduction and Options for Treatment. Am. J. Reprod. Immunol. 2010, 63, 576–586. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, Y.M.; Belland, R.J. The chlamydial developmental cycle. FEMS Microbiol. Rev. 2005, 29, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Carabeo, R.A.; Hackstadt, T. Isolation and Characterization of a Mutant Chinese Hamster Ovary Cell Line That Is Resistant to Chlamydia trachomatis Infection at a Novel Step in the Attachment Process. Infect. Immun. 2001, 69, 5899–5904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Caldwell, H.D. Sulfated polysaccharides and a synthetic sulfated polymer are potent inhibitors of Chlamydia trachomatis infectivity in vitro but lack protective efficacy in an in vivo murine model of Chlamydial genital tract infection. Infect. Immun. 1998, 66, 1258–1260. [Google Scholar] [PubMed]
- Taraktchoglou, M.; Pacey, A.A.; Turnbull, J.E.; Eley, A. Infectivity of Chlamydia trachomatis Serovar LGV but Not E Is Dependent on Host Cell Heparan Sulfate. Infect. Immun. 2001, 69, 968–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, A.; Hocking, J.S.; Kong, F.Y. Rectal chlamydia infections: Implications for reinfection risk, screening, and treatment guidelines. Curr. Opin. Infect. Dis. 2021, 35, 42–48. [Google Scholar] [CrossRef]
- Witkin, S.S.; Minis, E.; Athanasiou, A.; Leizer, J.; Linhares, I.M. Chlamydia trachomatis: The Persistent Pathogen. Clin. Vaccine Immunol. 2017, 24, e00203-17. [Google Scholar] [CrossRef] [Green Version]
- Bavoil, P.M.; Hsia, R.-C.; Ojcius, D.M. Closing in on Chlamydia and its intracellular bag of tricks. Microbiology 2000, 146, 2723–2731. [Google Scholar] [CrossRef] [Green Version]
- Scidmore, M.A.; Rockey, D.D.; Fischer, E.R.; Heinzen, R.A.; Hackstadt, T. Vesicular interactions of the Chlamydia trachomatis inclusion are determined by chlamydial early protein synthesis rather than route of entry. Infect. Immun. 1996, 64, 5366–5372. [Google Scholar] [CrossRef] [Green Version]
- Grieshaber, S.; Grieshaber, N.; Yang, H.; Baxter, B.; Hackstadt, T.; Omsland, A. Impact of Active Metabolism on Chlamydia trachomatis Elementary Body Transcript Profile and Infectivity. J. Bacteriol. 2018, 200, 65–83. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Bavoil, P. (Eds.) Intracellular Pathogens I: Chlamydiales; American Society for Microbiology Press: Washington, DC, USA, 2012. [Google Scholar]
- Murthy, A.K.; Arulanandam, B.P.; Zhong, G. Chlamydia Vaccine: Progress and Challenges. Intracell. Pathog. Chlamydia I 2014, 311–333. [Google Scholar] [CrossRef]
- Bastidas, R.J.; Elwell, C.A.; Engel, J.N.; Valdivia, R.H. Chlamydial Intracellular Survival Strategies. Cold Spring Harb. Perspect. Med. 2013, 3, a010256. [Google Scholar] [CrossRef] [Green Version]
- Hybiske, K.; Stephens, R.S. Mechanisms of host cell exit by the intracellular bacterium Chlamydia. Proc. Natl. Acad. Sci. USA 2007, 104, 11430–11435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, K.; Crother, T.R.; Arditi, M. Innate immune responses to Chlamydia pneumoniae infection: Role of TLRs, NLRs, and the inflammasome. Microbes Infect. 2012, 14, 1301–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scurtu, L.G.; Simionescu, O. Soluble Factors and Receptors Involved in Skin Innate Immunity—What Do We Know So Far? Biomedicines 2021, 9, 1795. [Google Scholar] [CrossRef] [PubMed]
- Itoh, R. Chlamydia pneumoniae harness host NLRP3 inflammasome-mediated caspase-1 activation for optimal intracellular growth in murine macrophages. Biochem. Biophys. Res. Commun. 2014, 452, 689–694. [Google Scholar] [CrossRef]
- Groves, M.J. Genital Herpes: A Review. Am. Fam. Physician 2016, 93, 928–934. [Google Scholar]
- Cunningham, A.L.; Diefenbach, R.; Miranda-Saksena, M.; Bosnjak, L.; Kim, M.; Jones, C.; Douglas, M.W. The Cycle of Human Herpes Simplex Virus Infection: Virus Transport and Immune Control. J. Infect. Dis. 2006, 194, S11–S18. [Google Scholar] [CrossRef] [Green Version]
- Lamers, S.L.; Newman, R.M.; Laeyendecker, O. Global Diversity within and between Human Herpesvirus 1 and 2 Glycoproteins. J. Virol. 2015, 89, 8206–8218. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, R.J.; Atanasiu, D.; Cairns, T.M.; Gallagher, J.R.; Krummenacher, C.; Cohen, G.H. Herpes Virus Fusion and Entry: A Story with Many Characters. Viruses 2012, 4, 800–832. [Google Scholar] [CrossRef]
- Holmes, T.D.; Wilson, E.B. Licensed human natural killer cells aid dendritic cell maturation via TNFSF14/LIGHT. Proc. Natl. Acad. Sci. USA 2014, 111, E5688–E5696. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Bai, Q.; Baek, H.J. Soluble V domain of nectin-1/HveC enables entry of herpes simplex virus type 1 (HSV-1) into HSV-resistant cells by binding to viral glycoprotein D. J. Virol. 2006, 80, 138–148. [Google Scholar]
- Desai, P.; Abboud, G.; Stanfield, J. HVEM Imprints Memory Potential on Effector CD8 T Cells Required for Protective Mucosal Immunity. J. Immunol. 2017, 199, 2968–2975. [Google Scholar] [CrossRef] [PubMed]
- Wakeley, M.E.; Shubin, N.J.; Monaghan, S.F.; Gray, C.C.; Ayala, A.; Heffernan, D.S. Herpes Virus Entry Mediator (HVEM): A Novel Potential Mediator of Trauma-Induced Immunosuppression. J. Surg. Res. 2020, 245, 610–618. [Google Scholar] [CrossRef]
- Chew, T.; Taylor, K.E.; Mossman, K.L. Innate and Adaptive Immune Responses to Herpes Simplex Virus. Viruses 2009, 1, 979–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, K.P.; Wu, S.; Wigdahl, B.; Jennings, S.R. Immunological control of herpes simplex virus infections. J. Neurovirol. 2013, 19, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M.L.; Sheridan, B.S.; Bonneau, R.H. Psychological Stress Compromises CD8+T Cell Control of Latent Herpes Simplex Virus Type 1 Infections. J. Immunol. 2007, 179, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mork, N.J.; Kofodolsen, E.; Sorensen, K.; Bach, E.; Orntoft, T.F.; Østergaard, L.; Paludan, S.R.; Christiansen, M.; Mogensen, T.H. Mutations in the TLR3 signaling pathway and beyond in adult patients with herpes simplex encephalitis. Genes Immun. 2015, 16, 552–566. [Google Scholar] [CrossRef] [Green Version]
- Janier, M.; Unemo, M.; Dupin, N.; Tiplica, G.S.; Potočnik, M.; Patel, R. 2020 European guideline on the management of syphilis. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 574–588. [Google Scholar] [CrossRef]
- Kingston, M.; French, P.; Higgins, S.; McQuillan, O.; Sukthankar, A.; Stott, C.J.T.; McBrien, B.; Tipple, C.; Turner, A.J.; Sullivan, A.K.; et al. UK national guidelines on the management of syphilis. Int. J. STD AIDS 2016, 27, 421–446. [Google Scholar] [CrossRef]
- Stamm, L.V. Syphilis: Antibiotic treatment and resistance. Epidemiol. Infect. 2015, 143, 1567–1574. [Google Scholar] [CrossRef] [Green Version]
- Šmajs, D.; Grillová, L.; Paštěková, L. Macrolide Resistance in the Syphilis Spirochete, Treponema pallidum ssp. pallidum: Can We Also Expect Macrolide-Resistant Yaws Strains? Am. J. Trop. Med. Hyg. 2015, 93, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Eguiluz, M.; Vargas, S.; Giacani, L.; Reyes-Diaz, M.; Konda, K.; Caceres, C.; Klausner, J. P229 Prevalence of macrolide and tetracycline resistant Treponema pallidum strains in syphilis cases. PeruSex. Transm. Infect. 2021, 97, 120. [Google Scholar]
- Sanchez, A.; Mayslich, C.; Malet, I.; Grange, P.; Janier, M.; Saule, J.; Martinet, P.; Robert, J.; Moulene, D.; Truchetet, F.; et al. Surveillance of Antibiotic Resistance Genes in Treponema Pallidum Subspecies Pallidum from Patients with Early Syphilis in France. Acta Derm. Venereol. 2020, 100, 221. [Google Scholar] [CrossRef]
- Stamm, L.V. Global Challenge of Antibiotic-Resistant Treponema pallidum. Antimicrob. Agents Chemother. 2010, 54, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myint, M.; Bashiri, H.; Harrington, R.D.; Marra, C.M. Relapse of secondary syphilis after benzathine penicillin G: Molecular analysis. Sex. Transm. Dis. 2004, 31, 196–205. [Google Scholar]
- Cruz, A.R. Secondary Syphilis in Cali, Colombia: New Concepts in Disease Pathogenesis. PLoS Neglected Trop. Dis. 2010, 4, e690. [Google Scholar] [CrossRef]
- Xia, W.; Zhao, J.; Su, B. Syphilitic infection impairs immunity by inducing both apoptosis and pyroptosis of CD4+ and CD8+ T lymphocytes. Innate Immun. 2021, 27, 99–106. [Google Scholar] [CrossRef]
- Babolin, C.; Amedei, A.; Ozoliņš, D.; Žileviča, A.; D’elios, M.M.; de Bernard, M. TpF1 fromTreponema pallidumActivates Inflammasome and Promotes the Development of Regulatory T Cells. J. Immunol. 2011, 187, 1377–1384. [Google Scholar] [CrossRef] [Green Version]
- Pozzobon, T.; Facchinello, N.; Bossi, F.; Capitani, N.; Benagiano, M.; DiBenedetto, G.; Zennaro, C.; West, N.; Codolo, G.; Bernardini, M.; et al. Treponema pallidum (syphilis) antigen TpF1 induces angiogenesis through the activation of the IL-8 pathway. Sci. Rep. 2016, 6, 18785. [Google Scholar]
- Unemo, M.; Ross, J.; Serwin, A.B.; Gomberg, M.; Cusini, M.; Jensen, J.S. 2020 European guideline for the diagnosis and treatment of gonorrhoea in adults. Int. J. STD AIDS 2020. [Google Scholar] [CrossRef]
- Kanesaka, I.; Ohno, A.; Katsuse, A.K.; Takahashi, H.; Kobayashi, I. The emergence of the ceftriaxone-resistant Neisseria gonorrhoeae FC428 clone by transfer of resistance from an oral Neisseria subflava reservoir of resistance. J. Antimicrob. Chemother. 2021, 77, 390. [Google Scholar] [CrossRef]
- Lin, X.; Chen, W.; Xie, Q.; Yu, Y.; Liao, Y.; Feng, Z.; Qin, X.; Wu, X.; Tang, S.; Zheng, H. Dissemination and genome analysis of high-level ceftriaxone-resistant penA 60.001 Neisseria gonorrhoeae strains from the Guangdong Gonococcal antibiotics susceptibility Programme (GD-GASP), 2016–2019. Emerg. Microbes Infect. 2022, 11, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Chen, S.-C.; Yang, F.; Van Der Veen, S.; Yin, Y.-P. Impact of the gonococcal FC428 penA allele 60.001 on ceftriaxone resistance and biological fitness. Emerg. Microbes Infect. 2020, 9, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Sawatzky, P.; Demczuk, W.; Lefebvre, B.; Allen, V.; Diggle, M.; Hoang, L.; Van Caeseele, P.; Haldane, D.; Minion, J.; Mulvey, M.R.; et al. Increasing azithromycin resistance in Neisseria gonorrhoeae due to NG-MAST 12302 clonal spread in Canada, 2015–2018. Antimicrob. Agents Chemother. 2022, 66, e01688-21. [Google Scholar] [CrossRef]
- Carballo, R.; Povoa, M.C.; Abad, R.; Navarro, C.; Martin, E.; Alvarez, M.; Salgado, A.; Potel, C. Large Increase in Azithromycin-Resistant Neisseria gonorrhoeae in Northern Spain. Microb. Drug Resist. 2021, 28, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Cephalosporin-Resistant Neisseria gonorrhoeae Public Health Response Plan; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2012; pp. 1–43.
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. Available online: http://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (accessed on 15 December 2021).
- Lanjouw, E.; Ouburg, S.; de Vries, H.J.; Stary, A.; Radcliffe, K.; Unemo, M. 2015 European guideline on the management of Chlamydia trachomatis infections. Int. J. STD AIDS 2016, 27, 333–348. [Google Scholar] [CrossRef]
- Rank, R.G.; Yeruva, L. Hidden in Plain Sight: Chlamydial Gastrointestinal Infection and Its Relevance to Persistence in Human Genital Infection. Infect. Immun. 2014, 82, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Yeruva, L.; Melnyk, S.; Spencer, N.; Bowlin, A.; Rank, R.G. Differential Susceptibilities to Azithromycin Treatment of Chlamydial Infection in the Gastrointestinal Tract and Cervix. Antimicrob. Agents Chemother. 2013, 57, 6290–6294. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.Y.S.; Tabrizi, S.N.; Fairley, C.K.; Vodstrcil, L.A.; Huston, W.M.; Chen, M.; Bradshaw, C.; Hocking, J.S. The efficacy of azithromycin and doxycycline for the treatment of rectal chlamydia infection: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2015, 70, 1290–1297. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.P.; Kong, F.Y.S.; Yeruva, L.; Hocking, J.S.; Rank, R.G.; Wilson, D.P.; Donovan, B. Is it time to switch to doxycycline from azithromycin for treating genital chlamydial infections in women? Modelling the impact of autoinoculation from the gastrointestinal tract to the genital tract. BMC Infect. Dis. 2015, 15, 200. [Google Scholar] [CrossRef] [Green Version]
- Benamri, I.; Azzouzi, M.; Sanak, K.; Moussa, A.; Radouani, F. An overview of genes and mutations associated with Chlamydiae species’ resistance to antibiotics. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 59. [Google Scholar] [CrossRef]
- Shao, L.; You, C.; Cao, J.; Jiang, Y.; Liu, Y.; Liu, Q. High treatment failure rate is better explained by resistance gene detection than by minimum inhibitory concentration in patients with urogenital Chlamydia trachomatis infection. Int. J. Infect. Dis. 2020, 96, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, J.; Frosch, M.; Vogel, U. Manual of Clinical Microbiology; ASM Press: Washington, DC, USA, 2011; Volume 1, pp. 559–569. [Google Scholar]
- Ala-Almohadesin, A.; Mohammadbeygi, M.; Bahavar, A.; Mohammadi, M.A.; Mohamadzadeh, N.; Abolhasani, M.; Dabiri, H. Molecular Detection of Pathogens Causing Sexually Transmissible Infections in Patients with Prostate Cancer and Hyperplasia by Quantitative TaqMan Real-Time PCR Assay. Clin. Lab. 2019, 65. [Google Scholar] [CrossRef]
- Lian, W.-Q.; Luo, F.; Song, X.-L.; Lu, Y.-J.; Zhao, S.-C. Gonorrhea and Prostate Cancer Incidence: An Updated Meta-Analysis of 21 Epidemiologic Studies. Med. Sci. Monit. 2015, 21, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutcliffe, S.B.; Nevin, R.; Pakpahan, R. Prostate involvement during sexually transmitted infections as measured by prostate-specific antigen concentration. Br. J. Cancer 2011, 105, 602–605. [Google Scholar] [CrossRef] [Green Version]
- Motrich, R.D.; Salazar, F.C.; Breser, M.L.; Mackern-Oberti, J.P.; Godoy, G.J.; Olivera, C.; Paira, D.A.; Rivero, V.E. Implications of prostate inflammation on male fertility. Andrologia 2018, 50, e13093. [Google Scholar] [CrossRef] [Green Version]
- Mackern-Oberti, J.P.; Motrich, R.D.; Breser, M.L.; Sánchez, L.R.; Cuffini, C.; Rivero, V.E. Chlamydia trachomatis infection of the male genital tract: An update. J. Reprod. Immunol. 2013, 100, 37–53. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Anyalechi, G.E.; Cohen, C.R.; Haggerty, C.L.; Manhart, L.E.; Hillier, S.L. Etiology and Diagnosis of Pelvic Inflammatory Disease: Looking Beyond Gonorrhea and Chlamydia. J. Infect. Dis. 2021, 224, S29–S35. [Google Scholar]
- Mohseni, M.; Simon, L.V.; Sheele, J.M. Epidemiologic and Clinical Characteristics of Tubo-Ovarian Abscess, Hydrosalpinx, Pyosalpinx, and Oophoritis in Emergency Department Patients. Cureus 2020, 1, e11647. [Google Scholar] [CrossRef]
- Kessler, M.; Zielecki, J.; Thieck, O.; Mollenkopf, H.-J.; Fotopoulou, C.; Meyer, T.F. Chlamydia trachomatis Disturbs Epithelial Tissue Homeostasis in Fallopian Tubes via Paracrine Wnt Signaling. Am. J. Pathol. 2012, 180, 186–198. [Google Scholar] [CrossRef]
- Smolarczyk, K.; Mlynarczyk-Bonikowska, B.; Rudnicka, E. The Impact of Selected Bacterial Sexually Transmitted Diseases on Pregnancy and Female Fertility. Int. J. Mol. Sci. 2021, 22, 2170. [Google Scholar] [CrossRef]
- Nelson, H.D.; Zakher, B.; Cantor, A.; Deagas, M.; Pappas, M. Screening for Gonorrhea and Chlamydia: Systematic Review to Update the U.S. Preventive Services Task Force Recommendations; Report No.: 13-05184-EF-PMID: 25356451; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2014.
- Van Ess, E.F.; Eck-Hauer, A.; Land, J.A.; Morré, S.A.; Ouburg, S. Combining individualChlamydia trachomatisIgG antibodies MOMP, TARP, CPAF, OMP2, and HSP60 for tubal factor infertility prediction. Am. J. Reprod. Immunol. 2019, 81, e13091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linhares, I.M.; Witkin, S.S. Immunopathogenic consequences of Chlamydia trachomatis 60 kDa heat shock protein expression in the female reproductive tract. Cell Stress Chaperones 2010, 15, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, A.Z.; Diamond, M.P.; Legro, R.S.; Schlaff, W.D.; Barnhart, K.T.; Casson, P.R.; Christman, G.M.; Alvero, R.; Hansen, K.R.; Geisler, W.M.; et al. Chlamydia trachomatis immunoglobulin G3 seropositivity is a predictor of reproductive outcomes in infertile women with patent fallopian tubes. Fertil. Steril. 2015, 104, 1522–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.; Kennedy, O.J.; Clarke, E.; Geretti, A.; Nilsen, A.; Lautenschlager, S.; Green, J.; Donders, G.; Van Der Meijden, W.; Gomberg, M.; et al. 2017 European guidelines for the management of genital herpes. Int. J. STD AIDS 2017, 28, 1366–1379. [Google Scholar] [CrossRef] [PubMed]
- Enescu, C.D.; Utz Petry, S.; Nartker, N.; Moossavi, M. Treatment of acyclovir-resistant herpes simplex virus with intralesional cidofovir. JAAD Case Rep. 2021, 13, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Cannon, L.; Tholouli, E.; Ward, C.; Farooq, H.; Kingston, M. Use of pritelivir in refractory aciclovir-resistant herpes simplex virus type 2. Int. J. STD AIDS 2021, 32, 978–980. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, N.; Donner, A.-K.; Gege, C.; Lange, F.; Kleymann, G.; Grunwald, T. Helicase primase inhibitors (HPIs) are efficacious for therapy of human herpes simplex virus (HSV) disease in an infection mouse model. Antivir. Res. 2021, 195, 105190. [Google Scholar] [CrossRef]
- de Oliveira, S.R.; Assoni, A.S.; Vargas, T.J.D.S.; Daxbacher, E. Acyclovir-resistant chronic mucocutaneous herpes with good response to the association with imiquimod in an AIDS patient: Case report. An. Bras. Dermatol. 2022, 97, 264–266. [Google Scholar] [CrossRef]
- Siqueira, S.M.; Gonçalves, B.B.; Loss, J.B.; Estrella, R.R. Vegetative chronic genital herpes with satisfactory response to imiquimod. An. Bras. Dermatol. 2019, 94, 221–223. [Google Scholar] [CrossRef] [Green Version]
- Verzosa, A.L.; McGeever, L.A.; Bhark, S.-J.; Delgado, T.; Salazar, N.; Sanchez, E.L. Herpes Simplex Virus 1 Infection of Neuronal and Non-Neuronal Cells Elicits Specific Innate Immune Responses and Immune Evasion Mechanisms. Front. Immunol. 2021, 12, 644664. [Google Scholar] [CrossRef]
- Strang, B.L.; Stow, N.D. Circularization of the Herpes Simplex Virus Type 1 Genome upon Lytic Infection. J. Virol. 2005, 79, 12487–12494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waisner, H.; Kalamvoki, M. The ICP0 Protein of Herpes Simplex Virus 1 (HSV-1) Downregulates Major Autophagy Adaptor Proteins Sequestosome 1 and Optineurin during the Early Stages of HSV-1 Infection. J. Virol. 2019, 93, 1258–1277. [Google Scholar] [CrossRef]
- Shahnazaryan, D.; Khalil, R.; Wynne, C.; Jefferies, C.A.; Ní Gabhann-Dromgoole, J.; Murphy, C.C. Herpes simplex virus 1 targets IRF7 via ICP0 to limit type I IFN induction. Sci. Rep. 2020, 10, 22216. [Google Scholar] [CrossRef]
- Suzich, J.B.; Cliffe, A.R. Strength in diversity: Understanding the pathways to herpes simplex virus reactivation. Virology 2018, 522, 81–91. [Google Scholar] [CrossRef]
- DuRaine, G.; Johnson, D.C. Anterograde transport of α-herpesviruses in neuronal axons. Virology 2021, 559, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Pegg, C.E.; Zaichick, S.V.; Bomba-Warczak, E.; Jovasevic, V.; Kim, D.; Kharkwal, H.; Wilson, D.W.; Walsh, D.; Sollars, P.J.; Pickard, G.E.; et al. Herpesviruses assimilate kinesin to produce motorized viral particles. Nature 2021, 599, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, D.G.; Hu, B.; Norris, S.J. Long-Term In Vitro Culture of the Syphilis Spirochete Treponema pallidum subsp. pallidum. mBio 2018, 9, e01153-18. [Google Scholar] [CrossRef] [Green Version]
- Haynes, A.M.; Giacani, L.; Mayans, M.V.; Ubals, M.; Nieto, C.; Pérez-Mañá, C.; Quintó, L.; Romeis, E.; Mitjà, O. Efficacy of linezolid on Treponema pallidum, the syphilis agent: A preclinical study. EBioMedicine 2021, 65, 103281. [Google Scholar] [CrossRef]
- Qin, Y.; Xu, L.; Teng, Y.; Wang, Y.; Ma, P. Discovery of novel antibacterial agents: Recent developments in D-alanyl-D-alanine ligase inhibitors. Chem. Biol. Drug Des. 2021, 98, 305–322. [Google Scholar] [CrossRef]
- Qing, C.; Zhang, J.S.; Ding, J. In vitro cytotoxicity of salvicine, a novel diterpenoid quinone. Zhongguo Yao Li Xue Bao. 1999, 20, 297–302. [Google Scholar]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2009, 19, 3533. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, U.N.; Tiwari, S.; Singh, P.; Singh, S.; Awasthi, M.; Pandey, V. Treponema pallidumPutative Novel Drug Target Identification and Validation: Rethinking Syphilis Therapeutics with Plant-Derived Terpenoids. OMICS 2015, 19, 104–114. [Google Scholar] [PubMed] [Green Version]
- Vries, H.D.; Laat, M.D.; Jongen, V.W.; Heijman, T.; Wind, C.M.; Boyd, A.; Korne-Elenbaas, J.D.; Dam, A.P.V. Efficacy of ertapenem, gentamicin, fosfomycin, and ceftriaxone for the treatment of anogenital gonorrhoea (NABOGO): A randomised, non-inferiority trial. Lancet Infect. Dis. 2022, 21, 625–628. [Google Scholar]
- Cern, A.; Bavli, Y.; Hod, A.; Zilbersheid, D.; Mushtaq, S.; Michael-Gayego, A.; Barasch, D.; Rotkopf, Y.F.; Moses, A.E.; Livermore, D.M.; et al. Therapeutic Potential of Injectable Nano-Mupirocin Liposomes for Infections Involving Multidrug-Resistant Bacteria. Pharmaceutics 2021, 13, 2186. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Nandi, N. Dynamics of the Catalytic Active Site of Isoleucyl tRNA Synthetase from Staphylococcus aureus bound with Adenylate and Mupirocin. J. Phys. Chem. B 2022, 126, 620–633. [Google Scholar] [CrossRef]
- O’donnell, J.; Lawrence, K.; Vishwanathan, K.; Hosagrahara, V.; Mueller, J.P. Single-Dose Phar-macokinetics, Excretion, and Metabolism of Zoliflodacin, a Novel Spiropyrimidinetrione Antibiotic, in Healthy Volunteers. Antimicrob. Agents Chemother. 2018, 63, e01808-18. [Google Scholar] [PubMed] [Green Version]
- Unemo, M.; Ahlstrand, J.; Sánchez-Busó, L.; Day, M.; Aanensen, D.; Golparian, D.; Jacobsson, S.; Cole, M.J.; European Collaborative Group. High susceptibility to zoliflodacin and conserved target (GyrB) for zoliflodacin among 1209 consecutive clinical Neisseria gonorrhoeae isolates from 25 European countries, 2018. J. Antimicrob. Chemother. 2021, 76, 1221–1228. [Google Scholar] [CrossRef]
- Jacobsson, S.; Golparian, D.; Oxelbark, J.; Alirol, E.; Franceschi, F.; Gustafsson, T.N.; Brown, D.; Louie, A.; Drusano, G.; Unemo, M. Pharmacodynamic Evaluation of Dosing, Bacterial Kill, and Resistance Suppression for Zoliflodacin Against Neisseria gonorrhoeae in a Dynamic Hollow Fiber Infection Model. Front. Pharmacol. 2021, 12, 682135. [Google Scholar] [CrossRef]
- Newman, L.M.; Kankam, M.; Nakamura, A.; Conrad, T.; Mueller, J.; O’donnell, J.; Osborn, B.L.; Gu, K.; Saviolakis, G.A. Thorough QT Study To Evaluate the Effect of Zoliflodacin, a Novel Therapeutic for Gonorrhea, on Cardiac Repolarization in Healthy Adults. Antimicrob. Agents Chemother. 2021, 65, e0129221. [Google Scholar] [CrossRef]
- Le, W.; Su, X.; Lou, X.; Li, X.; Gong, X.; Wang, B.; Genco, C.A.; Mueller, J.P.; Rice, P.A. Susceptibility Trends of Zoliflodacin against Multidrug-Resistant Neisseria gonorrhoeae Clinical Isolates in Nanjing, China, 2014 to 2018. Antimicrob. Agents Chemother. 2021, 65, e00863-20. [Google Scholar] [CrossRef]
- Aron, Z.D.; Mehrani, A.; Hoffer, E.D.; Connolly, K.L.; Srinivas, P.; Torhan, M.C.; Alumasa, J.N.; Cabrera, M.; Hosangadi, D.; Barbor, J.S.; et al. trans-Translation inhibitors bind to a novel site on the ribosome and clear Neisseria gonorrhoeae in vivo. Nat. Commun. 2021, 12, 1799. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Wen, Y.; Zhao, L.; Su, S.; Zhao, Y.; Lei, W.; Li, Z. Chlamydia trachomatis induces lncRNA MIAT upregulation to regulate mitochondria-mediated host cell apoptosis and chlamydial development. J. Cell. Mol. Med. 2022, 26, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Loeper, N.; Graspeuntner, S.; Ledig, S.; Kaufhold, I.; Hoellen, F.; Schiefer, A.; Henrichfreise, B.; Pfarr, K.; Hoerauf, A.; Shima, K.; et al. Elaborations on Corallopyronin A as a Novel Treatment Strategy Against Genital Chlamydial Infections. Front. Microbiol. 2019, 10, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shima, K.; Ledig, S.; Loeper, N.; Schiefer, A.; Pfarr, K.; Hoerauf, A.; Graspeuntner, S.; Rupp, J. Effective inhibition of rifampicin-resistant Chlamydia trachomatis by the novel DNA-dependent RNA polymerase inhibitor corallopyronin A. Int. J. Antimicrob. Agents 2018, 52, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Traore, Y.; Jimenez, C.; Ho, E.A. Autophagy induction and PDGFR-β-knockdown by siRNA-encapsulated nanoparticles reduce chlamydia trachomatis infection. Sci. Rep. 2019, 9, 1306. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Otero, C.; Bahnan, W.; Vielfort, K.; Silver, J.; Singh, P.; Elbir, H.; Almqvist, F.; Bergström, S. Gylfe Åsa A 2-Pyridone Amide Inhibitor of Transcriptional Activity in Chlamydia trachomatis. Antimicrob. Agents Chemother. 2021, 65, e01826-20. [Google Scholar] [CrossRef]
- Sureram, S.; Arduino, I.; Ueoka, R.; Rittà, M.; Francese, R.; Srivibool, R.; Darshana, D.; Piel, J.; Ruchirawat, S.; Muratori, L.; et al. The Peptide A-3302-B Isolated from a Marine Bacterium Micromonospora sp. Inhibits HSV-2 Infection by Preventing the Viral Egress from Host Cells. Int. J. Mol. Sci. 2022, 23, 947. [Google Scholar] [CrossRef]
- van de Sand, L.; Bormann, M.; Schmitz, Y.; Heilingloh, C.S.; Witzke, O.; Krawczyk, A. Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses. Viruses 2021, 13, 1386. [Google Scholar] [CrossRef]
- Akilesh, M.S.; Wadhwani, A. Novel Applications of Nanotechnology in Controlling HIV and HSV Infections. Curr. Drug. Res. Rev. 2021, 13, 120–129. [Google Scholar]
- Donalisio, M.; Argenziano, M.; Rittà, M.; Bastiancich, C.; Civra, A.; Lembo, D.; Cavalli, R. Acyclovir-loaded sulfobutyl ether-β-cyclodextrin decorated chitosan nanodroplets for the local treatment of HSV-2 infections. Int. J. Pharm. 2020, 587, 119676. [Google Scholar] [CrossRef]
- Chindamo, G.; Sapino, S.; Peira, E.; Chirio, D.; Gallarate, M. Recent Advances in Nanosystems and Strategies for Vaginal Delivery of Antimicrobials. Nanomaterials 2021, 11, 311. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scurtu, L.G.; Jinga, V.; Simionescu, O. Fascinating Molecular and Immune Escape Mechanisms in the Treatment of STIs (Syphilis, Gonorrhea, Chlamydia, and Herpes Simplex). Int. J. Mol. Sci. 2022, 23, 3550. https://doi.org/10.3390/ijms23073550
Scurtu LG, Jinga V, Simionescu O. Fascinating Molecular and Immune Escape Mechanisms in the Treatment of STIs (Syphilis, Gonorrhea, Chlamydia, and Herpes Simplex). International Journal of Molecular Sciences. 2022; 23(7):3550. https://doi.org/10.3390/ijms23073550
Chicago/Turabian StyleScurtu, Lucian G., Viorel Jinga, and Olga Simionescu. 2022. "Fascinating Molecular and Immune Escape Mechanisms in the Treatment of STIs (Syphilis, Gonorrhea, Chlamydia, and Herpes Simplex)" International Journal of Molecular Sciences 23, no. 7: 3550. https://doi.org/10.3390/ijms23073550