
Alteration of Oral and Perioral Soft Tissue in Mice following Incisor Tooth Extraction

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
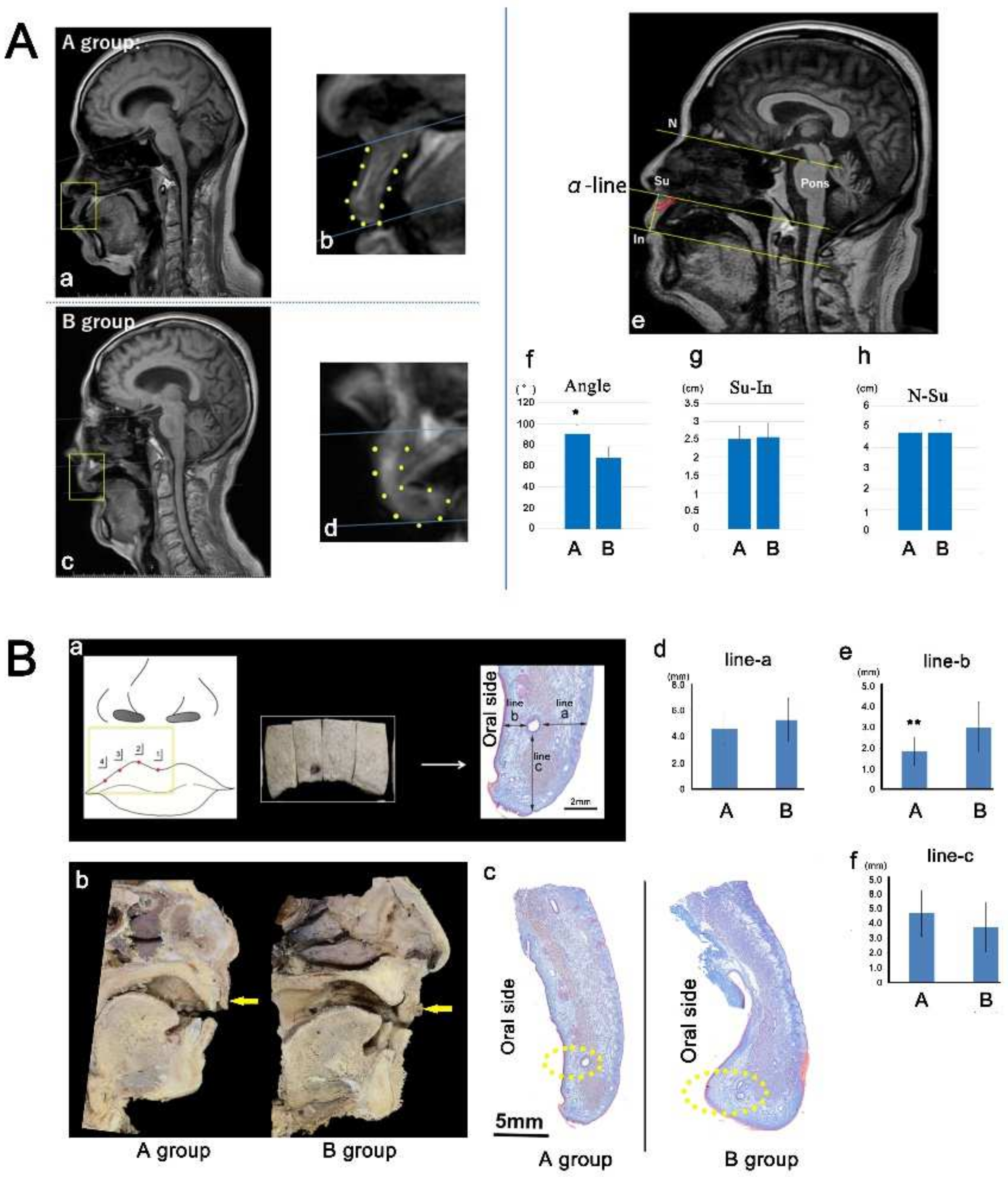
2.1. Morphology of the Upper Lip in Humans
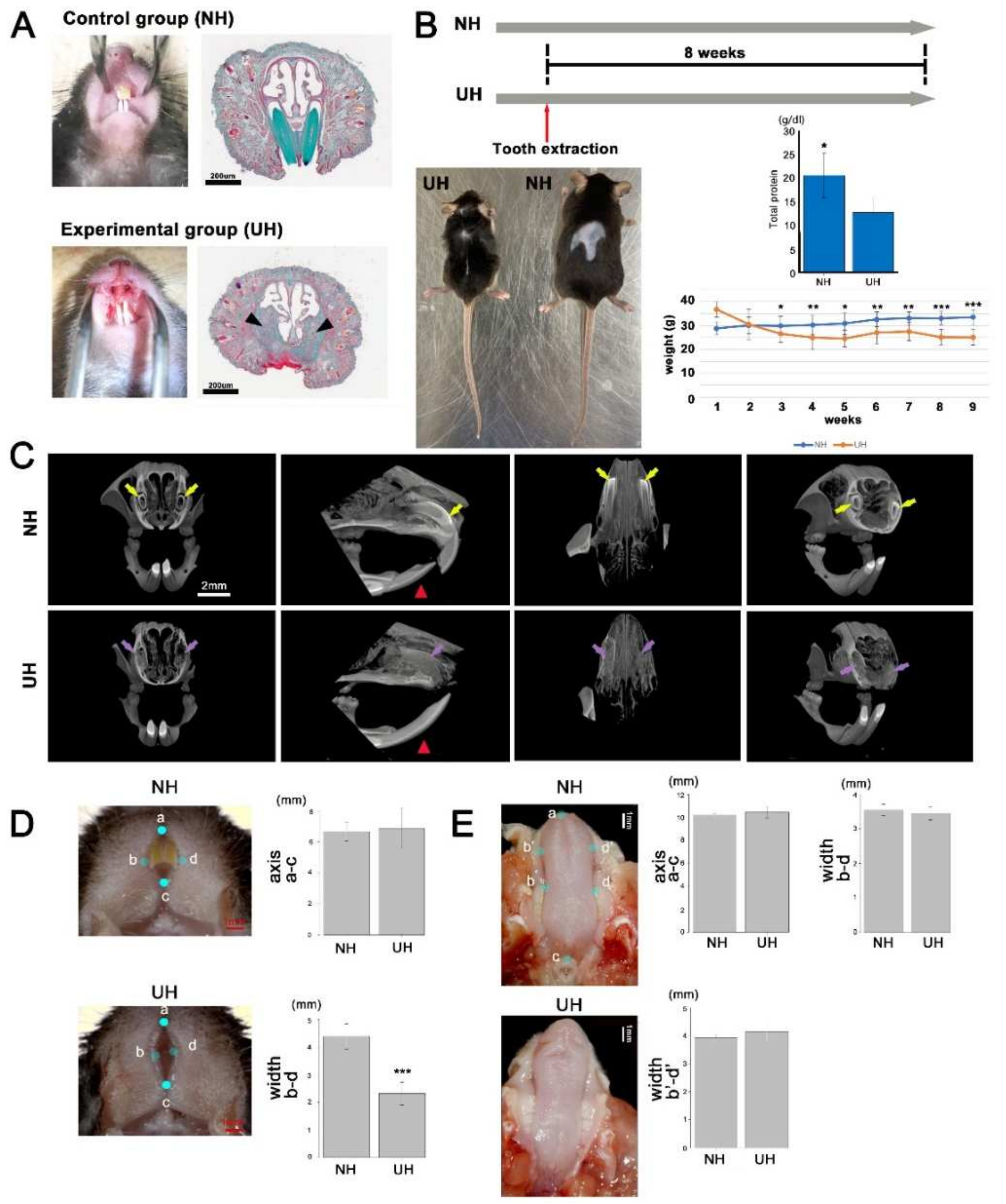
2.2. Incisor-Tooth-Deficient Mice Were Obtained by Extracting an Incisor Tooth
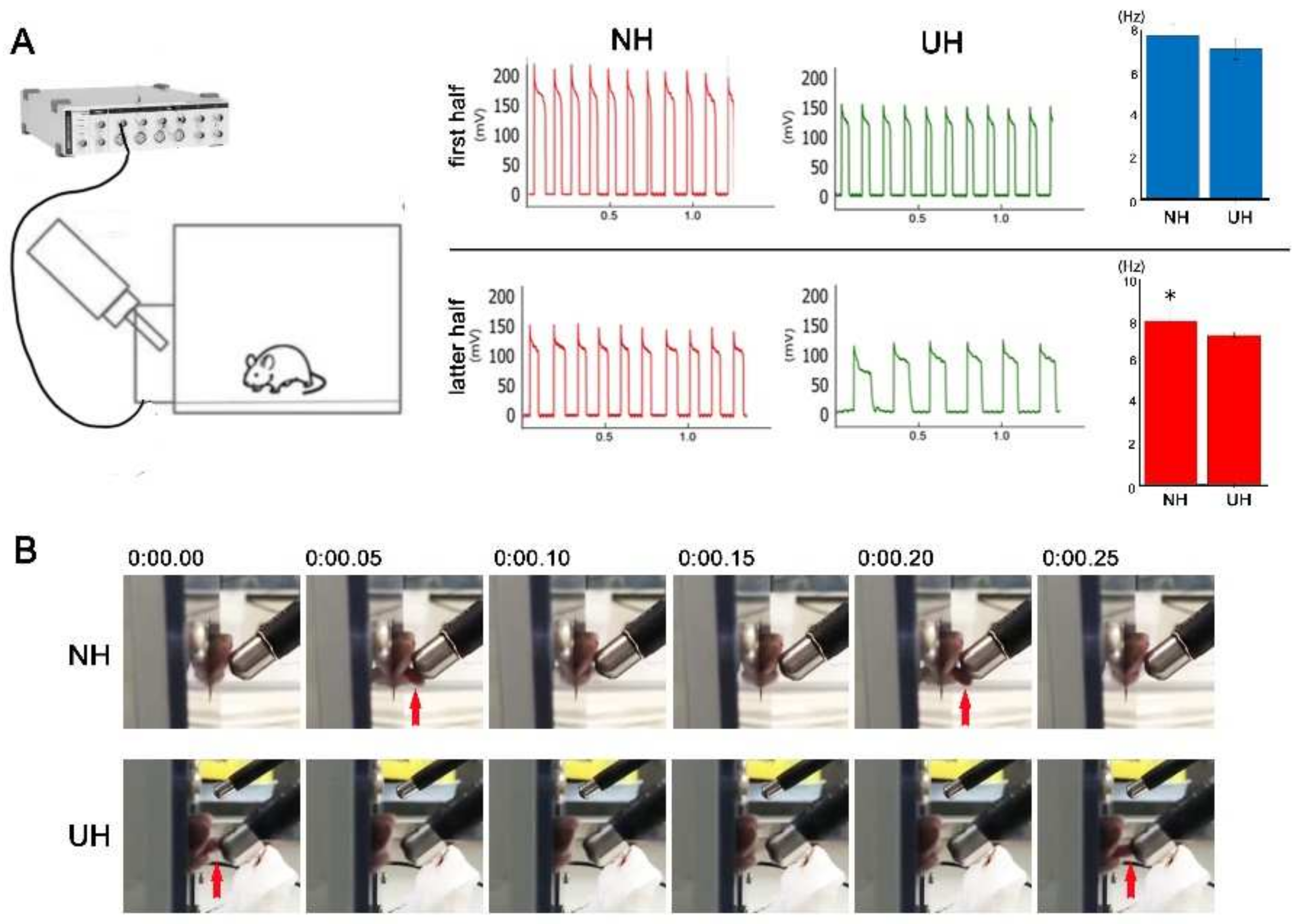
2.3. Measuring and Recording Lapping Behavior in Mice
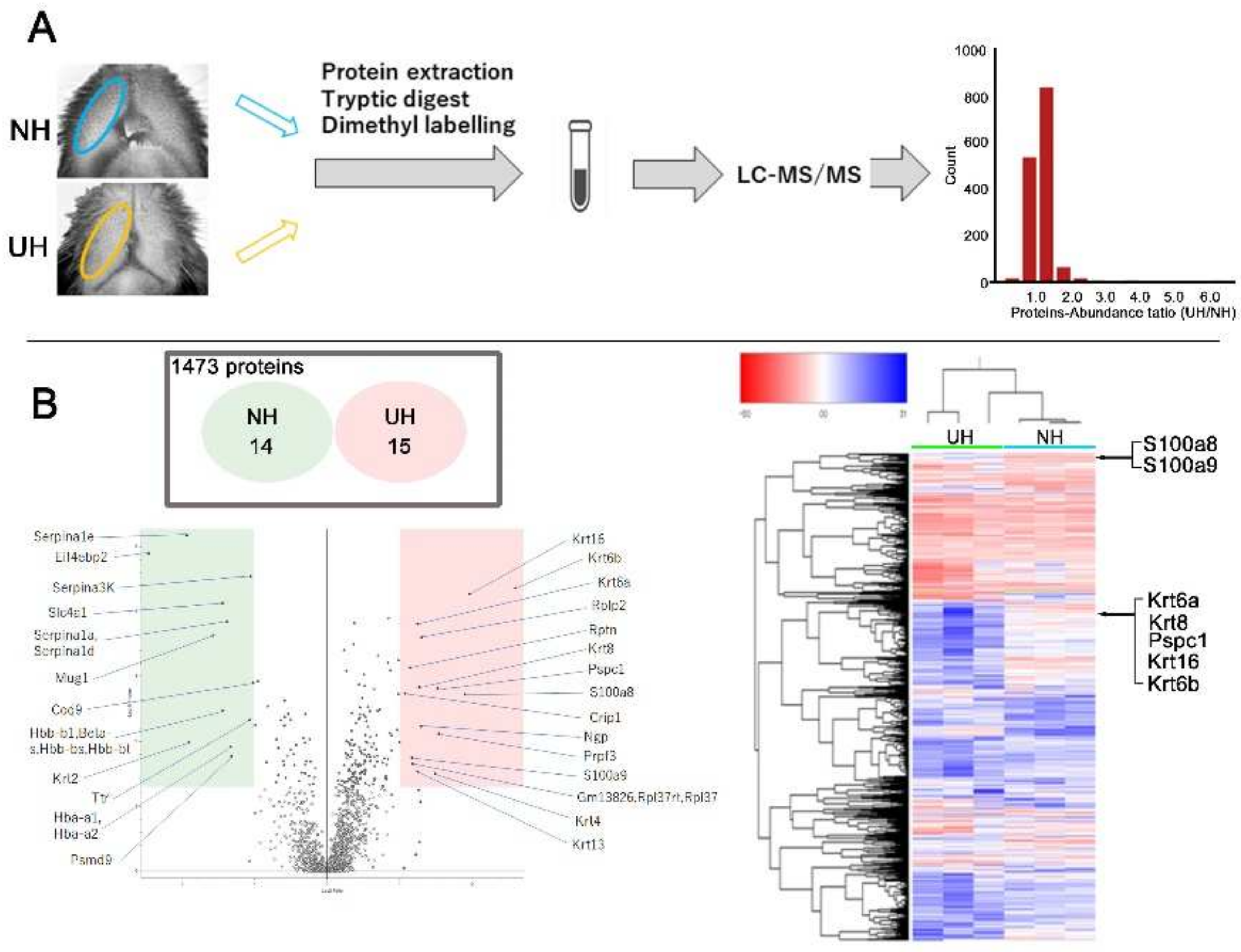
2.4. Quantitative Proteome Analysis of Differences in Expression between NH and UH Mice’s Lips
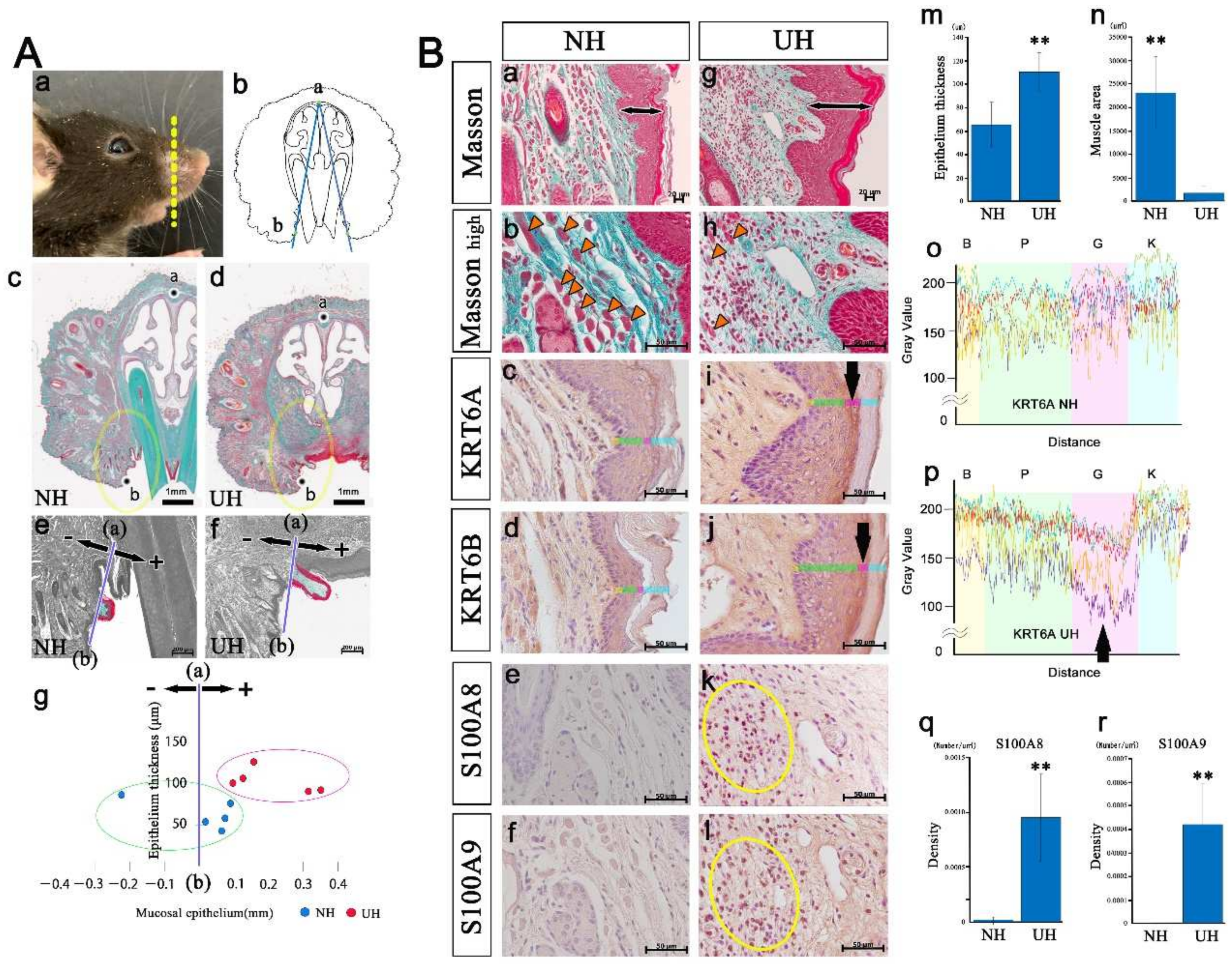
2.5. An Upper Lip Comparison between Normal and Incisor-Tooth-Deficient Mice
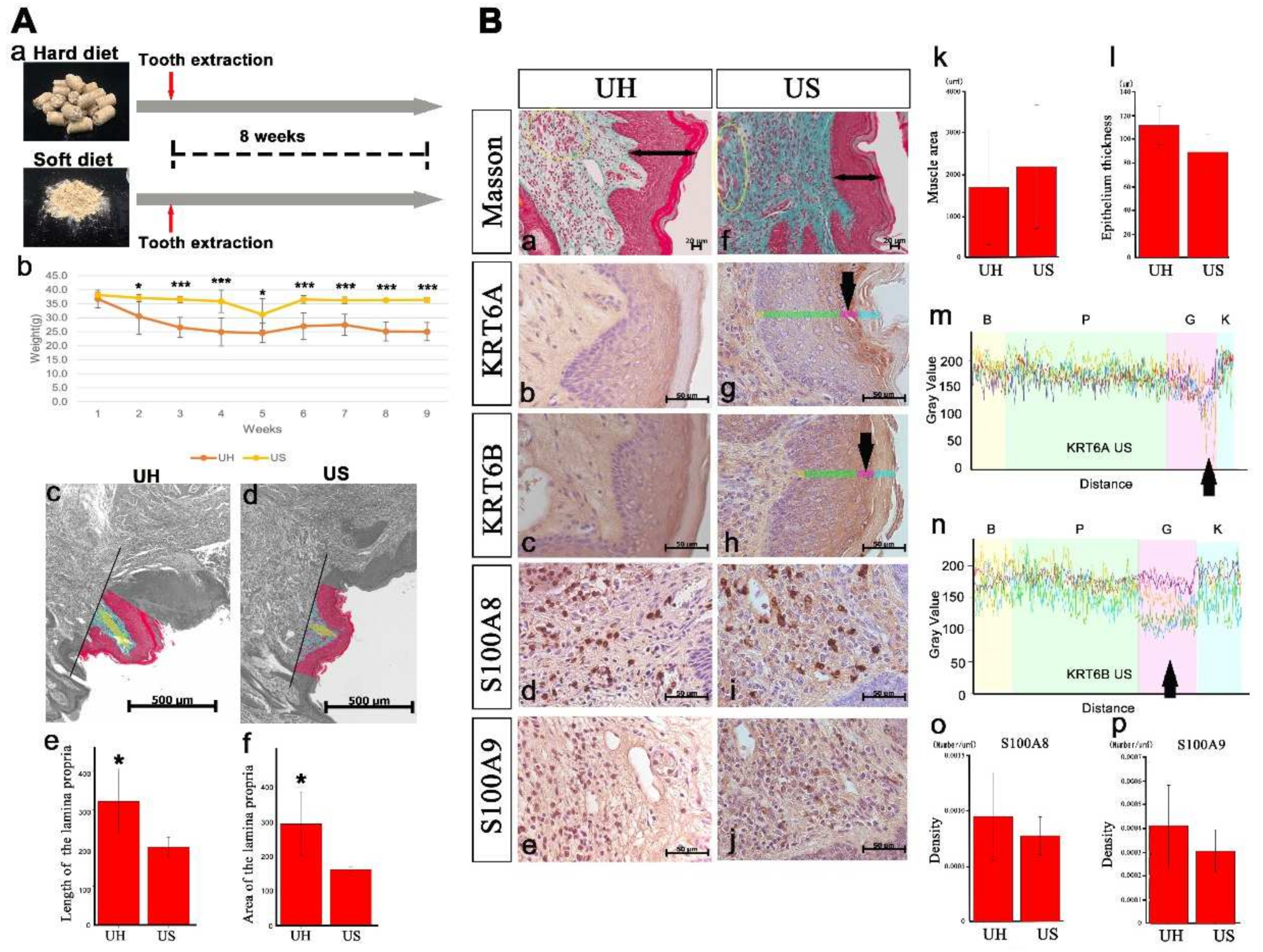
2.6. An Upper Lip Comparison between Hard-Diet and Soft-Diet Feeding in Incisor-Tooth-Deficient Mice
3. Discussion
4. Materials and Methods
4.1. Magnetic Resonance Imaging (MRI) in Humans
4.2. Gross and Histological Anatomy in Humans
4.3. Generation of Incisor-Tooth-Deficient Mice
4.4. Histological Analysis
4.5. Micro-CT Analysis
4.6. Lickometer and Recording of Licking Behavior
4.7. Quantitative Proteome Analysis
4.8. Immunohistochemical Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adegboye, A.R.; Twetman, S.; Christensen, L.B.; Heitmann, B.L. Intake of dairy calcium and tooth loss among adult Danish men and women. Nutrition 2012, 28, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kisely, S.; Quek, L.H.; Pais, J.; Lalloo, R.; Johnson, N.W.; Lawrence, D. Advanced dental disease in people with severe mental illness: Systematic review and meta-analysis. Br. J. Psychiatry 2011, 199, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Leng, W.D.; Zeng, X.T.; Kwong, J.S.; Hua, X.P. Periodontal disease and risk of coronary heart disease: An updated meta-analysis of prospective cohort studies. Int. J. Cardiol. 2015, 201, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Watt, R.G.; Tsakos, G.; de Oliveira, C.; Hamer, M. Tooth loss and cardiovascular disease mortality risk-results from the Scottish Health Survey. PLoS ONE 2012, 7, e30797. [Google Scholar] [CrossRef] [PubMed]
- Desvarieux, M.; Demmer, R.T.; Rundek, T.; Boden-Albala, B.; Jacobs, D.R.; Papapanou, P.N.; Sacco, R.L.; Oral Infections and Vascular Disease Epidemiology Study (INVEST). Relationship between periodontal disease, tooth loss, and carotid artery plaque: The Oral Infections and Vascular Disease Epidemiology Study (INVEST). Stroke 2003, 34, 2120–2125. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.T.; Luo, W.; Huang, W.; Wang, Q.; Guo, Y.; Leng, W.D. Tooth loss and head and neck cancer: A meta-analysis of observational studies. PLoS ONE 2013, 8, e79074. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.L.; Zeng, X.T.; Luo, Z.X.; Duan, X.L.; Qin, J.; Leng, W.D. Tooth loss is associated with increased risk of esophageal cancer: Evidence from a meta-analysis with dose–response analysis. Sci. Rep. 2016, 6, 18900. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Kubota, M.; Takahashi, T.; Nakasato, A.; Nomura, T.; Furuya, J.; Kondo, H. Effects of tooth loss on brain structure: A voxel-based morphometry study. J. Prosthodont. Res. 2018, 62, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Moreno, P.; Lopez-Chaichio, L.; Padial-Molina, M.; Avila-Ortiz, G.; O’Valle, F.; Ravida, A.; Catena, A. The impact of tooth loss on cognitive function. Clin. Oral Investig. 2021, 1–8. [Google Scholar] [CrossRef]
- Cho, H.A.; Shin, H. Effects of tooth loss on functional impairment in elderly: A secondary data analysis of elderly cohort in the South Korea. Int. J. Dent. Hyg. 2021, 10. [Google Scholar] [CrossRef]
- Chen, J.; Ren, C.J.; Wu, L.; Xia, L.Y.; Shao, J.; Leng, W.D.; Zeng, X.T. Tooth loss is associated with increased risk of dementia and with a dose–response relationship. Front. Aging Neurosci. 2018, 10, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffcoat, M.K. Bone loss in the oral cavity. J. Bone Miner. Res. 1993, 8 (Suppl. 2), S467–S473. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Chen, H.K.; Hidetomo, H.; Watanabe, A.; Sakiyama, K.; Kim, H.J.; Murakami, G.; Rodríguez-Vázquez, J.F.; Abe, S. Superior labial artery and vein anastomosis configuration to be considered in lip augmentation. Ann. Anat.-Anat. Anz. 2022, 239, 151808. [Google Scholar] [CrossRef] [PubMed]
- Waldman, B.H. Change in lip contour with maxillary incisor retraction. Angle Orthod. 1982, 52, 129–134. [Google Scholar]
- Hayashida, H.; Ioi, H.; Nakata, S.; Takahashi, I.; Counts, A.L. Effects of retraction of anterior teeth and initial soft tissue variables on lip changes in Japanese adults. Eur. J. Orthod. 2011, 33, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Iblher, N.; Kloepper, J.; Penna, V.; Bartholomae, J.P.; Stark, G.B. Changes in the aging upper lip—A photomorphometric and MRI-based study (on a quest to find the right rejuvenation approach). J. Plast. Reconstr. Aesthetic Surg. 2008, 61, 1170–1176. [Google Scholar] [CrossRef]
- Zitzmann, N.U.; Marinello, C.P. Treatment plan for restoring the edentulous maxilla with implant-supported restorations: Removable overdenture versus fixed partial denture design. J. Prosthet. Dent. 1999, 82, 188–196. [Google Scholar] [CrossRef]
- Hayar, A.; Bryant, J.L.; Boughter, J.D.; Heck, D.H. A low-cost solution to measure mouse licking in an electrophysiological setup with a standard analog-to-digital converter. J. Neurosci. Methods 2006, 153, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef]
- Yamamoto, M.; Takada, H.; Ishizuka, S.; Kitamura, K.; Jeong, J.; Sato, M.; Hinata, N.; Abe, S. Morphological association between the muscles and bones in the craniofacial region. PLoS ONE 2020, 15, e0227301. [Google Scholar] [CrossRef]
- Ishizuka, S.; Yamamoto, M.; Hirouchi, H.; Yotsuya, M.; Ohkubo, M.; Sato, M.; Abe, S. Muscle-bone relationship in temporomandibular joint disorders after partial discectomy. J. Oral Biosci. 2021, 63, 436–443. [Google Scholar] [CrossRef]
- Abe, S.; Yamamoto, M. Factors involved in morphogenesis in the muscle-tendon-bone complex. Int. J. Mol. Sci. 2021, 22, 6365. [Google Scholar] [CrossRef] [PubMed]
- Brakefield, P.M. Evo-devo and accounting for Darwin’s endless forms. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.H. Genetic assimilation of the bithorax phenotype. Evolution 1956, 10, 1–13. [Google Scholar] [CrossRef]
- Noble, D.D. Conrad Waddington and the origin of epigenetics. J. Exp. Biol. 2015, 218, 816–818. [Google Scholar] [CrossRef] [Green Version]
- Glastad, K.M.; Hunt, B.G.; Goodisman, M.A.D. Epigenetics in insects: Genome regulation and the generation of phenotypic diversity. Annu. Rev. Entomol. 2019, 64, 185–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahony, D.; Karunaratne, S.; Cam, G.; Rothnagel, J.A. Analysis of mouse keratin 6A regulatory sequences in transgenic mice reveals constitutive, tissue-specific expression by a keratin 6A minigene. J. Investig. Dermatol. 2000, 115, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Coulombe, P.A.; Miyachi, Y. Using transgenic models to study the pathogenesis of keratin-based inherited skin diseases. J. Dermatol. Sci. 1999, 21, 73–95. [Google Scholar] [CrossRef]
- Wojcik, S.M.; Bundman, D.S.; Roop, D.R. Delayed wound healing in keratin 6A knockout mice. Mol. Cell. Biol. 2000, 20, 5248–5255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, P.; Colucci-Guyon, E.; Takahashi, K.; Gu, C.; Babinet, C.; Coulombe, P.A. Introducing a null mutation in the mouse K6alpha and K6beta genes reveals their essential structural role in the oral mucosa. J. Cell. Biol. 2000, 150, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Vilmann, H.; Kirkeby, S.; Kronborg, D. Histomorphometrical analysis of the influence of soft diet on masticatory muscle development in the muscular dystrophic mouse. Arch. Oral Biol. 1990, 35, 37–42. [Google Scholar] [CrossRef]
- Fujishita, A.; Koga, Y.; Utsumi, D.; Nakamura, A.; Yoshimi, T.; Yoshida, N. Effects of feeding a soft diet and subsequent rehabilitation on the development of the masticatory function. J. Oral Rehabil. 2015, 42, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Honda, A.; Abe, S.; Hiroki, E.; Honda, H.; Iwanuma, O.; Yanagisawa, N.; Ide, Y. Activation of caspase 3,9,12, and Bax in masseter musle of mdx mice during necrosis. J. Muscle Res. Cell Motil. 2007, 28, 243–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, S.; Soejima, M.; Iwanuma, O.; Saka, H.; Matsunaga, S.; Sakiyama, K.; Ide, Y. Expression fo myostatin and follistatin in mdx mice, an animal model for muscular dystrophy. Zool. Sci. 2009, 26, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, M.; Niijima-Yaoita, F.; Yoneda, H.; Chiba, K.; Tsuchiya, S.; Hagiwara, Y.; Sasaki, K.; Sugawara, S.; Endo, Y.; Tan-No, K.; et al. Long-term feeding on powdered food causes hyperglycemia and signs of systemic illness in mice. Life Sci. 2014, 103, 8–14. [Google Scholar] [CrossRef]
- Abe, S.; Maejima, M.; Watanabe, H.; Shibahara, T.; Agematsu, H.; Doi, T.; Sakiyama, K.; Usami, A.; Gojyo, K.; Hashimoto, M.; et al. Muscle-fiber characteristics in adult mouse-tongue muscles. Anat. Sci. Int. 2002, 77, 145–148. [Google Scholar] [CrossRef]
- Usami, A.; Abe, S.; Ide, Y. Myosin heavy chain isoforms of the murine masseter muscle during pre- and post-natal development. Anat. Histol. Embryol. 2003, 32, 244–248. [Google Scholar] [CrossRef]
- Kurokawa, K.; Abe, S.; Sakiyama, K.; Takeda, T.; Ide, Y.; Ishigami, K. Effects of stretching stimulation with different rates on the expression of MyHC mRNA in mouse cultured myoblasts. Biomed Res. 2007, 28, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Gojyo, K.; Abe, S.; Ide, Y. Characteristics of myofibres in the masseter muscle of mice during postnatal growth period. Anat. Histol. Embryoll. 2002, 31, 105–112. [Google Scholar] [CrossRef]
- Okubo, K.; Abe, S.; Usami, A.; Agematsu, H.; Nakamura, H.; Hashimoto, M.; Ide, Y. Changes in muscle-fiber properties of the murine digastric muscle before and after weaning. Zool. Sci. 2006, 23, 1079–1084. [Google Scholar] [CrossRef] [Green Version]
- Almazrooa, S.A.; Woo, S.B.; Mawardi, H.; Treister, N. Characterization and management of exfoliative cheilitis: A single-center experience. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2013, 116, e485–e489. [Google Scholar] [CrossRef]
- Bork, K. Diseases of the lips and mouth. In Braun-Falco’s Dermatology, 3rd ed.; Burgdorf, W.H.C., Plewig, G., Wolf, H.H., Landthaler, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1081–1108. [Google Scholar]
- Collet, E.; Jeudy, G.; Dalac, S. Cheilitis, perioral dermatitis and contact allergy. Eur. J. Dermatol. 2013, 23, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Lugović-Mihić, L.; Pilipović, K.; Crnarić, I.; Šitum, M.; Duvančić, T. Differential Diagnosis of Cheilitis-How to Classify Cheilitis? Acta Clin. Croat. 2018, 57, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Litaiem, N.; Ben Slimane, M.; Bacha, T.; Rammeh, S.; Zeglaoui, F. Cheilitis with hemorrhagic crusts of the vermilion lips. Int. J. Dermatol. 2020, 59, e234–e236. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K. Reference lines on diagnostic imagings of the head. Part 2: Reference lines on MRI. Nihon Igaku Hoshasen Gakkai Zasshi. Nippon. Acta Radiol. 1989, 49, 264–269. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takagi, T.; Yamamoto, M.; Sugano, A.; Kanehira, C.; Kitamura, K.; Katayama, M.; Sakai, K.; Sato, M.; Abe, S. Alteration of Oral and Perioral Soft Tissue in Mice following Incisor Tooth Extraction. Int. J. Mol. Sci. 2022, 23, 2987. https://doi.org/10.3390/ijms23062987
Takagi T, Yamamoto M, Sugano A, Kanehira C, Kitamura K, Katayama M, Sakai K, Sato M, Abe S. Alteration of Oral and Perioral Soft Tissue in Mice following Incisor Tooth Extraction. International Journal of Molecular Sciences. 2022; 23(6):2987. https://doi.org/10.3390/ijms23062987
Chicago/Turabian StyleTakagi, Takahiro, Masahito Yamamoto, Aki Sugano, Chiemi Kanehira, Kei Kitamura, Masateru Katayama, Katsuhiko Sakai, Masaki Sato, and Shinichi Abe. 2022. "Alteration of Oral and Perioral Soft Tissue in Mice following Incisor Tooth Extraction" International Journal of Molecular Sciences 23, no. 6: 2987. https://doi.org/10.3390/ijms23062987