
Modeling Trophoblast Cell-Guided Uterine Spiral Artery Transformation in the Rat


Abstract
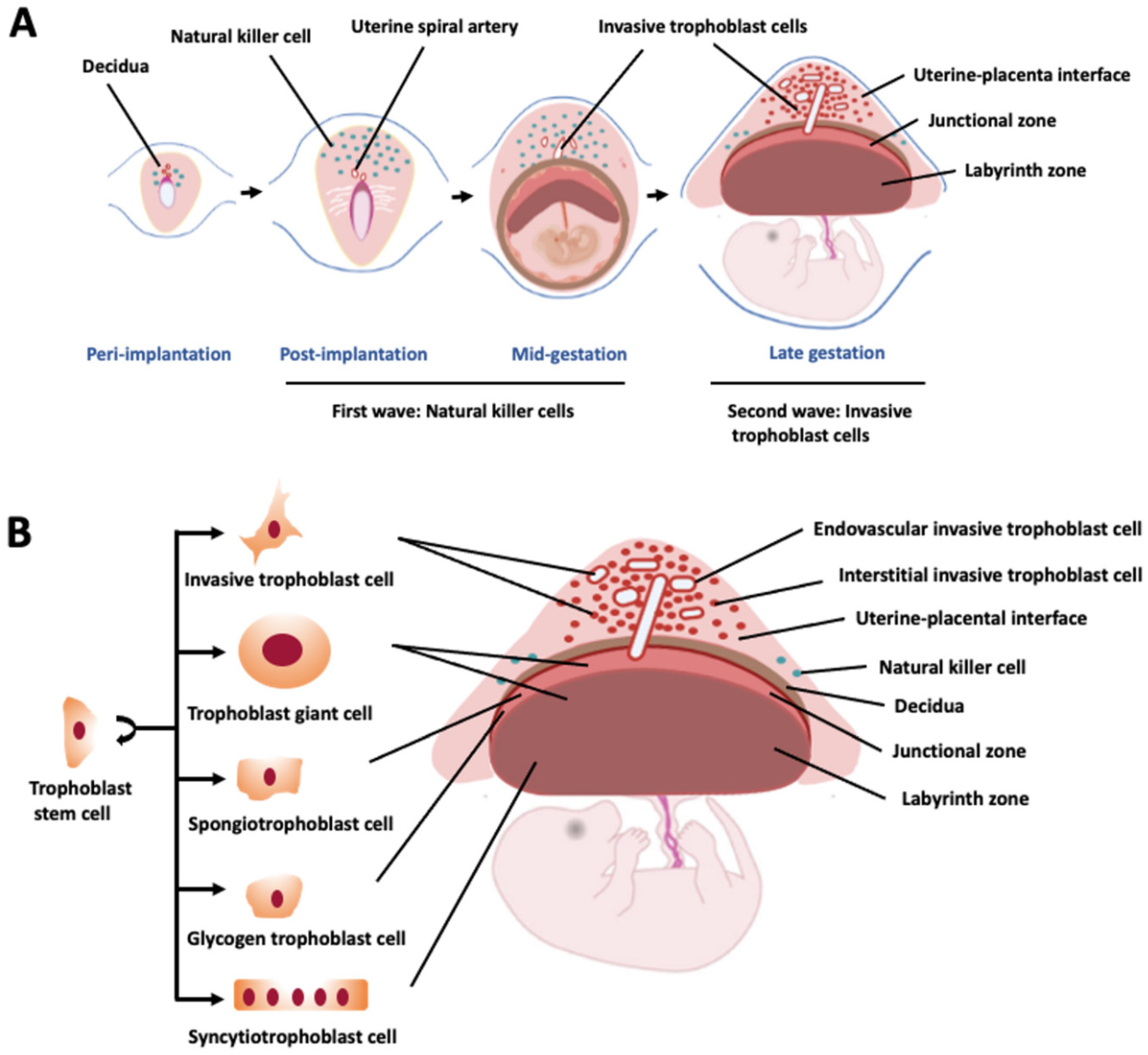
:1. Introduction
2. Organization of the Rat Placentation Site
3. Life Cycle of Invasive Trophoblast Cells
4. Genetics of Trophoblast Cell Invasion
5. Experimental Manipulation of Trophoblast Cell Invasion and Uterine Spiral Artery Remodeling
5.1. Oxygen Tension
5.2. Immune Cells
5.3. Drug, Toxicant, and Miscellaneous Exposures
5.4. Disease States
5.4.1. Diabetes
5.4.2. Hypertension/Preeclampsia
5.4.3. Malnutrition/Obesity/Hyperthermia
5.4.4. Overview
6. A Path to Conserved Mechanisms Controlling Invasive Trophoblast Cell-Guided Uterine Transformation
7. Final Thoughts
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burton, G.J.; Fowden, A.L.; Thornburg, K.L. Placental origins of chronic disease. Physiol. Rev. 2016, 96, 1509–1565. [Google Scholar] [CrossRef] [PubMed]
- Maltepe, E.; Fisher, S.J. Placenta: The forgotten organ. Annu. Rev. Cell. Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.M.; Green, J.A.; Schulz, L.C. The evolution of the placenta. Reproduction 2016, 152, R179–R189. [Google Scholar] [CrossRef] [Green Version]
- Pijnenborg, R.; Robertson, W.B.; Brosens, I.; Dixon, G. Trophoblast invasion and the establishment of haemochorial placentation in man and laboratory animals. Placenta 1981, 2, 71–91. [Google Scholar] [CrossRef]
- Soares, M.J.; Varberg, K.M.; Iqbal, K. Hemochorial placentation: Development, function, and adaptations. Biol. Reprod. 2018, 99, 196–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.J.B.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [Green Version]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The uterine spiral arteries in human pregnancy: Facts and controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef]
- Pollheimer, J.; Vondra, S.; Baltayeva, J.; Beristain, A.G.; Knöfler, M. Regulation of placental extravillous trophoblasts by the maternal uterine environment. Front. Immunol. 2018, 9, 2597. [Google Scholar] [CrossRef]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular trophoblast invasion: Implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol. Reprod. 2003, 69, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Brosens, I.; Puttemans, P.; Benagiano, G. Placental bed research: I. The placental bed: From spiral arteries remodeling to the great obstetrical syndromes. Am. J. Obstet. Gynecol. 2019, 221, 437–456. [Google Scholar] [CrossRef]
- Wooding, P.; Burton, G. Comparative Placentation; Springer: Heidelberg, Germany, 2008. [Google Scholar]
- Pijnenborg, R.; Vercruysse, L. Animal models of deep trophoblast invasion. In Placental Bed Disorders; Pijnenborg, R., Brosens, I., Romero, R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 127–139. [Google Scholar]
- Soares, M.J.; Chakraborty, D.; Rumi, M.A.K.; Konno, T.; Renaud, S.J. Rat placentation: An experimental model for investigating the hemochorial maternal-fetal interface. Placenta 2012, 33, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef] [PubMed]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef] [PubMed]
- Turco, M.Y.; Moffett, A. Development of the human placenta. Development 2019, 146, dev163428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridgman, J. A morphological study of the development of the placenta of the rat; an histological and cytological study of the development of the chorioallantoic placenta of the white rat. J. Morphol. 1948, 83, 195–223. [Google Scholar] [CrossRef]
- Enders, A.C. A comparative study of the fine structure of the trophoblast in several hemochorial placentas. Am. J. Anat. 1965, 116, 29–67. [Google Scholar] [CrossRef]
- Jollie, W.P. Fine structural changes in the junctional zone of the rat placenta with increasing gestational age. J. Ultrastruct. Res. 1965, 12, 420–438. [Google Scholar] [CrossRef]
- Davies, J.; Glasser, S.R. Histological and fine structural observations on the placenta of the rat. Acta Anat. 1968, 69, 542–608. [Google Scholar] [CrossRef]
- Rossant, J. Stem cells from the mammalian blastocyst. Stem Cells 2001, 19, 477–482. [Google Scholar] [CrossRef]
- Gardner, R.L.; Beddington, R.S. Multi-lineage ‘stem’ cells in the mammalian embryo. J. Cell Sci. Suppl. 1988, 10, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Simmons, D.G.; Cross, J.C. Determinants of trophoblast lineage and cell subtype specification in the mouse placenta. Dev. Biol. 2005, 284, 12–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walentin, K.; Hinze, C.; Schmidt-Ott, K.M. The basal chorionic trophoblast cell layer: An emerging coordinator of placenta development. Bioessays 2016, 38, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.J.; Chapman, B.M.; Rasmussen, C.A.; Dai, G.; Kamei, T.; Orwig, K.E. Differentiation of trophoblast endocrine cells. Placenta 1996, 17, 277–289. [Google Scholar] [CrossRef]
- Cross, J.C.; Nakano, H.; Natale, D.R.; Simmons, D.G.; Watson, E.D. Branching morphogenesis during development of placental villi. Differentiation 2006, 74, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Knipp, G.T.; Audus, K.L.; Soares, M.J. Nutrient transport across the placenta. Adv. Drug Deliv. Rev. 1999, 38, 41–58. [Google Scholar] [CrossRef]
- Watson, E.D.; Cross, J.C. Development of structures and transport functions in the mouse placenta. Physiology 2005, 20, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Peel, S.; Bulmer, D. Proliferation and differentiation of trophoblast in the establishment of the rat chorio-allantoic placenta. J. Anat. 1977, 124 Pt 3, 675–687. [Google Scholar]
- John, R.M. Epigenetic regulation of placental endocrine lineages and complications of pregnancy. Biochem. Soc. Trans. 2013, 41, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Tunster, S.J.; Watson, E.D.; Fowden, A.L.; Burton, G.J. Placental glycogen stores and fetal growth: Insights from genetic mouse models. Reproduction 2020, 159, R213–R235. [Google Scholar] [CrossRef]
- Varberg, K.M.; Soares, M.J. Paradigms for investigating invasive trophoblast cell development and contributions to uterine spiral artery remodeling. Placenta 2021, 113, 48–56. [Google Scholar] [CrossRef]
- Selye, H.; McKeown, T. Studies on the physiology of the maternal placenta in the rat. Proc. R. Soc. Lond. Biol. 1935, 119, 1–31. [Google Scholar]
- Ain, R.; Canham, L.N.; Soares, M.J. Gestation stage-dependent intrauterine trophoblast cell invasion in the rat and mouse: Novel endocrine phenotype and regulation. Dev. Biol. 2003, 260, 176–190. [Google Scholar] [CrossRef] [Green Version]
- Caluwaerts, S.; Vercruysse, L.; Luyten, C.; Pijnenborg, R. Endovascular trophoblast invasion and associated structural changes in uterine spiral arteries of the pregnant rat. Placenta 2005, 26, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, L.; Caluwaerts, S.; Luyten, C.; Pijnenborg, R. Interstitial trophoblast invasion in the decidua and mesometrial triangle during the last third of pregnancy in the rat. Placenta 2006, 27, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Cross, J.C. Development and function of trophoblast giant cells in the rodent placenta. Int. J. Dev. Biol. 2010, 54, 341–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zybina, T.G.; Zybina, E.V. Role of cell cycling and polyploidy in placental trophoblast of different mammalian species. Reprod. Domest. Anim. 2020, 55, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Ain, R.; Konno, T.; Canham, L.N.; Soares, M.J. Phenotypic analysis of the rat placenta. Methods Mol. Med. 2006, 121, 295–313. [Google Scholar]
- Furukawa, S.; Tsuji, N.; Sugiyama, A. Morphology and physiology of rat placenta for toxicological evaluation. J. Toxicol. Pathol. 2019, 32, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Campbell, W.J.; Deb, S.; Kwok, S.C.; Joslin, J.A.; Soares, M.J. Differential expression of placental lactogen-II and prolactin-like protein-A in the rat chorioallantoic placenta. Endocrinology 1989, 125, 1565–1574. [Google Scholar] [CrossRef]
- Correia-da-Silva, G.; Bell, S.C.; Pringle, J.H.; Teixeira, N. Expression of mRNA encoding insulin-like growth factors I and II by uterine tissues and placenta during pregnancy in the rat. Mol. Reprod. Dev. 1999, 53, 294–305. [Google Scholar] [CrossRef]
- Toft, D.J.; Linzer, D.I. Identification of three prolactin-related hormones as markers of invasive trophoblasts in the rat. Biol. Reprod. 2000, 63, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zybina, T.G.; Stein, G.I.; Zybina, E.V. Endopolyploid and proliferating trophoblast cells express different patterns of intracellular cytokeratin and glycogen localization in the rat placenta. Cell Biol. Int. 2011, 35, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Wiemers, D.O.; Ain, R.; Ohboshi, S.; Soares, M.J. Migratory trophoblast cells express a newly identified member of the prolactin gene family. J. Endocrinol. 2003, 179, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosario, G.X.; Konno, T.; Soares, M.J. Maternal hypoxia activates endovascular trophoblast cell invasion. Dev. Biol. 2008, 314, 362–375. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, D.; Rumi, M.A.; Konno, T.; Soares, M.J. Natural killer cells direct hemochorial placentation by regulating hypoxia-inducible factor dependent trophoblast lineage decisions. Proc. Natl. Acad. Sci. USA 2011, 108, 16295–16300. [Google Scholar] [CrossRef] [Green Version]
- Muto, M.; Chakraborty, D.; Varberg, K.M.; Moreno-Irusta, A.; Iqbal, K.; Scott, R.L.; McNally, R.P.; Choudhury, R.H.; Aplin, J.D.; Okae, H.; et al. Intersection of regulatory pathways controlling hemostasis and hemochorial placentation. Proc. Natl. Acad. Sci. USA 2021, 118, e2111267118. [Google Scholar] [CrossRef]
- Nandy, D.; Das, S.; Islam, S.; Ain, R. Molecular regulation of vascular smooth muscle cell phenotype switching by trophoblast cells at the maternal-fetal interface. Placenta 2020, 93, 64–73. [Google Scholar] [CrossRef]
- Rosario, G.X.; Ain, R.; Konno, T.; Soares, M.J. Intrauterine fate of invasive trophoblast cells. Placenta 2009, 30, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Konno, T.; Rempel, L.A.; Arroyo, J.A.; Soares, M.J. Pregnancy in the brown Norway rat: A model for investigating the genetics of placentation. Biol. Reprod. 2007, 76, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Konno, T.; Graham, A.R.; Rempel, L.A.; Ho-Chen, J.K.; Alam, S.M.; Bu, P.; Rumi, M.A.; Soares, M.J. Subfertility linked to combined luteal insufficiency and uterine progesterone resistance. Endocrinology 2010, 151, 4537–4550. [Google Scholar] [CrossRef] [Green Version]
- Konno, T.; Rempel, L.A.; Rumi, M.A.; Graham, A.R.; Asanoma, K.; Renaud, S.J.; Soares, M.J. Chromosome-substituted rat strains provide insights into the genetics of placentation. Physiol. Genom. 2011, 43, 930–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, R.; Yellon, S.M.; Longo, L.D.; Mata-Greenwood, E. Placental gene expression in a rat ‘model’ of placental insufficiency. Placenta 2010, 31, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Buresova, M.; Zidek, V.; Musilova, A.; Simakova, M.; Fucikova, A.; Bila, V.; Kren, V.; Kazdova, L.; Di Nicolantonio, R.; Pravenec, M. Genetic relationship between placental and fetal weights and markers of the metabolic syndrome in rat recombinant inbred strains. Physiol. Genom. 2006, 26, 226–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, K.P.; Rabaglino, M.B.; Post Uiterweer, E.D. Emerging role for dysregulated decidualization in the genesis of preeclampsia. Placenta 2017, 60, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Gomez, T.; Dominguez, F.; Quiñonero, A.; Diaz-Gimeno, P.; Kapidzic, M.; Gormley, M.; Ona, K.; Padilla-Iserte, P.; McMaster, M.; Genbacev, O.; et al. Defective decidualization during and after severe preeclampsia reveals a possible maternal contribution to the etiology. Proc. Natl. Acad. Sci. USA 2017, 114, E8468–E8477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Gómez, T.; Castillo-Marco, N.; Cordero, T.; Simón, C. Decidualization resistance in the origin of preeclampsia. Am. J. Obstet. Gynecol. 2020, 226, S886–S894. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J. Oxygen, the Janus gas; its effects on human placental development and function. J. Anat. 2009, 215, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Fryer, B.H.; Simon, M.C. Hypoxia, HIF and the placenta. Cell Cycle 2006, 5, 495–498. [Google Scholar] [CrossRef]
- Soares, M.J.; Iqbal, K.; Kozai, K. Hypoxia and placental development. Birth Defects Res. 2017, 109, 1309–1329. [Google Scholar] [CrossRef]
- Chang, C.W.; Wakeland, A.K.; Parast, M.M. Trophoblast lineage specification, differentiation and their regulation by oxygen tension. J. Endocrinol. 2018, 236, R43–R56. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, A.M.; Camm, E.J.; Sferruzzi-Perri, A.N.; Ashmore, T.J.; Yung, H.W.; Cindrova-Davies, T.; Spiroski, A.M.; Sutherland, M.R.; Logan, A.; Austin-Williams, S.; et al. Placental adaptation to early-onset hypoxic pregnancy and mitochondria-targeted antioxidant therapy in a rodent model. Am. J. Pathol. 2018, 188, 2704–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, D.; Cui, W.; Rosario, G.X.; Scott, R.L.; Dhakal, P.; Renaud, S.J.; Tachibana, M.; Rumi, M.A.; Mason, C.W.; Krieg, A.J.; et al. HIF-KDM3A-MMP12 regulatory circuit ensures trophoblast plasticity and placental adaptations to hypoxia. Proc. Natl. Acad. Sci. USA 2016, 113, E7212–E7221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiden, U.; Eyth, C.P.; Majali-Martinez, A.; Desoye, G.; Tam-Amersdorfer, C.; Huppertz, B.; Ghaffari Tabrizi-Wizsy, N. Expression of matrix metallopeptidase 12 is highly specific for non-proliferating invasive trophoblasts in the first trimester and temporally regulated by oxygen-dependent mechanisms including HIF-1A. Histochem. Cell Biol. 2018, 149, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, L.K.; Keogh, R.J.; Wareing, M.; Baker, P.N.; Cartwright, J.E.; Whitley, G.S.; Aplin, J.D. BeWo cells stimulate smooth muscle cell apoptosis and elastin breakdown in a model of spiral artery transformation. Hum. Reprod. 2007, 22, 2834–2841. [Google Scholar] [CrossRef] [Green Version]
- Harris, L.K.; Smith, S.D.; Keogh, R.J.; Jones, R.L.; Baker, P.N.; Knöfler, M.; Cartwright, J.E.; Whitley, G.S.; Aplin, J.D. Trophoblast- and vascular smooth muscle cell-derived MMP-12 mediates elastolysis during uterine spiral artery remodeling. Am. J. Pathol. 2010, 177, 2103–2115. [Google Scholar] [CrossRef]
- Asanoma, K.; Rumi, M.A.; Kent, L.N.; Chakraborty, D.; Renaud, S.J.; Wake, N.; Lee, D.S.; Kubota, K.; Soares, M.J. FGF4-dependent stem cells derived from rat blastocysts differentiate along the trophoblast lineage. Dev. Biol. 2011, 351, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Wakeland, A.K.; Soncin, F.; Moretto-Zita, M.; Chang, C.W.; Horii, M.; Pizzo, D.; Nelson, K.K.; Laurent, L.C.; Parast, M.M. Hypoxia directs human extravillous trophoblast differentiation in a hypoxia-inducible factor-dependent manner. Am. J. Pathol. 2017, 187, 767–780. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Xiao, D.; Hu, Y.; Wang, Z.; Paradis, A.; Mata-Greenwood, E.; Zhang, L. Gestational hypoxia induces preeclampsia-like symptoms via heightened endothelin-1 signaling in pregnant rats. Hypertension 2013, 62, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Kimball, R.; Wayment, M.; Merrill, D.; Wahlquist, T.; Reynolds, P.R.; Arroyo, J.A. Hypoxia reduces placental mTOR activation in a hypoxia-induced model of intrauterine growth restriction (IUGR). Physiol. Rep. 2015, 3, e12651. [Google Scholar] [CrossRef]
- Lin, C.; He, H.; Cui, N.; Ren, Z.; Zhu, M.; Khalil, R.A. Decreased uterine vascularization and uterine arterial expansive remodeling with reduced matrix metalloproteinase-2 and -9 in hypertensive pregnancy. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H165–H180. [Google Scholar] [CrossRef]
- Li, H.; Ohta, H.; Tahara, Y.; Nakamura, S.; Taguchi, K.; Nakagawa, M.; Oishi, Y.; Goto, Y.; Wada, K.; Kaga, M.; et al. Artificial oxygen carriers rescue placental hypoxia and improve fetal development in the rat pre-eclampsia model. Sci. Rep. 2015, 5, 15271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaete, L.G.; Khan, S.; Neerhof, M.G. Endothelin receptor A antagonism prevents damage to glycogen-rich placental cells following uterine ischemia-reperfusion in the rat. Reprod. Sci. 2016, 23, 1518–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Liu, H.; Zeng, J.; Miao, X.; Huang, W.; Chen, H.; Huang, Y.; Li, Y.; Ye, D. Glucocorticoid exposure in early placentation induces preeclampsia in rats via interfering trophoblast development. Gen. Comp. Endocrinol. 2016, 225, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.J.; Chakraborty, D.; Kubota, K.; Renaud, S.J.; Rumi, M.A. Adaptive mechanisms controlling uterine spiral artery remodeling during the establishment of pregnancy. Int. J. Dev. Biol. 2014, 58, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, D.; Rumi, M.A.; Soares, M.J. NK cells, hypoxia and trophoblast cell differentiation. Cell Cycle 2012, 11, 2427–2430. [Google Scholar] [CrossRef] [Green Version]
- Faas, M.M.; de Vos, P. Uterine NK cells and macrophages in pregnancy. Placenta 2017, 56, 44–52. [Google Scholar] [CrossRef]
- Jena, M.K.; Nayak, N.; Chen, K.; Nayak, N.R. Role of macrophages in pregnancy and related complications. Arch. Immunol. Ther. Exp. 2019, 67, 295–309. [Google Scholar] [CrossRef]
- Mor, G.; Abrahams, V.M. Potential role of macrophages as immunoregulators of pregnancy. Reprod. Biol. Endocrinol. 2003, 1, 119. [Google Scholar] [CrossRef] [Green Version]
- Head, J.R. Uterine natural killer cells during pregnancy in rodents. Nat. Immun. 1996, 15, 7–21. [Google Scholar]
- Verlohren, S.; Geusens, N.; Morton, J.; Verhaegen, I.; Hering, L.; Herse, F.; Dudenhausen, J.W.; Muller, D.N.; Luft, F.C.; Cartwright, J.E.; et al. Inhibition of trophoblast-induced spiral artery remodeling reduces placental perfusion in rat pregnancy. Hypertension 2010, 56, 304–310. [Google Scholar] [CrossRef] [Green Version]
- Groen, B.; Uuldriks, G.A.; de Vos, P.; Visser, J.T.; Links, T.P.; Faas, M.M. Impaired trophoblast invasion and increased numbers of immune cells at day 18 of pregnancy in the mesometrial triangle of type 1 diabetic rats. Placenta 2015, 36, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, B.M.; Almada, M.; Costa, M.A.; Teixeira, N.A.; Correia-da-Silva, G. Rat spontaneous foetal resorption: Altered α2-macroglobulin levels and uNK cell number. Histochem. Cell Biol. 2014, 142, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Melgert, B.N.; Spaans, F.; Borghuis, T.; Klok, P.A.; Groen, B.; Bolt, A.; de Vos, P.; van Pampus, M.G.; Wong, T.Y.; van Goor, H.; et al. Pregnancy and preeclampsia affect monocyte subsets in humans and rats. PLoS ONE 2012, 7, e45229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessier, D.R.; Raha, S.; Holloway, A.C.; Yockell-Lelièvre, J.; Tayade, C.; Gruslin, A. Characterization of immune cells and cytokine localization in the rat utero-placental unit mid- to late gestation. J. Reprod. Immunol. 2015, 110, 89–101. [Google Scholar] [CrossRef]
- Brandes, D.; Anton, E. An electron microscopic cytochemical study of macrophages during uterine involution. J. Cell Biol. 1969, 41, 450–461. [Google Scholar] [CrossRef]
- Padykula, H.A.; Tansey, T.R. The occurrence of uterine stromal and intraepithelial monocytes and heterophils during normal late pregnancy in the rat. Anat. Rec. 1979, 193, 329–356. [Google Scholar] [CrossRef]
- Hamilton, S.; Oomomian, Y.; Stephen, G.; Shynlova, O.; Tower, C.L.; Garrod, A.; Lye, S.J.; Jones, R.L. Macrophages infiltrate the human and rat decidua during term and preterm labor: Evidence that decidual inflammation precedes labor. Biol. Reprod. 2012, 86, 39. [Google Scholar] [CrossRef]
- Renaud, S.J.; Scott, R.L.; Chakraborty, D.; Rumi, M.A.; Soares, M.J. Natural killer-cell deficiency alters placental development in rats. Biol. Reprod. 2017, 96, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, Z.; Smith, G.N.; Croy, B.A. Natural killer cell-triggered vascular transformation: Maternal care before birth? Cell. Mol. Immunol. 2011, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hatta, K.; MacLeod, R.J.; Gerber, S.A.; Croy, B.A. Emerging themes in uterine natural killer cell heterogeneity and function. Am. J. Reprod. Immunol. 2012, 68, 282–289. [Google Scholar] [CrossRef]
- Sharma, S. Natural killer cells and regulatory T cells in early pregnancy loss. Int. J. Dev. Biol. 2014, 58, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arad, M.; Atzil, S.; Shakhar, K.; Adoni, A.; Ben-Eliyahu, S. Poly I-C induces early embryo loss in F344 rats: A potential role for NK cells. Am. J. Reprod. Immunol. 2005, 54, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Reister, F.; Frank, H.G.; Kingdom, J.C.; Heyl, W.; Kaufmann, P.; Rath, W.; Huppertz, B. Macrophage-induced apoptosis limits endovascular trophoblast invasion in the uterine wall of preeclamptic women. Lab. Investig. 2001, 81, 1143–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahams, V.M.; Kim, Y.M.; Straszewski, S.L.; Romero, R.; Mor, G. Macrophages and apoptotic cell clearance during pregnancy. Am. J. Reprod. Immunol. 2004, 51, 275–282. [Google Scholar] [CrossRef]
- Cotechini, T.; Komisarenko, M.; Sperou, A.; Macdonald-Goodfellow, S.; Adams, M.A.; Graham, C.H. Inflammation in rat pregnancy inhibits spiral artery remodeling leading to fetal growth restriction and features of preeclampsia. J. Exp. Med. 2014, 211, 165–179. [Google Scholar] [CrossRef]
- Phillips, P.; Brown, M.B.; Progulske-Fox, A.; Wu, X.J.; Reyes, L. Porphyromonas gingivalis strain-dependent inhibition of uterine spiral artery remodeling in the pregnant rat. Biol. Reprod. 2018, 99, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.J.; Cotechini, T.; Quirt, J.S.; Macdonald-Goodfellow, S.K.; Othman, M.; Graham, C.H. Spontaneous pregnancy loss mediated by abnormal maternal inflammation in rats is linked to deficient uteroplacental perfusion. J. Immunol. 2011, 186, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Falcón, B.J.; Cotechini, T.; Macdonald-Goodfellow, S.K.; Othman, M.; Graham, C.H. Abnormal inflammation leads to maternal coagulopathies associated with placental haemostatic alterations in a rat model of foetal loss. Thromb. Haemost. 2012, 107, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Cotechini, T.; Hopman, W.J.; Graham, C.H. Inflammation-induced fetal growth restriction in rats is associated with altered placental morphometrics. Placenta 2014, 35, 575–581. [Google Scholar] [CrossRef]
- Gong, P.; Liu, M.; Hong, G.; Li, Y.; Xue, P.; Zheng, M.; Wu, M.; Shen, L.; Yang, M.; Diao, Z.; et al. Curcumin improves LPS-induced preeclampsia-like phenotype in rat by inhibiting the TLR4 signaling pathway. Placenta 2016, 41, 45–52. [Google Scholar] [CrossRef]
- Robb, K.P.; Cotechini, T.; Allaire, C.; Sperou, A.; Graham, C.H. Inflammation-induced fetal growth restriction in rats is associated with increased placental HIF-1α accumulation. PLoS ONE 2017, 12, e0175805. [Google Scholar] [CrossRef] [PubMed]
- Xue, P.; Zheng, M.; Gong, P.; Lin, C.; Zhou, J.; Li, Y.; Shen, L.; Diao, Z.; Yan, G.; Sun, H.; et al. Single administration of ultra-low-dose lipopolysaccharide in rat early pregnancy induces TLR4 activation in the placenta contributing to preeclampsia. PLoS ONE 2015, 10, e0124001. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Li, X.; Gao, X.; Dong, L.; Xin, G.; Chen, L.; Qiu, J.; Xu, Y. LPS induces preeclampsia-like phenotype in rats and HTR8/SVneo cells dysfunction through TLR4/p38 MAPK pathway. Front. Physiol. 2019, 10, 1030. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.H.; Wang, L.L.; Liu, H.; Muyayalo, K.P.; Huang, X.B.; Mor, G.; Liao, A.H. Galectin-9 alleviates LPS-induced preeclampsia-like impairment in rats via switching decidual macrophage polarization to M2 subtype. Front. Immunol. 2019, 9, 3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozai, K.; Iqbal, K.; Moreno-Irusta, A.; Scott, R.L.; Simon, M.E.; Dhakal, P.; Fields, P.E.; Soares, M.J. Protective role of IL33 signaling in negative pregnancy outcomes associated with lipopolysaccharide exposure. FASEB J. 2021, 35, e21272. [Google Scholar] [CrossRef]
- Spaans, F.; Melgert, B.N.; Chiang, C.; Borghuis, T.; Klok, P.A.; de Vos, P.; van Goor, H.; Bakker, W.W.; Faas, M.M. Extracellular ATP decreases trophoblast invasion, spiral artery remodeling and immune cells in the mesometrial triangle in pregnant rats. Placenta 2014, 35, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Waddell, B.J.; Hisheh, S.; Dharmarajan, A.M.; Burton, P.J. Apoptosis in rat placenta is zone dependent and stimulated by glucocorticoids. Biol. Reprod. 2000, 63, 1913–1917. [Google Scholar] [CrossRef]
- Ain, R.; Canham, L.N.; Soares, M.J. Dexamethasone-induced intrauterine growth restriction impacts the placental prolactin family, insulin-like growth factor-II and the Akt signaling pathway. J. Endocrinol. 2005, 185, 253–263. [Google Scholar] [CrossRef]
- Yokoi, R.; Suda, S.; Kobayashi, K.; Kuroda, J.; Kusama, H.; Kagami, H.; Ono, T. Differential susceptibility of rat embryos to methyl methanesulfonate during the pregastrulation period. J. Toxicol. Sci. 2007, 32, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Usuda, K.; Abe, M.; Hayashi, S.; Ogawa, I. Effect of 6-mercaptopurine on rat placenta. J. Vet. Med. Sci. 2008, 70, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Hayashi, S.; Usuda, K.; Abe, M.; Ogawa, I. The relationship between fetal growth restriction and small placenta in 6-mercaptopurine exposed rat. Exp. Toxicol. Pathol. 2011, 63, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Taki, K.; Fukushima, T.; Ise, R.; Horii, I.; Yoshida, T. 6-Mercaptopurine-induced histopathological changes and xanthine oxidase expression in rat placenta. J. Toxicol. Sci. 2012, 37, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Hayashi, S.; Usuda, K.; Abe, M.; Hagio, S.; Ogawa, I. Effect of cisplatin on rat placenta development. Exp. Toxicol. Pathol. 2013, 65, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Sugiyama, A.; Inoue, S.; Takeuchi, T.; Takeuchi, T.; Furukawa, S. Effect of methotrexate on rat placenta development. Exp. Toxicol. Pathol. 2013, 65, 995–1002. [Google Scholar] [CrossRef]
- Furukawa, S.; Hayashi, S.; Abe, M.; Hagio, S.; Irie, K.; Kuroda, Y.; Ogawa, I.; Sugiyama, A. Effect of chlorpromazine on rat placenta development. Exp. Toxicol. Pathol. 2014, 66, 41–47. [Google Scholar] [CrossRef]
- Lee, C.K.; Lee, C.U.; Kim, J.H.; Son, B.C.; Kim, D.H.; Lee, C.H.; Kim, H.D.; Kim, J.W.; Yoon, Y.D.; Kang, S.G.; et al. Effects of Aroclor 1254 on the expression of rat placental PRL-family genes. Mol. Cells. 2003, 15, 114–121. [Google Scholar]
- Nishimura, K.; Nakano, N.; Chowdhury, V.S.; Kaneto, M.; Torii, M.; Hattori, M.A.; Yamauchi, N.; Kawai, M. Effect of PPARβ/δ agonist on the placentation and embryo-fetal development in rats. Birth Defects Res. B Dev. Reprod. Toxicol. 2013, 98, 164–169. [Google Scholar] [CrossRef]
- Holloway, A.C.; Salomon, X.A.; Soares, M.J.; Garnier, V.; Raha, S.; Sergent, F.; Nicholson, C.J.; Feige, J.J.; Benharouga, M.; Alfaidy, N. Characterization of the adverse effects of nicotine on placental development: In vivo and in vitro studies. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E443–E456. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Maternal thyroid dysfunction affects placental profile of inflammatory mediators and the intrauterine trophoblast migration kinetics. Reproduction 2014, 147, 803–816. [Google Scholar] [CrossRef]
- Chakraborty, S.; Islam, S.; Saha, S.; Ain, R. Dexamethasone-induced intra-uterine growth restriction impacts nostrin and its downstream effector genes in the rat mesometrial uterus. Sci. Rep. 2018, 8, 8342. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Tsuji, N.; Hayashi, S.; Kuroda, Y.; Kimura, M.; Hayakawa, C.; Takeuchi, K.; Sugiyama, A. The effects of β-naphthoflavone on rat placental development. J. Toxicol. Pathol. 2019, 32, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Hayashi, S.; Usuda, K.; Abe, M.; Ogawa, I. The impairment of metrial gland development in tamoxifen exposed rats. Exp. Toxicol. Pathol. 2012, 64, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Pierce, S.H.; Kozai, K.; Dhakal, P.; Scott, R.L.; Roby, K.F.; Vyhlidal, C.A.; Soares, M.J. Evaluation of placentation and the role of the aryl hydrocarbon receptor pathway in a rat model of dioxin exposure. Environ. Health Perspect. 2021, 129, 117001. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.; Piskorski, A.; Kao, J.C.; Hess, L.A.; de la Monte, S.; Gündoğan, F. Placental Morphology in Fetal Alcohol Spectrum Disorders. Alcohol Alcohol. 2017, 52, 138–144. [Google Scholar] [CrossRef]
- Gundogan, R.; Gilligan, J.; Ooi, J.-H.; Sung, J.; Qi, W.; Naram, R.; de la Monte, S.M. Dual mechanisms of ethanol-impaired placentation: Experimental model. Clin. Exper. Pathol. 2013, 3, 2. [Google Scholar] [CrossRef]
- Gårdebjer, E.M.; Cuffe, J.S.; Pantaleon, M.; Wlodek, M.E.; Moritz, K.M. Periconceptional alcohol consumption causes fetal growth restriction and increases glycogen accumulation in the late gestation rat placenta. Placenta 2014, 35, 50–57. [Google Scholar] [CrossRef]
- Gundogan, F.; Elwood, G.; Longato, L.; Tong, M.; Feijoo, A.; Carlson, R.I.; Wands, J.R.; de la Monte, S.M. Impaired placentation in fetal alcohol syndrome. Placenta 2008, 29, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Gundogan, F.; Gilligan, J.; Qi, W.; Chen, E.; Naram, R.; de la Monte, S.M. Dose effect of gestational ethanol exposure on placentation and fetal growth. Placenta 2015, 36, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Akay, M.T.; Kockaya, E.A. The effects of alcohol on rat placenta. Cell Biochem. Funct. 2005, 23, 435–445. [Google Scholar]
- Furukawa, S.; Hayashi, S.; Usuda, K.; Abe, M.; Ogawa, I. Histopathological effect of ketoconazole on rat placenta. J. Vet. Med. Sci. 2008, 70, 1179–1184. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Flores, E.; Yáñez Mendoza, C. Histology of rat placentas treated with methylhydrazine Ro 4-6467. Bol. Estud. Med. Biol. 1990, 38, 59–64. [Google Scholar] [PubMed]
- Ahbab, M.A.; Güven, C.; Koçkaya, E.A.; Barlas, N. Comparative developmental toxicity evaluation of di- n-hexyl phthalate and dicyclohexyl phthalate in rats. Toxicol. Ind. Health 2017, 33, 696–716. [Google Scholar] [CrossRef] [PubMed]
- Jawerbaum, A.; White, V. Animal models in diabetes and pregnancy. Endocr. Rev. 2010, 31, 680–701. [Google Scholar] [CrossRef] [Green Version]
- Gewolb, I.H.; Merdian, W.; Warshaw, J.B.; Enders, A.C. Fine structural abnormalities of the placenta in diabetic rats. Diabetes 1986, 35, 1254–1261. [Google Scholar] [CrossRef]
- Ne’eman, Z.; Barash, V.; Rosenmann, E.; Shafrir, E. Localization of glycogen in the placenta of diabetic rats: A light and electron microscopic study. Placenta 1987, 8, 201–208. [Google Scholar] [CrossRef]
- Zorn, T.M.; Zúñiga, M.; Madrid, E.; Tostes, R.; Fortes, Z.; Giachini, F.; San Martín, S. Maternal diabetes affects cell proliferation in developing rat placenta. Histol. Histopathol. 2011, 26, 1049–1056. [Google Scholar] [PubMed]
- Farias, P.S.; dos SSouza, K.; Fioretto, E.T.; dos Santos, M.R.; Aires, M.B. Maternal diabetes affects rat placental morphology and pregnancy. Endocrine 2014, 45, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Nteeba, J.; Varberg, K.M.; Scott, R.L.; Simon, M.E.; Iqbal, K.; Soares, M.J. Poorly controlled diabetes mellitus alters placental structure, efficiency, and plasticity. BMJ Open Diabetes Res. Care 2020, 8, e001243. [Google Scholar] [CrossRef]
- Phillips, J.K.; Vance, A.M.; Raj, R.S.; Mandalà, M.; Linder, E.A.; Gokina, N.I. Impact of experimental diabetes on the maternal uterine vascular remodeling during rat pregnancy. Reprod. Sci. 2012, 19, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Fisher, S.J. Why is placentation abnormal in preeclampsia? Am. J. Obstet. Gynecol. 2015, 213, S115–S122. [Google Scholar] [CrossRef] [Green Version]
- Bakrania, B.A.; George, E.M.; Granger, J.P. Animal models of preeclampsia: Investigating pathophysiology and therapeutic targets. Am. J. Obstet. Gynecol. 2020, 226, S973–S987. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, challenges, and perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Croskerry, P.G.; Smith, G.K.; Hall, S.; Shepard, B.J. Fetal and placental growth in the rat following differential reduction of litter size. Biol. Neonate 1978, 33, 31–38. [Google Scholar] [CrossRef]
- vom Saal, F.S. Variation in phenotype due to random intrauterine positioning of male and female fetuses in rodents. J. Reprod. Fertil. 1981, 62, 633–650. [Google Scholar] [CrossRef] [Green Version]
- McLaurin, K.A.; Mactutus, C.F. Polytocus focus: Uterine position effect is dependent upon horn size. Int. J. Dev. Neurosci. 2015, 40, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nüsken, K.D.; Warnecke, C.; Hilgers, K.F.; Schneider, H. Intrauterine growth after uterine artery ligation in rats: Dependence on the fetal position in the uterine horn and need for prenatal marking of the animals. J. Hypertens. 2007, 25, 247–248. [Google Scholar] [CrossRef]
- Bohlender, J.; Ganten, D.; Luft, F.C. Rats transgenic for human renin and human angiotensinogen as a model for gestational hypertension. J. Am. Soc. Nephrol. 2000, 11, 2056–2061. [Google Scholar] [CrossRef]
- Dechend, R.; Gratze, P.; Wallukat, G.; Shagdarsuren, E.; Plehm, R.; Bräsen, J.H.; Fiebeler, A.; Schneider, W.; Caluwaerts, S.; Vercruysse, L.; et al. Agonistic autoantibodies to the AT1 receptor in a transgenic rat model of preeclampsia. Hypertension 2005, 45, 742–746. [Google Scholar] [CrossRef] [Green Version]
- Verlohren, S.; Niehoff, M.; Hering, L.; Geusens, N.; Herse, F.; Tintu, A.N.; Plagemann, A.; LeNoble, F.; Pijnenborg, R.; Muller, D.N.; et al. Uterine vascular function in a transgenic preeclampsia rat model. Hypertension 2008, 51, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Geusens, N.; Verlohren, S.; Luyten, C.; Taube, M.; Hering, L.; Vercruysse, L.; Hanssens, M.; Dudenhausen, J.W.; Dechend, R.; Pijnenborg, R. Endovascular trophoblast invasion, spiral artery remodelling and uteroplacental haemodynamics in a transgenic rat model of pre-eclampsia. Placenta 2008, 29, 614–623. [Google Scholar] [CrossRef]
- Geusens, N.; Hering, L.; Verlohren, S.; Luyten, C.; Drijkoningen, K.; Taube, M.; Vercruysse, L.; Hanssens, M.; Dechend, R.; Pijnenborg, R. Changes in endovascular trophoblast invasion and spiral artery remodelling at term in a transgenic preeclamptic rat model. Placenta 2010, 31, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Hering, L.; Herse, F.; Geusens, N.; Verlohren, S.; Wenzel, K.; Staff, A.C.; Brosnihan, K.B.; Huppertz, B.; Luft, F.C.; Muller, D.N.; et al. Effects of circulating and local uteroplacental angiotensin II in rat pregnancy. Hypertension 2010, 56, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golic, M.; Haase, N.; Herse, F.; Wehner, A.; Vercruysse, L.; Pijnenborg, R.; Balogh, A.; Saether, P.C.; Dissen, E.; Luft, F.C.; et al. Natural killer cell reduction and uteroplacental vasculopathy. Hypertension 2016, 68, 964–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, K.; Aoki, K. Development of a strain of spontaneously hypertensive rats. Jpn. Circ. J. 1963, 27, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Barrientos, G.; Pussetto, M.; Rose, M.; Staff, A.C.; Blois, S.M.; Toblli, J.E. Defective trophoblast invasion underlies fetal growth restriction and preeclampsia-like symptoms in the stroke-prone spontaneously hypertensive rat. Mol. Hum. Reprod. 2017, 23, 509–519. [Google Scholar] [CrossRef]
- Li, J.; LaMarca, B.; Reckelhoff, J.F. A model of preeclampsia in rats: The reduced uterine perfusion pressure (RUPP) model. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1–H8. [Google Scholar] [CrossRef]
- Coats, L.E.; Bamrick-Fernandez, D.R.; Ariatti, A.M.; Bakrania, B.A.; Rawls, A.Z.; Ojeda, N.B.; Alexander, B.T. Stimulation of soluble guanylate cyclase diminishes intrauterine growth restriction in a rat model of placental ischemia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R149–R161. [Google Scholar] [CrossRef]
- Podjarny, E.; Bernheim, J.; Katz, B.; Green, J.; Mekler, J.; Bursztyn, M. Chronic exogenous hyperinsulinaemia in pregnancy: A rat model of pregnancy-induced hypertension. J. Am. Soc. Nephrol. 1998, 9, 9–13. [Google Scholar] [CrossRef]
- Skarzinski, G.; Khamaisi, M.; Bursztyn, M.; Mekler, J.; Lan, D.; Evdokimov, P.; Ariel, I. Intrauterine growth restriction and shallower implantation site in rats with maternal hyperinsulinemia are associated with altered NOS expression. Placenta 2009, 30, 898–906. [Google Scholar] [CrossRef]
- Kreiser, D.; Nguyen, X.; Wong, R.; Seidman, D.; Stevenson, D.; Quan, S.; Abraham, N.; Dennery, P.A. Heme oxygenase-1 modulates fetal growth in the rat. Lab. Investig. 2002, 82, 687–692. [Google Scholar] [CrossRef] [Green Version]
- Oltra, L.; Reverte, V.; Garcés, B.; Li Volti, G.; Moreno, J.M.; Salazar, F.J.; Llinás, M.T. Trophoblast-induced spiral artery remodelling and uteroplacental haemodynamics in pregnant rats with increased blood pressure induced by heme oxygenase inhibition. Placenta 2020, 89, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, K.M.; Tsai, K.Y.; Davis, T.; Clark, J.C.; Knowlton, M.N.; Bikman, B.T.; Reynolds, P.R.; Arroyo, J.A. Growth arrest-specific protein-6/AXL signaling induces preeclampsia in rats. Biol. Reprod. 2020, 102, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, L.; Desai, M.; Nelson, D.M.; Ross, M.G. Altered mitochondrial apoptotic pathway in placentas from undernourished rat gestations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1599–R1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebelato, H.J.; Esquisatto, M.A.; de Sousa Righi, E.F.; Catisti, R. Gestational protein restriction alters cell proliferation in rat placenta. J. Mol. Histol. 2016, 47, 203–211. [Google Scholar] [CrossRef]
- Padmanabhan, R.; Al-Menhali, N.M.; Ahmed, I.; Kataya, H.H.; Ayoub, M.A. Histological, histochemical and electron microscopic changes of the placenta induced by maternal exposure to hyperthermia in the rat. Int. J. Hyperth. 2005, 21, 29–44. [Google Scholar] [CrossRef]
- Roberts, H.; Woodman, A.G.; Baines, K.J.; Jeyarajah, M.J.; Bourque, S.L.; Renaud, S.J. Maternal iron deficiency alters trophoblast differentiation and placental development in rat pregnancy. Endocrinology 2021, 162, bqab215. [Google Scholar] [CrossRef]
- Mark, P.J.; Sisala, C.; Connor, K.; Patel, R.; Lewis, J.L.; Vickers, M.H.; Waddell, B.J.; Sloboda, D.M. A maternal high-fat diet in rat pregnancy reduces growth of the fetus and the placental junctional zone, but not placental labyrinth zone growth. J. Dev. Origins Health Dis. 2011, 2, 63–70. [Google Scholar] [CrossRef]
- Hayes, E.K.; Tessier, D.R.; Percival, M.E.; Holloway, A.C.; Petrik, J.J.; Gruslin, A.; Raha, S. Trophoblast invasion and blood vessel remodeling are altered in a rat model of lifelong maternal obesity. Reprod. Sci. 2014, 21, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Faria, T.N.; Soares, M.J. Trophoblast cell differentiation: Establishment, characterization, and modulation of a rat trophoblast cell line expressing members of the placental prolactin family. Endocrinology 1991, 129, 2895–2906. [Google Scholar] [CrossRef]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane, K.; Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of human trophoblast stem cells. Cell Stem Cell 2018, 22, 50–63.e6. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, J.A.; Konno, T.; Khalili, D.C.; Soares, M.J. A simple in vivo approach to investigate invasive trophoblast cells. Int. J. Dev. Biol. 2005, 49, 977–980. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Rumi, M.A.; Konno, T.; Soares, M.J. In vivo genetic manipulation of the rat trophoblast cell lineage using lentiviral vector delivery. Genesis 2009, 47, 433–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, D.; Muto, M.; Soares, M.J. Ex vivo trophoblast-specific genetic manipulation using lentiviral delivery. Bio. Protoc. 2017, 7, e2652. [Google Scholar] [CrossRef] [Green Version]
- Varberg, K.M.; Iqbal, K.; Muto, M.; Simon, M.E.; Scott, R.L.; Kozai, K.; Choudhury, R.H.; Aplin, J.D.; Biswell, R.; Gibson, M.; et al. ASCL2 reciprocally controls key trophoblast lineage decisions during hemochorial placenta development. Proc. Natl. Acad. Sci. USA 2021, 118, e2016517118. [Google Scholar] [CrossRef] [PubMed]
- Kent, L.N.; Konno, T.; Soares, M.J. Phosphatidylinositol 3 kinase modulation of trophoblast cell differentiation. BMC Dev. Biol. 2010, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Kent, L.N.; Rumi, M.A.; Kubota, K.; Lee, D.S.; Soares, M.J. FOSL1 is integral to establishing the maternal-fetal interface. Mol. Cell Biol. 2011, 31, 4801–4813. [Google Scholar] [CrossRef] [Green Version]
- Kubota, K.; Kent, L.N.; Rumi, M.A.; Roby, K.F.; Soares, M.J. Dynamic regulation of AP-1 transcriptional complexes directs trophoblast differentiation. Mol. Cell Biol. 2015, 35, 3163–3177. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.J.; Kubota, K.; Rumi, M.A.; Soares, M.J. The FOS transcription factor family differentially controls trophoblast migration and invasion. J. Biol. Chem. 2014, 289, 5025–5039. [Google Scholar] [CrossRef] [Green Version]
- Amoroso, E.C. The evolution of viviparity. Proc. R. Soc. Med. 1968, 61, 1188–1200. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Experimental Maternal Manipulation | Phenotype of the Uterine-Placental Interface | References |
|---|---|---|
| Oxygen | ||
| Moderate hypoxia during placental morphogenesis | Expansion of the junctional zone; enhanced endovascular invasive trophoblast restructuring of uterine spiral arteries | [46,63,64] |
| Severe/chronic hypoxia | Impairments in junctional zone development and trophoblast-guided spiral artery remodeling | [70,71,72,73,74] |
| Immune cells | ||
| NK cells | Remodel uterine arterial vessels; restrain intrauterine endovascular trophoblast cell invasion | [47,90] |
| Macrophages | Uterine angiogenesis, tissue repair, artery remodeling, immune tolerance, postpartum trophoblast removal | [78,83,85,86,87,88,89] |
| Drug, toxicant, and miscellaneous exposure | ||
| Anti-cancer drugs, dexa-methasone, AHR agonists, GW501516, nicotine | Hypoplasia of the junctional zone, abnormalities in glycogen trophoblast cell development, and/or decreased intrauterine interstitial trophoblast cell invasion | [109,110,111,112,113,114,115,116,117,118,119,120,121,122,123] |
| Tamoxifen | Disrupts the uterine-placental interface; decreases NK cell numbers; defective uterine spiral artery transformation | [124] |
| Doxycycline | Impairs endovascular trophoblast cell invasion and uterine spiral artery remodeling and placental perfusion | [82] |
| Dioxin | Accelerates endovascular trophoblast cell invasion | [125] |
| Chronic ethanol | Decreased glycogen trophoblast cell numbers; shallow trophoblast invasion; failed uterine spiral artery remodeling | [127,129,130,131] |
| Ketoconazole, methylhydrazine | Expansion of junctional zone and glycogen trophoblast cell clusters | [132,133] |
| Disease states | ||
| Diabetes: assorted hyperglycemia models | Junctional zone and glycogen trophoblast cell expansion; impaired trophoblast invasion and spiral artery remodeling; retention/expansion of NK cells and macrophages | [136,137,138,139,140,141] |
| Hypertension/preeclampsia: Angiotensinogen-renin transgenic model | Enhanced endovascular trophoblast cell invasion and uterine spiral artery remodeling | [152,153,154] |
| Hypertension/preeclampsia: Stroke prone spontaneous hypertensive rat (SHSRP) | Impaired junctional zone and glycogen trophoblast cell development; shallow trophoblast cell invasion; failed uterine spiral artery remodeling | [157] |
| Hypertension/preeclampsia: Reduced uterine perfusion pressure (RUPP) model | Expanded junctional zone and impairments in intrauterine trophoblast cell invasion and uterine spiral artery remodeling | [72,158,159] |
| Hypertension/preeclampsia: Maternal hyperinsulinemia | Enhanced endovascular trophoblast cell invasion | [161] |
| Hypertension/preeclampsia: Hemoxygenase inhibition | Diminished endovascular trophoblast cell-guided uterine spiral artery remodeling | [163] |
| Hypertension/preeclampsia: GAS6 treatment | Decreased interstitial trophoblast cell invasion | [164] |
| Malnutrition/protein restriction/hyperthermia | Negative impact on junctional zone development | [165,166,167] |
| Iron deficiency | Expanded junctional zone | [168] |
| High fat diet | Diminished junctional zone; early increase in intrauterine trophoblast invasion; ate gestation decline in interstitial trophoblast cell invasion | [169,170] |
| Genes | Mutation | Phenotype | References |
|---|---|---|---|
| Fosl1 | Trophoblast-LOF a | Inhibition of endovascular trophoblast cell invasion | [178] |
| Kdm3a | Trophoblast-LOF | Disruption of hypoxia-activated endovascular trophoblast cell invasion | [64] |
| Mmp12 | Global-LOF | Disruption of hypoxia-activated endovascular trophoblast cell invasion | [64] |
| Ascl2 | Global-LOF | Junctional zone dysgenesis and failed intrauterine trophoblast cell invasion | [176] |
| Ahr | Global-LOF | Maternal AHR regulates TCDD-induced endovascular trophoblast cell invasion | [125] |
| Tfpi | Trophoblast-LOF | Inhibition of endovascular and interstitial trophoblast cell invasion | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shukla, V.; Soares, M.J. Modeling Trophoblast Cell-Guided Uterine Spiral Artery Transformation in the Rat. Int. J. Mol. Sci. 2022, 23, 2947. https://doi.org/10.3390/ijms23062947
Shukla V, Soares MJ. Modeling Trophoblast Cell-Guided Uterine Spiral Artery Transformation in the Rat. International Journal of Molecular Sciences. 2022; 23(6):2947. https://doi.org/10.3390/ijms23062947
Chicago/Turabian StyleShukla, Vinay, and Michael J. Soares. 2022. "Modeling Trophoblast Cell-Guided Uterine Spiral Artery Transformation in the Rat" International Journal of Molecular Sciences 23, no. 6: 2947. https://doi.org/10.3390/ijms23062947