
Growth Repressor GmRAV Binds to the GmGA3ox Promoter to Negatively Regulate Plant Height Development in Soybean

Abstract
:1. Introduction
2. Results
2.1. GmRAV Inhibited Plant Height and Yield in Soybean
2.2. Dwarfism of GmRAV-ox Soybean Rescued by Exogenous GA3
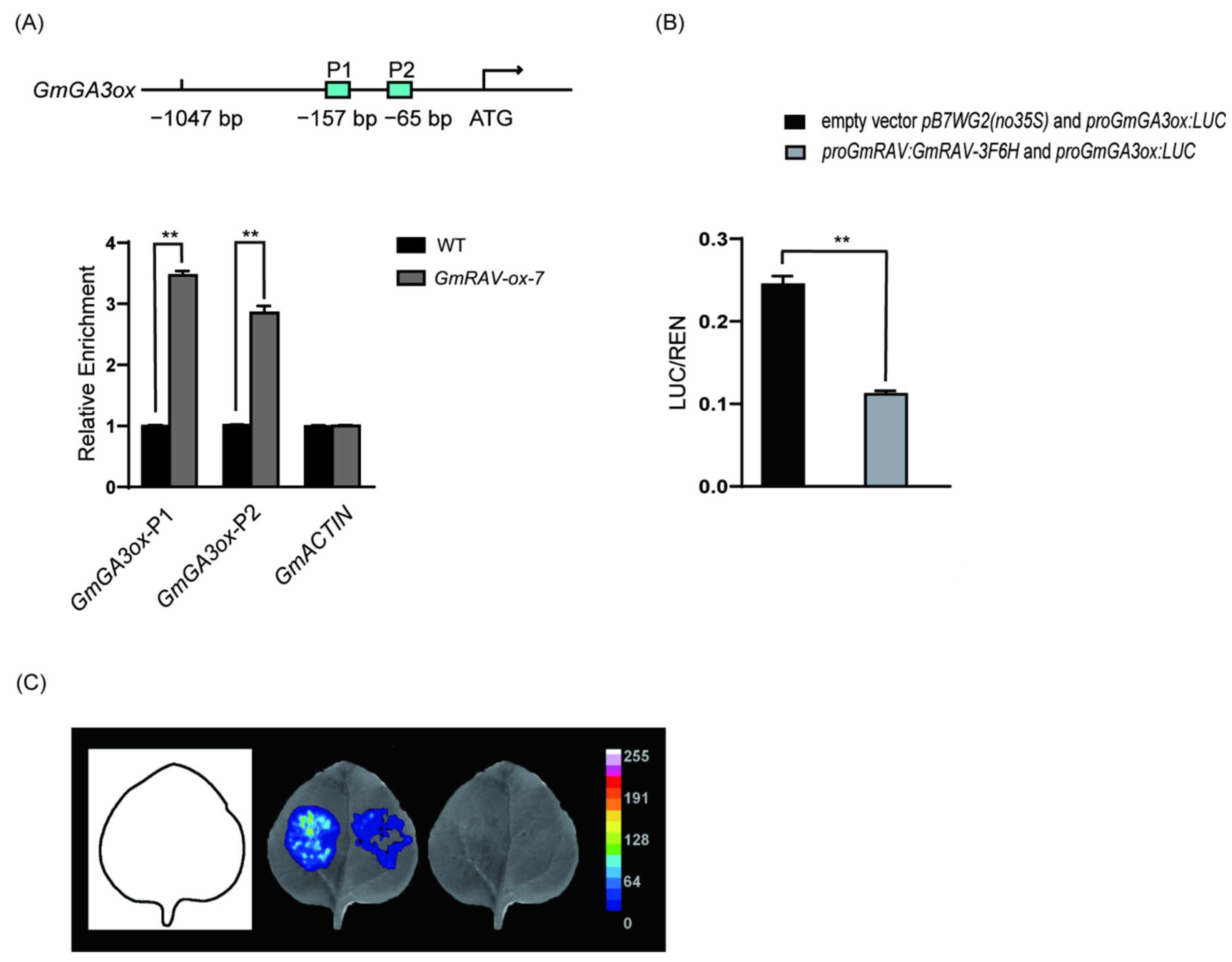
2.3. Identification of GmGA3ox as Direct Target of GmRAV
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions and Records of Data
4.2. Plasmids Construction
4.3. Quantitative Real-Time RT-PCR
4.4. ChIP-qPCR
4.5. Transient Assay of GmGA3ox Promoter Affected by GmRAV Protein in N. benthamiana
4.6. Hypocotyl and Stem Growth Assay of Seedlings
4.7. Scanning Electron Microscopy
4.8. Endogenous GA3 Determination
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| qGmGA3ox 1-ChIP-F | TCTCCCTCAACTTTCAGTTCCA |
| qGmGA3ox 1-ChIP-R | TCAATTCTCCCTTTCAGCCTTC |
| qGmGA3ox 2-ChIP-F | TCCTGAAGGCTGAAAGGGAGAA |
| qGmGA3ox 2-ChIP-R | ACCGCATGGAAATTAGATAGAG |
| qGmGA3ox-F | AGCCATGATGATCATACTCCTG |
| qGmGA3ox-R | CAAGTTATGCATGCATGGTGTA |
| proGmGA3ox: LUC-F | GAATTCCTGCAGCCCAACCGATTCAAATTGAGGGTAT |
| proGmGA3ox: LUC-R | ACTAGTGGATCCCCCACCGCATGGAAATTAGATAGAG |
| qGmACTIN4-F | GTGTCAGCCATACTGTCCCCATTT |
| qGmACTIN4-R | GTTTCAAGCTCTTGCTCGTAATCA |
References
- Anderson, E.J.; Ali, M.L.; Beavis, W.D.; Chen, P.; Clemente, T.E.; Diers, B.W.; Graef, G.L.; Grassini, P.; Hyten, D.L.; McHale, L.K. Soybean [Glycine max (L.) Merr.] breeding: History, improvement, production and future opportunities. In Advances in Plant Breeding Strategies: Legumes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 431–516. [Google Scholar]
- Hu, M.; Wiatrak, P. Effect of planting date on soybean growth, yield, and grain quality. Agron. J. 2012, 104, 785–790. [Google Scholar] [CrossRef]
- Donald, C.T. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Cooper, R.; Mendiola, T.; St Martin, S.; Fioritto, R.; Dorrance, A. Registration of ‘Apex’ soybean (Registration of Cultivars). Crop Sci. 2003, 43, 1563–1564. [Google Scholar] [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Ellis, M.H.; Chandler, P.M. Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. USA 2002, 99, 9043–9048. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Guo, Y.; Ou, L.; Hong, H.; Wang, J.; Liu, Z.; Guo, B.; Zhang, L.; Qiu, L. Identification of the dwarf gene GmDW1 in soybean (Glycine max L.) by combining mapping-by-sequencing and linkage analysis. Theor. Appl. Genet. 2018, 131, 1001–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Dong, L.; Su, T.; Li, T.; Gan, Z.; Nan, H.; Lu, S.; Fang, C.; Kong, L.; Li, H.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmLHY genes alters plant height and internode length in soybean. BMC Plant Biol. 2019, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Matías-Hernández, L.; Aguilar-Jaramillo, A.E.; Marín-González, E.; Suárez-López, P.; Pelaz, S. RAV genes: Regulation of floral induction and beyond. Ann. Bot. 2014, 114, 1459–1470. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Inoue, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E.; et al. A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef]
- Swaminathan, K.; Peterson, K.; Jack, T. The plant B3 superfamily. Trends Plant Sci. 2008, 13, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D. The APETALA2 domain is related to a novel type of DNA binding domain. Plant Cell 1995, 7, 388. [Google Scholar] [PubMed] [Green Version]
- Okamuro, J.K.; Caster, B.; Villarroel, R.; Van Montagu, M.; Jofuku, K.D. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 7076–7081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Xu, Z.; Zheng, W.; Zhao, W.; Wang, Y.; Yu, T.; Chen, M.; Zhou, Y.; Min, D.; Ma, Y.; et al. Genome-wide analysis of the RAV family in soybean and functional identification of GmRAV-03 involvement in salt and drought stresses and exogenous ABA treatment. Front. Plant Sci. 2017, 8, 905. [Google Scholar] [CrossRef] [Green Version]
- Osnato, M.; Matias-Hernandez, L.; Aguilar-Jaramillo, A.E.; Kater, M.M.; Pelaz, S. Genes of the RAV family control heading date and carpel development in rice. Plant Physiol. 2020, 183, 1663–1680. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhao, L.; Li, D.; Hao, D.; Zhan, Y.; Li, W. A GmRAV ortholog is involved in photoperiod and sucrose control of flowering time in soybean. PLoS ONE 2014, 9, e89145. [Google Scholar]
- Kagaya, Y.; Ohmiya, K.; Hattori, T. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res. 1999, 27, 470–478. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Sun, J.; Dong, L.; Li, M.; Liu, Y.; Wang, J.; Zhang, X.; Li, D.; Sun, J.; et al. GmRAV confers ecological adaptation through photoperiod control of flowering time and maturity in soybean. Plant Physiol. 2021, 187, 361–377. [Google Scholar] [CrossRef]
- Itoh, H.; Ueguchi-Tanaka, M.; Sentoku, N.; Kitano, H.; Matsuoka, M.; Kobayashi, M. Cloning and functional analysis of two gibberellin 3β-hydroxylase genes that are differently expressed during the growth of rice. Proc. Natl. Acad. Sci. USA 2001, 98, 8909–8914. [Google Scholar] [CrossRef] [Green Version]
- Chiang, H.; Hwang, I.; Goodman, H. Isolation of the Arabidopsis GA4 locus. Plant Cell 1995, 7, 195–201. [Google Scholar]
- Cheng, Q.; Gan, Z.; Wang, Y.; Lu, S.; Hou, Z.; Li, H.; Xiang, H.; Liu, B.; Kong, F.; Dong, L. The soybean gene J contributes to salt stress tolerance by up-regulating salt-responsive genes. Front. Plant Sci. 2020, 11, 272. [Google Scholar]
- Wang, H.; Li, Y.; Chern, M.; Zhu, Y.; Zhang, L.; Lu, J.; Li, X.; Dang, W.-Q.; Ma, X.-C.; Yang, Z.-R.; et al. Suppression of rice miR168 improves yield, flowering time and immunity. Nat. Plants 2021, 7, 129–136. [Google Scholar] [CrossRef]
- Dong, L.; Fang, C.; Cheng, Q.; Su, T.; Kou, K.; Kong, L.; Zhang, C.; Li, H.; Hou, Z.; Zhang, Y.; et al. Genetic basis and adaptation trajectory of soybean from its temperate origin to tropics. Nat. Commun. 2021, 12, 1–11. [Google Scholar]
- Reinhardt, D.; Kuhlemeier, C. Plant architecture. EMBO Reports 2002, 3, 846–851. [Google Scholar] [CrossRef]
- Kaneko, M.; Itoh, H.; Inukai, Y.; Sakamoto, T.; Ueguchi-Tanaka, M.; Ashikari, M.; Matsuoka, M. Where do gibberellin biosynthesis and gibberellin signaling occur in rice plants? Plant J. 2003, 35, 104–115. [Google Scholar] [CrossRef]
- Sun, T.; Gubler, F. Molecular mechanism of gibberellin signaling in plants. Annu. Rev. Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Jing, Y.; Zhao, X.; Wang, J.; Lian, M.; Teng, W.; Qiu, L.; Han, Y.; Li, W. Identification of loci and candidate genes for plant height in soybean (Glycine max) via genome-wide association study. Plant Breed. 2019, 138, 721–732. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, H.; Fan, M.; He, Y.; Guo, P. A mutation in the intron splice acceptor site of a GA3ox gene confers dwarf architecture in watermelon (Citrullus lanatus L.). Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Osnato, M.; Castillejo, C.; Matías-Hernández, L.; Pelaz, S. TEMPRANILLO genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis. Nat. Commun. 2012, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, A.H.; Raghuram, B.; Eschen-Lippold, L.; Scheel, D.; Lee, J.; Sinha, A.K. Agroinfiltration by cytokinin-producing Agrobacterium sp. strain GV3101 primes defense responses in Nicotiana tabacum. Mol. Plant-Microbe Interact. 2014, 27, 1175–1185. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, X.; Shan, J.; Li, Y.; Zhang, Y.; Wang, Y.; Li, W.; Zhao, L. Overexpression of GmGAMYB accelerates the transition to flowering and increases plant height in soybean. Front. Plant Sci. 2021, 12, 667242. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Y.; Zhang, Y.; Shan, J.; Ji, Y.; Zhang, X.; Li, W.; Li, D.; Zhao, L. Growth Repressor GmRAV Binds to the GmGA3ox Promoter to Negatively Regulate Plant Height Development in Soybean. Int. J. Mol. Sci. 2022, 23, 1721. https://doi.org/10.3390/ijms23031721
Xue Y, Zhang Y, Shan J, Ji Y, Zhang X, Li W, Li D, Zhao L. Growth Repressor GmRAV Binds to the GmGA3ox Promoter to Negatively Regulate Plant Height Development in Soybean. International Journal of Molecular Sciences. 2022; 23(3):1721. https://doi.org/10.3390/ijms23031721
Chicago/Turabian StyleXue, Yongguo, Yuntong Zhang, Jinming Shan, Yujia Ji, Xiaoming Zhang, Wenbin Li, Dongmei Li, and Lin Zhao. 2022. "Growth Repressor GmRAV Binds to the GmGA3ox Promoter to Negatively Regulate Plant Height Development in Soybean" International Journal of Molecular Sciences 23, no. 3: 1721. https://doi.org/10.3390/ijms23031721