
PaOctβ2R: Identification and Functional Characterization of an Octopamine Receptor Activating Adenylyl Cyclase Activity in the American Cockroach Periplaneta americana

 , ,
, ,
Abstract
:1. Introduction
2. Results
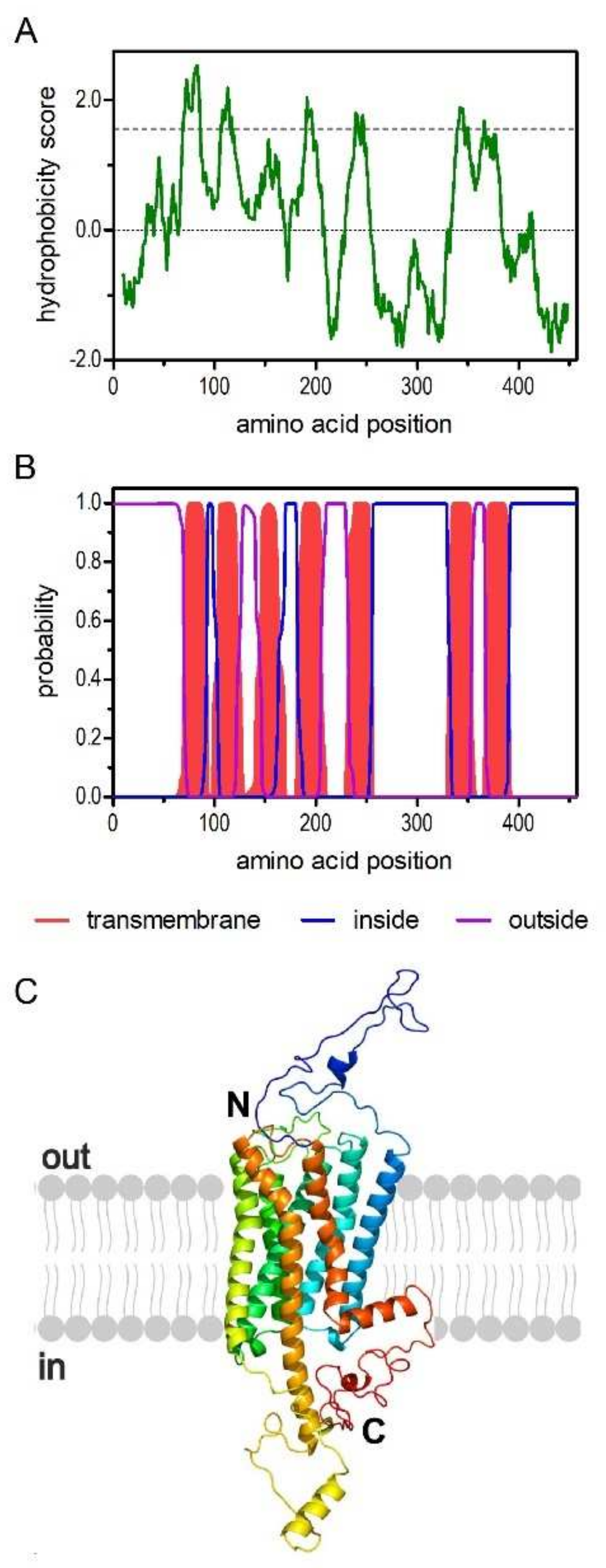
2.1. Molecular and Structural Properties of the PaOctβ2R
2.2. Annotation of P. americana Biogenic Amine Receptors and Phylogenetic Analysis of PaOctβ2R
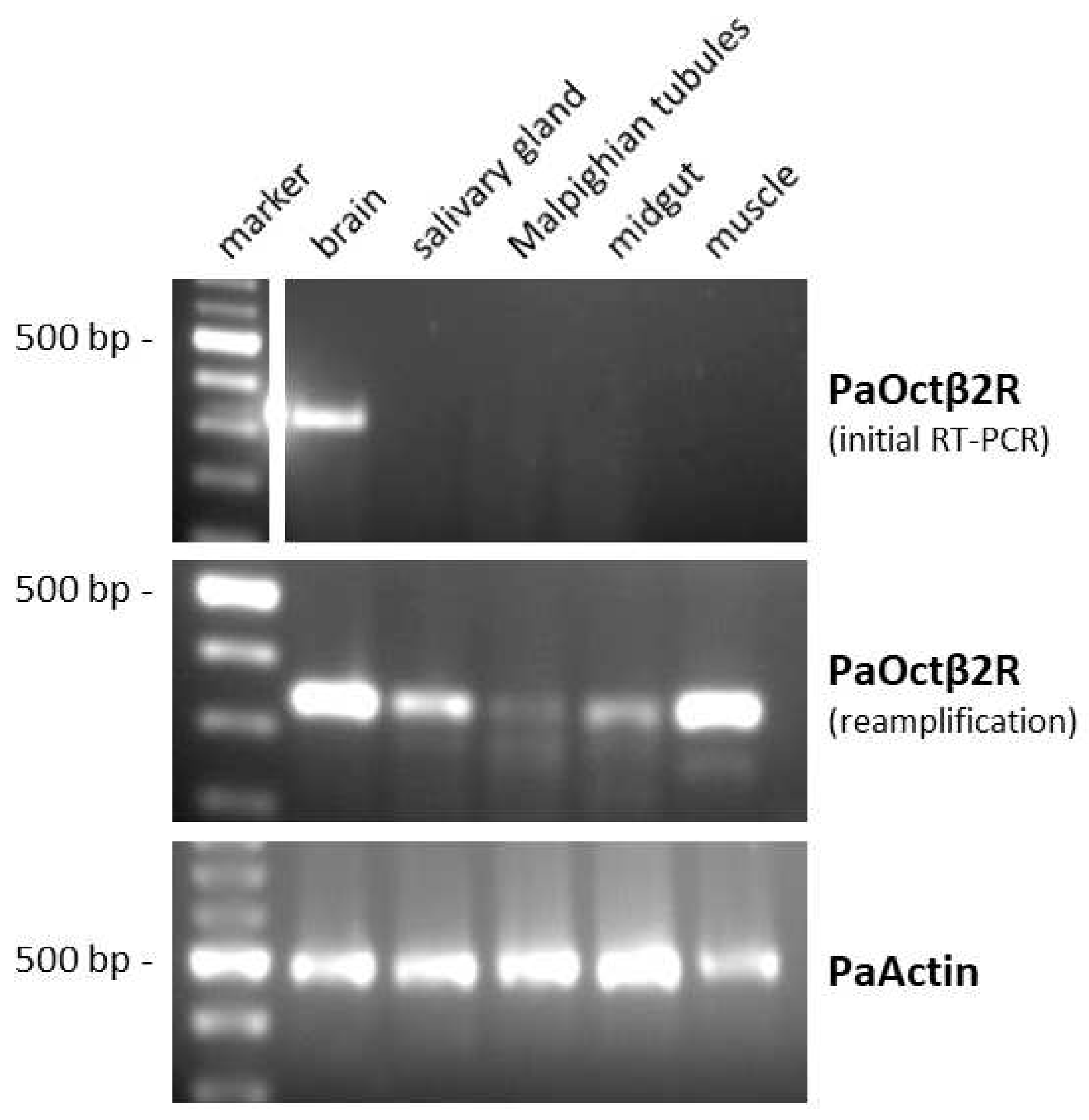
2.3. Tissue Distribution of PaOctβ2R mRNA
2.4. Expression of PaOctβ2R-HA in flpTM Cells
2.5. Ligand Specificity of the PaOctβ2R-HA Receptor
2.6. Pharmacological Properties of the PaOctβ2R-HA Receptor
3. Discussion
3.1. Structural Properties of the PaOctβ2R Protein and Phylogenetic Classification
3.2. Posttranslational Modification of PaOctβ2R
3.3. Pharmacological Properties of the PaOctβ2R Protein
3.4. Octopamine Receptors as Molecular Targets of Insecticides
4. Materials and Methods
4.1. Amplification of a Cockroach β-Adrenergic-Like Octopamine Receptor (PaOctβ2R) cDNA
4.2. Transcriptome Sequencing and De novo Assembly of Nucleotide Sequences
4.3. Multiple Sequence Alignment and Phylogenetic Analysis
4.4. RT-PCR Amplification of PaOctβ2R Fragments
4.5. Construction of PaOctβ2R-HA Expression Vector
4.6. Functional Expression of the PaOctβ2R-HA Receptor
4.7. Immunofluorescent Staining of Cell Lines
4.8. Western Blot Analysis
4.9. Functional Analysis of the PaOctβ2R-HA Receptor
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Verlinden, H.; Vleugels, R.; Marchal, E.; Badisco, L.; Pflüger, H.J.; Blenau, W.; Vanden Broeck, J. The role of octopamine in locusts and other arthropods. J. Insect Physiol. 2010, 56, 854–867. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Ozoe, Y. Molecular signalling, pharmacology, and physiology of octopamine and tyramine receptors as potential insect pest control targets. Adv. Insect Physiol. 2014, 46, 73–166. [Google Scholar]
- Blenau, W.; Baumann, A. Octopaminergic and tyraminergic signaling in the honeybee (Apis mellifera) brain: Behavioral, pharmacological, and molecular aspects. In Trace Amines and Neurological Disorders, 1st ed.; Farooqui, A., Ed.; Academic Press: Oxford, UK, 2016; pp. 203–220. [Google Scholar]
- Roeder, T. The control of metabolic traits by octopamine and tyramine in invertebrates. J. Exp. Biol. 2020, 223, jeb194282. [Google Scholar] [CrossRef] [PubMed]
- Hammer, M. An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees. Nature 1993, 366, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Schulz, D.J.; Robinson, G.E. Octopamine influences division of labor in honey bee colonies. J. Comp. Physiol. A 2001, 187, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Scheiner, R.; Baumann, A.; Blenau, W. Aminergic control and modulation of honeybee behaviour. Curr. Neuropharmacol. 2006, 4, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Schützler, N.; Girwert, C.; Hügli, I.; Mohana, G.; Roignant, J.Y.; Ryglewski, S.; Duch, C. Tyramine action on motoneuron excitability and adaptable tyramine/octopamine ratios adjust Drosophila locomotion to nutritional state. Proc. Natl. Acad. Sci. USA 2019, 116, 3805–3810. [Google Scholar] [CrossRef] [Green Version]
- Selcho, M.; Pauls, D. Linking physiological processes and feeding behaviors by octopamine. Curr. Opin. Insect Sci. 2019, 36, 125–130. [Google Scholar] [CrossRef]
- Walz, B.; Baumann, O.; Krach, C.; Baumann, A.; Blenau, W. The aminergic control of cockroach salivary glands. Arch. Insect Biochem. Physiol. 2006, 62, 141–152. [Google Scholar] [CrossRef]
- Watanabe, H.; Mizunami, M. Pavlov’s cockroach: Classical conditioning of salivation in an insect. PLoS ONE 2007, 2, e529. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.W.; Kim, J.H.; Pfeiffer, R.; Ahn, Y.J.; Page, T.L.; Kwon, H.W. Neuromodulation of olfactory sensitivity in the peripheral olfactory organs of the American cockroach, Periplaneta americana. PLoS ONE 2013, 8, e81361. [Google Scholar] [CrossRef]
- Saudou, F.; Amlaiky, N.; Plassat, J.L.; Borrelli, E.; Hen, R. Cloning and characterization of a Drosophila tyramine receptor. EMBO J. 1990, 9, 3611–3617. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Balfanz, S.; Baumann, A. Amtyr1: Characterization of a gene from honeybee (Apis mellifera) brain encoding a functional tyramine receptor. J. Neurochem. 2000, 74, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Mustard, J.A.; Kurshan, P.T.; Hamilton, I.S.; Blenau, W.; Mercer, A.R. Developmental expression of a tyramine receptor gene in the brain of the honey bee, Apis mellifera. J. Comp. Neurol. 2005, 483, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Cazzamali, G.; Klaerke, D.A.; Grimmelikhuijzen, C.J. A new family of insect tyramine receptors. Biochem. Biophys. Res. Commun. 2005, 338, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, A.; Roselli, G.; Evans, P.D. A comparison of the signalling properties of two tyramine receptors from Drosophila. J. Neurochem. 2013, 125, 37–48. [Google Scholar] [CrossRef]
- Reim, T.; Balfanz, S.; Baumann, A.; Blenau, W.; Thamm, M.; Scheiner, R. AmTAR2: Functional characterization of a honeybee tyramine receptor stimulating adenylyl cyclase activity. Insect Biochem. Mol. Biol. 2017, 80, 91–100. [Google Scholar] [CrossRef]
- Evans, P.D.; Maqueira, B. Insect octopamine receptors: A new classification scheme based on studies of cloned Drosophila G-protein coupled receptors. Invert. Neurosci. 2005, 5, 111–118. [Google Scholar] [CrossRef]
- Han, K.A.; Millar, N.S.; Davis, R.L. A novel octopamine receptor with preferential expression in Drosophila mushroom bodies. J. Neurosci. 1998, 18, 3650–3658. [Google Scholar] [CrossRef] [Green Version]
- Grohmann, L.; Blenau, W.; Erber, J.; Ebert, P.R.; Strünker, T.; Baumann, A. Molecular and functional characterization of an octopamine receptor from honeybee (Apis mellifera) brain. J. Neurochem. 2003, 86, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.X.; Xu, G.; Gu, G.X.; Mao, F.; Ye, G.Y.; Liu, W.; Huang, J. A new Drosophila octopamine receptor responds to serotonin. Insect Biochem. Mol. Biol. 2017, 90, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Wilms, J.A.; Balfanz, S.; Baumann, A. AmOctα2R: Functional characterization of a honeybee octopamine receptor inhibiting adenylyl cyclase activity. Int. J. Mol. Sci. 2020, 21, 9334. [Google Scholar] [CrossRef] [PubMed]
- Balfanz, S.; Strünker, T.; Frings, S.; Baumann, A. A family of octopamine receptors that specifically induce cyclic AMP production or Ca2+ release in Drosophila melanogaster. J. Neurochem. 2005, 93, 440–451. [Google Scholar] [CrossRef]
- Maqueira, B.; Chatwin, H.; Evans, P.D. Identification and characterization of a novel family of Drosophila β-adrenergic-like octopamine G-protein coupled receptors. J. Neurochem. 2005, 94, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Balfanz, S.; Jordan, N.; Langenstück, T.; Breuer, J.; Bergmeier, V.; Baumann, A. Molecular, pharmacological, and signaling properties of octopamine receptors from honeybee (Apis mellifera) brain. J. Neurochem. 2014, 129, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Troppmann, B.; Balfanz, S.; Krach, C.; Baumann, A.; Blenau, W. Characterization of an invertebrate-type dopamine receptor of the American cockroach, Periplaneta americana. Int. J. Mol. Sci. 2014, 15, 629–653. [Google Scholar] [CrossRef] [Green Version]
- Troppmann, B.; Balfanz, S.; Baumann, A.; Blenau, W. Inverse agonist and neutral antagonist actions of synthetic compounds at an insect 5-HT1 receptor. Br. J. Pharmacol. 2010, 159, 1450–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotte, C.; Krach, C.; Balfanz, S.; Baumann, A.; Walz, B.; Blenau, W. Molecular characterization and localization of the first tyramine receptor of the American cockroach (Periplaneta americana). Neuroscience 2009, 162, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Balfanz, S.; Baumann, A. PeaTAR1B: Characterization of a second type 1 tyramine receptor of the American cockroach, Periplaneta americana. Int. J. Mol. Sci. 2017, 18, 2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischof, L.J.; Enan, E.E. Cloning, expression and functional analysis of an octopamine receptor from Periplaneta americana. Insect Biochem. Mol. Biol. 2004, 34, 511–521. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, J.A.; Weinstein, H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 1995, 25, 366–428. [Google Scholar]
- Eilers, M.; Hornak, V.; Smith, S.O.; Konopka, J.B. Comparison of class a and dg protein-coupled receptors: Common features in structure and activation. Biochemistry 2005, 44, 8959–8975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ohta, H.; Ozoe, F.; Miyazawa, K.; Huang, J.; Ozoe, Y. Functional and pharmacological characterization of a β-adrenergic-like octopamine receptor from the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 476–486. [Google Scholar] [CrossRef]
- Hauser, F.; Cazzamali, G.; Williamson, M.; Blenau, W.; Grimmelikhuijzen, C.J. A review of neurohormone GPCRs present in the fruitfly Drosophila melanogaster and the honey bee Apis mellifera. Prog. Neurobiol. 2006, 80, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Bauknecht, P.; Jékely, G. Ancient coexistence of norepinephrine, tyramine, and octopamine signaling in bilaterians. BMC Biol. 2017, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Mizunami, M.; Matsumoto, Y. Roles of octopamine and dopamine neurons for mediating appetitive and aversive signals in Pavlovian conditioning in crickets. Front. Physiol. 2017, 8, 1027. [Google Scholar] [CrossRef] [Green Version]
- Selcho, M.; Pauls, D.; El Jundi, B.; Stocker, R.F.; Thum, A.S. The role of octopamine and tyramine in Drosophila larval locomotion. J. Comp. Neurol. 2012, 520, 3764–3785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, A.B.; Orchard, I. Biogenic Monoamines in the Control of Triatomine Physiology with Emphasis on Rhodnius prolixus. In Triatominae—The Biology of Chagas Disease Vectors, 1st ed.; Guarneri, A., Lorenzo, M., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 145–166. [Google Scholar]
- Stankiewicz, M.; Dąbrowski, M.; de Lima, M.E. Nervous system of Periplaneta americana cockroach as a model in toxinological studies: A short historical and actual view. J. Toxicol. 2012, 2012, 143740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayali, A.; Couzin-Fuchs, E.; David, I.; Gal, O.; Holmes, P.; Knebel, D. Sensory feedback in cockroach locomotion: Current knowledge and open questions. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2015, 201, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Fuscà, D.; Kloppenburg, P. Odor processing in the cockroach antennal lobe-the network components. Cell Tissue Res. 2021, 383, 59–73. [Google Scholar] [CrossRef]
- Congreve, M.; Langmead, C.; Marshall, F.H. The use of GPCR structures in drug design. Adv. Pharmacol. 2011, 62, 1–36. [Google Scholar]
- Kobilka, B.K. G protein coupled receptor structure and activation. Biochim. Biophys. Acta 2007, 1768, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Karageorgos, V.; Venihaki, M.; Sakellaris, S.; Pardalos, M.; Kontakis, G.; Matsoukas, M.T.; Gravanis, A.; Margioris, A.; Liapakis, G. Current understanding of the structure and function of family B GPCRs to design novel drugs. Hormones 2018, 17, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Bockaert, J.; Pin, J.P. Molecular tinkering of G protein-coupled receptors: An evolutionary success. EMBO J. 1999, 18, 1723–1729. [Google Scholar] [CrossRef] [Green Version]
- Jékely, G. Global view of the evolution and diversity of metazoan neuropeptide signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 8702–8707. [Google Scholar] [CrossRef] [Green Version]
- Elphick, M.R.; Mirabeau, O.; Larhammar, D. Evolution of neuropeptide signalling systems. J. Exp. Biol. 2018, 221, jeb151092. [Google Scholar] [CrossRef] [Green Version]
- O’Dowd, B.F.; Hnatowich, M.; Caron, M.G.; Lefkowitz, R.J.; Bouvier, M. Palmitoylation of the human β2-adrenergic receptor. Mutation of Cys341 in the carboxyl tail leads to an uncoupled nonpalmitoylated form of the receptor. J. Biol. Chem. 1989, 264, 7564–7569. [Google Scholar] [PubMed]
- Wu, S.F.; Jv, X.M.; Li, J.; Xu, G.J.; Cai, X.Y.; Gao, C.F. Pharmacological characterisation and functional roles for egg-laying of a β-adrenergic-like octopamine receptor in the brown planthopper Nilaparvata lugens. Insect Biochem. Mol. Biol. 2017, 87, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.L.; Guo, L.; Ma, H.H.; Hu, X.P.; Zhou, X.M. Phenyl imidazolidin-2-ones antagonize a β-adrenergic-like octopamine receptor in diamondback moth (Plutella xylostella). Pest. Manag. Sci. 2021, 77, 3224–3232. [Google Scholar] [CrossRef] [PubMed]
- Robidoux, J.; Martin, T.L.; Collins, S. β-adrenergic receptors and regulation of energy expenditure: A family affair. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 297–323. [Google Scholar] [CrossRef]
- Roeder, T.; Degen, J.; Gewecke, M. Epinastine, a highly specific antagonist of insect neuronal octopamine receptors. Eur. J. Pharmacol. 1998, 349, 171–177. [Google Scholar] [CrossRef]
- Rillich, J.; Stevenson, P.A. Winning fights induces hyperaggression via the action of the biogenic amine octopamine in crickets. PLoS ONE 2011, 6, e28891. [Google Scholar] [CrossRef] [Green Version]
- Minhas, N.; Gole, J.W.D.; Orr, G.L.; Downer, R.G.H. Pharmacology of [3H]mianserin binding in the nerve cord of the American cockroach, Periplaneta americana. Arch. Insect Biochem. Physiol. 1987, 6, 191–201. [Google Scholar] [CrossRef]
- Roeder, T. High-affinity antagonists of the locust neuronal octopamine receptor. Eur. J. Pharmacol. 1990, 191, 221–224. [Google Scholar] [CrossRef]
- Blenau, W.; Rademacher, E.; Baumann, A. Plant essential oils and formamidines as insecticides/acaricides: What are the molecular targets? Apidologie. 2012, 43, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system-a review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [Green Version]
- Ozoe, Y. Ion channels and G protein-coupled receptors as targets for invertebrate pest control: From past challenges to practical insecticides. Biosci. Biotechnol. Biochem. 2021, 85, 1563–1571. [Google Scholar] [CrossRef]
- Gole, J.W.; Orr, G.L.; Downer, R.G.H. Interaction of formamidines with octopamine-sensitive adenylate cyclase receptor in the nerve cord of Periplaneta americana L. Life Sci. 1983, 32, 2939–2947. [Google Scholar] [CrossRef]
- Downer, R.G.H.; Gole, J.W.D.; Orr, G.L. Interaction of formamidines with octopamine-, dopamine-, and 5-hydroxytryptamine-sensitive adenylate cyclase in the nerve cord of Periplaneta americana. Pestic. Sci. 1985, 16, 472–478. [Google Scholar] [CrossRef]
- Dudai, Y.; Buxbaum, J.; Corfas, G.; Ofarim, M. Formamidines interact with Drosophila receptors, alter the flies’ behavior and reduce their learning ability. J. Comp. Physiol. A 1987, 161, 739–746. [Google Scholar] [CrossRef]
- Ismail, S.M.M.; Baines, R.A.; Downer, R.G.H.; Dekeyser, M.A. Dihydrooxadiazines: Octopaminergic system as a potential site of insecticidal action. Pestic. Sci. 1996, 46, 163–170. [Google Scholar] [CrossRef]
- Enan, E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Finetti, L.; Tiedemann, L.; Zhang, X.; Civolani, S.; Bernacchia, G.; Roeder, T. Monoterpenes alter TAR1-driven physiology in Drosophila species. J. Exp. Biol. 2021, 224, jeb232116. [Google Scholar] [CrossRef]
- Kita, T.; Hayashi, T.; Ohtani, T.; Takao, H.; Takasu, H.; Liu, G.; Ohta, H.; Ozoe, F.; Ozoe, Y. Amitraz and its metabolite differentially activate α- and β-adrenergic-like octopamine receptors. Pest Manag. Sci. 2017, 73, 984–990. [Google Scholar] [CrossRef]
- Ohtani, A.; Arai, Y.; Ozoe, F.; Ohta, H.; Narusuye, K.; Huang, J.; Enomoto, K.; Kataoka, H.; Hirota, A.; Ozoe, Y. Molecular cloning and heterologous expression of an α-adrenergic-like octopamine receptor from the silkworm Bombyx mori. Insect Mol. Biol. 2006, 15, 763–772. [Google Scholar] [CrossRef]
- Takata, M.; Misato, S.; Ozoe, F.; Ozoe, Y. A point mutation in the β-adrenergic-like octopamine receptor: Possible association with amitraz resistance. Pest Manag. Sci. 2020, 76, 3720–3728. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N.N.; Hope, M. Progress in the epidemiology and diagnosis of amitraz resistance in the cattle tick Boophilus microplus. Vet. Parasitol. 2007, 146, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N.N.; Klafke, G.; Corley, S.W.; Tidwell, J.; Berry, C.M.; Koh-Tan, H.C. Molecular biology of amitraz resistance in cattle ticks of the genus Rhipicephalus. Front. Biosci. (Landmark Ed.) 2018, 23, 796–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S96–S119. [Google Scholar] [CrossRef]
- Rinkevich, F.D. Detection of amitraz resistance and reduced treatment efficacy in the Varroa Mite, Varroa destructor, within commercial beekeeping operations. PLoS ONE 2020, 15, e0227264. [Google Scholar] [CrossRef] [Green Version]
- Corley, S.W.; Jonsson, N.N.; Piper, E.K.; Cutullé, C.; Stear, M.J.; Seddon, J.M. Mutation in the RmbetaAOR gene is associated with amitraz resistance in the cattle tick Rhipicephalus microplus. Proc. Natl. Acad. Sci. USA 2013, 110, 16772–16777. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Fan, X.Y.; Qiao, X.; Montell, C.; Huang, J. An octopamine receptor confers selective toxicity of amitraz on honeybees and Varroa mites. Elife 2021, 10, e68268. [Google Scholar] [CrossRef]
- García, F.; Notario, M.J.; Cabanás, J.M.; Jordano, R.; Medina, L.M. Incidence of bacteria of public health interest carried by cockroaches in different food-related environments. J. Med. Entomol. 2012, 49, 1481–1484. [Google Scholar] [CrossRef]
- Arruda, L.K.; Vailes, L.D.; Ferriani, V.P.; Santos, A.B.; Pomés, A.; Chapman, M.D. Cockroach allergens and asthma. J. Allergy Clin. Immunol. 2001, 107, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Sookrung, N.; Chaicumpa, W. A revisit to cockroach allergens. Asian Pac. J. Allergy Immunol. 2010, 28, 95–106. [Google Scholar]
- Nasirian, H.; Salehzadeh, A. Control of cockroaches (Blattaria) in sewers: A practical approach aystematic review. J. Med. Entomol. 2019, 56, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Baumann, A. Molecular characterization of the ebony gene from the American cockroach, Periplaneta americana. Arch. Insect Biochem. Physiol. 2005, 59, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef] [PubMed]
- Ragionieri, L.; Özbagci, B.; Neupert, S.; Salts, Y.; Davidovitch, M.; Altstein, M.; Predel, R. Identification of mature peptides from pban and capa genes of the moths Heliothis peltigera and Spodoptera littoralis. Peptides 2017, 94, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Bio. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, M. Compilation and analysis of sequences upstream from the translational start site in eukaryotic mRNAs. Nucleic Acids Res. 1984, 12, 857–872. [Google Scholar] [CrossRef]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis mellifera) and involvement of serotonin in phototactic behavior. Cell. Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef]
- Ludwig, J.; Margalit, T.; Eismann, E.; Lancet, D.; Kaupp, U.B. Primary structure of cAMP-gated channel from bovine olfactory epithelium. FEBS Lett. 1990, 270, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Okayama, H. High-efficiency transformation of mammalian cells by plasmid DNA. Mol. Cell. Biol. 1987, 7, 2745–2752. [Google Scholar]
- Wachten, S.; Schlenstedt, J.; Gauss, R.; Baumann, A. Molecular identification and functional characterization of an adenylyl cyclase from the honeybee. J. Neurochem. 2006, 96, 1580–1590. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Octopamine (n = 5) | Tyramine (n = 5) | |
|---|---|---|
| EC50 [M] | 4.67 × 10−10 | 4.30 × 10−8 |
| pEC50 | 9.35 ± 0.04 | 7.39 ± 0.04 |
| Substance | IC50 [M] | pIC50 | n | Maximal Inhibition [%] |
|---|---|---|---|---|
| epinastine | 1.2 × 10−8 | 7.944 ± 0.042 | 4 | 100 |
| mianserin | 1.0 × 10−7 | 7.006 ± 0.055 | 3 | 100 |
| phentolamine | 2.57 × 10−7 | 6.608 ± 0.069 | 4 | 100 |
| ketanserin | 5.12 × 10−6 | 5.291 ± 0.048 | 3 | 80 |
| 5-CT | n.d. | n.d. | 6 | 40 |
| yohimbine | - | - | 3 | - |
| 5-MT | - | - | 3 | - |
| 8-OH-DPAT | - | - | 3 | - |
| Substance | IC50 [M] | pIC50 | n | Maximal Inhibition [%] |
|---|---|---|---|---|
| epinastine | 1.3 × 10−8 | 7.951 ± 0.042 | 4 | 100 |
| mianserin | 1.16 × 10−7 | 6.938 ± 0.058 | 3 | 100 |
| phentolamine | 2.56 × 10−7 | 6.594 ± 0.022 | 4 | 100 |
| ketanserin | 1.68 × 10−6 | 5.766 ± 0.097 | 3 | 50 |
| 5-CT | n.d. | n.d. | 6 | 40 |
| yohimbine | - | - | 3 | - |
| 5-MT | - | - | 3 | - |
| 8-OH-DPAT | - | - | 3 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blenau, W.; Bremer, A.-S.; Schwietz, Y.; Friedrich, D.; Ragionieri, L.; Predel, R.; Balfanz, S.; Baumann, A. PaOctβ2R: Identification and Functional Characterization of an Octopamine Receptor Activating Adenylyl Cyclase Activity in the American Cockroach Periplaneta americana. Int. J. Mol. Sci. 2022, 23, 1677. https://doi.org/10.3390/ijms23031677
Blenau W, Bremer A-S, Schwietz Y, Friedrich D, Ragionieri L, Predel R, Balfanz S, Baumann A. PaOctβ2R: Identification and Functional Characterization of an Octopamine Receptor Activating Adenylyl Cyclase Activity in the American Cockroach Periplaneta americana. International Journal of Molecular Sciences. 2022; 23(3):1677. https://doi.org/10.3390/ijms23031677
Chicago/Turabian StyleBlenau, Wolfgang, Anna-Sophie Bremer, Yannik Schwietz, Daniel Friedrich, Lapo Ragionieri, Reinhard Predel, Sabine Balfanz, and Arnd Baumann. 2022. "PaOctβ2R: Identification and Functional Characterization of an Octopamine Receptor Activating Adenylyl Cyclase Activity in the American Cockroach Periplaneta americana" International Journal of Molecular Sciences 23, no. 3: 1677. https://doi.org/10.3390/ijms23031677