
Oxidative Stress Modulation by Carnosine in Scaffold Free Human Dermis Spheroids Model: A Proteomic Study

 , , ,
, , , 
Abstract
:1. Introduction
2. Results and Discussion
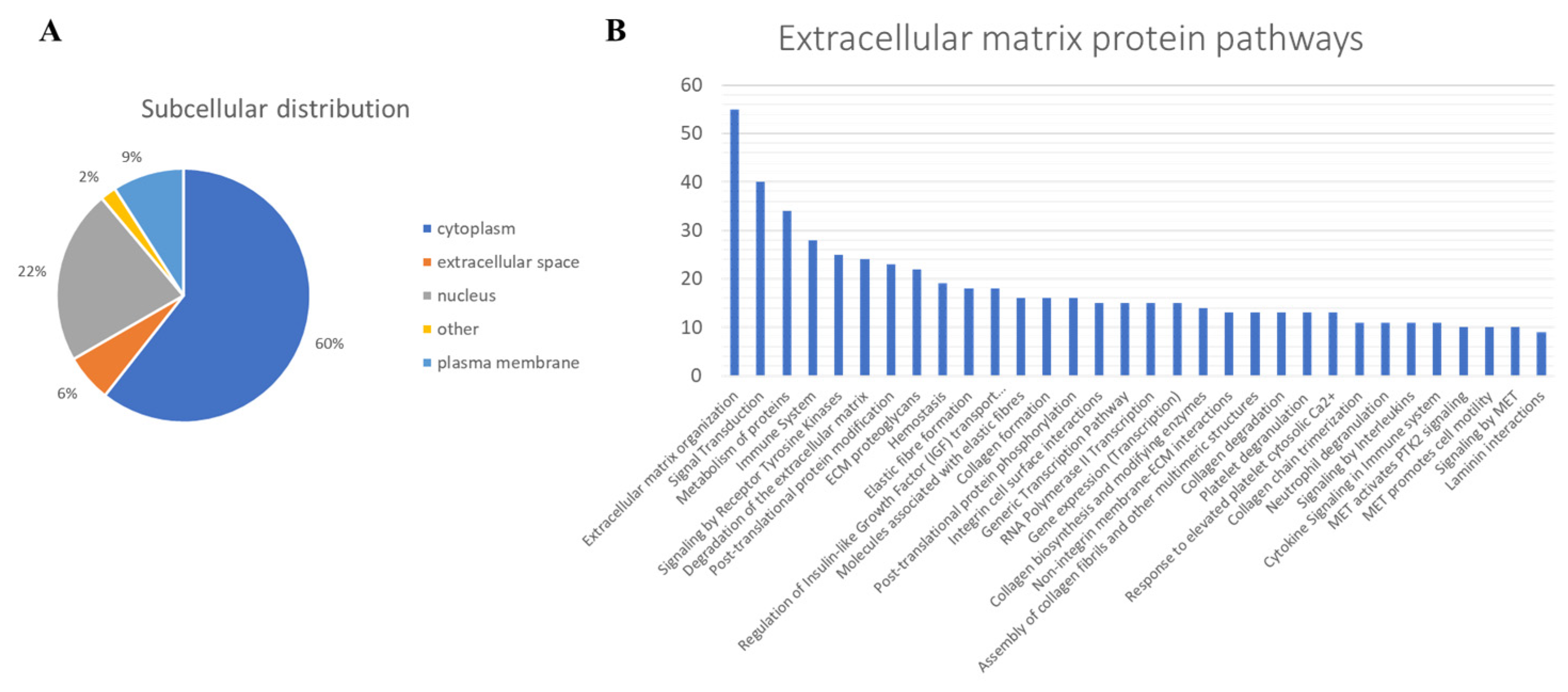
2.1. Protein Profiling of Human Dermis Spheroids Model
2.2. Differential Regulation of Protein Pathways by L-Carnosine in Dermis Spheroids
3. Materials and Methods
3.1. Cell Source and Culture
3.2. Dermis Spheroid Production and L-Carnosine Treatment
3.3. Immunofluorescence for Collagen Type III on 3D Whole Mount Samples and Signal Quantification
3.4. Protein Extraction and in-Solution Trypsin Digestion
3.5. High-Resolution LC-MS/MS Analysis and Data Elaboration
3.6. Protein Network Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Larroque-Cardoso, P.; Camare, C.; Nadal-Wollbold, F.; Grazide, M.H.; Pucelle, M.; Garoby-Salom, S.; Bogdanowicz, P.; Josse, G.; Schmitt, A.M.; Uchida, K.; et al. Elastin modification by 4-hydroxynonenal in hairless mice exposed to uv-a. Role in photoaging and actinic elastosis. J. Investig. Dermatol. 2015, 135, 1873–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldini, G.; de Courten, B.; Regazzoni, L.; Gilardoni, E.; Ferrario, G.; Baron, G.; Altomare, A.; D’Amato, A.; Vistoli, G.; Carini, M. Understanding the antioxidant and carbonyl sequestering activity of carnosine: Direct and indirect mechanisms. Free Radic. Res. 2021, 55, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Dissette, V.; Bignozzi, C.A.; Valacchi, G.; Pecorelli, A.; Manfredini, S.; Vertuani, S. Evaluation of the transepidermal penetration of a carnosine complex in gel formulation by 3d skin models. Cosmetics 2018, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Swiader, A.; Camare, C.; Guerby, P.; Salvayre, R.; Negre-Salvayre, A. 4-hydroxynonenal contributes to fibroblast senescence in skin photoaging evoked by uv-a radiation. Antioxidants 2021, 10, 365. [Google Scholar] [CrossRef]
- Aldini, G.; Carini, M.; Yeum, K.J.; Vistoli, G. Novel molecular approaches for improving enzymatic and nonenzymatic detoxification of 4-hydroxynonenal: Toward the discovery of a novel class of bioactive compounds. Free Radic. Biol. Med. 2014, 69, 145–156. [Google Scholar] [CrossRef]
- Radrezza, S.; Carini, M.; Baron, G.; Aldini, G.; Negre-Salvayre, A.; D’Amato, A. Study of carnosine’s effect on nude mice skin to prevent uv-a damage. Free Radic. Biol. Med. 2021, 173, 97–103. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Lo, L.M.; Raghunath, M.; Lee, K.K.H. Growing human dermal fibroblasts as spheroids renders them susceptible for early expression of pluripotency genes. Adv. Biosyst. 2019, 3, 94. [Google Scholar] [CrossRef]
- Rescigno, F.; Ceriotti, L.; Meloni, M. Extra cellular matrix deposition and assembly in dermis spheroids. Clin. Cosmet. Investig. Dermatol. 2021, 14, 935–943. [Google Scholar] [CrossRef]
- Oxlund, H.; Manschot, J.; Viidik, A. The role of elastin in the mechanical-properties of skin. J. Biomech. 1988, 21, 213–218. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielty, C.M.; Whittaker, S.P.; Grant, M.E.; Shuttleworth, C.A. Type-vi collagen microfibrils: Evidence for a structural association with hyaluronan. J. Cell Biol. 1992, 118, 979–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karsdal, M.A.; Sun, S. Chapter 6-Type VI collagen. In Biochemistry of Collagens, Laminins and Elastin: Structure, Function and Biomarkers; Academic Press: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Leeming, D.J.; Nielsen, S.H.; Vongsuvanh, R.; Uchila, P.; Nielsen, M.J.; Reese-Petersen, A.L.; van der Poorten, D.; Eslam, M.; Schuppan, D.; Karsdal, M.A.; et al. Endotrophin, a pro-peptide of type vi collagen, is a biomarker of survival in cirrhotic patients with hepatocellular carcinoma. Hepatic Oncol. 2021, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.K.; Hahn, R.A. Collagens. Cell Tissue Res. 2010, 339, 247–257. [Google Scholar] [CrossRef]
- Schiavinato, A.; Keene, D.R.; Wohl, A.P.; Corallo, D.; Colombatti, A.; Wagener, R.; Paulsson, M.; Bonaldo, P.; Sengle, G. Targeting of emilin-1 and emilin-2 to fibrillin microfibrils facilitates their incorporation into the extracellular matrix. J. Investig. Dermatol. 2016, 136, 1150–1160. [Google Scholar] [CrossRef] [Green Version]
- Adamo, C.S.; Zuk, A.V.; Sengle, G. The fibrillin microfibril/elastic fibre network: A critical extracellular supramolecular scaffold to balance skin homoeostasis. Exp. Dermatol. 2021, 30, 25–37. [Google Scholar] [CrossRef]
- Iacono, K.T.; Brown, A.L.; Greene, M.I.; Saouaf, S.J. Cd147 immunoglobulin superfamily receptor function and role in pathology. Exp. Mol. Pathol. 2007, 83, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Grass, G.D.; Toole, B.P. How, with whom and when: An overview of cd147-mediated regulatory networks influencing matrix metalloproteinase activity. Biosci. Rep. 2016, 36, e00283. [Google Scholar] [CrossRef] [Green Version]
- Bock-Marquette, I.; Saxena, A.; White, M.D.; DiMaio, J.M.; Srivastava, D. Thymosin beta 4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature 2004, 432, 466–472. [Google Scholar] [CrossRef]
- Fan, Y.; Gong, Y.Q.; Ghosh, P.K.; Graham, L.M.; Fox, P.L. Spatial coordination of actin polymerization and ilk-akt2 activity during endothelial cell migration. Dev. Cell 2009, 16, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Vitriol, E.A.; Shim, S.; Wise, A.L.; Velayutham, R.P.; Zheng, J.Q. Dynamic localization of g-actin during membrane protrusion in neuronal motility. Curr. Biol. 2013, 23, 1046–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, P.J.; Mancini, J.A.; Riendeau, D.; Ford-Hutchinson, A.W. Identification and characterization of a novel microsomal enzyme with glutathione-dependent transferase and peroxidase activities. J. Biol. Chem. 1997, 272, 22934–22939. [Google Scholar] [CrossRef] [Green Version]
- Bogaards, J.J.; Venekamp, J.C.; van Bladeren, P.J. Stereoselective conjugation of prostaglandin A2 and prostaglandin J2 with glutathione, catalyzed by the human glutathione S-transferases A1-1, A2-2, M1a-1a, and P1-1. Chem. Res. Toxicol. 1997, 10, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, Y.; Wang, H.; Liu, N.; Luo, L.; Zhao, C.; Zhou, D.; Tong, H.; Li, P.; Zhou, Q. Deficiency of MIF accentuates overloaded compres-sion-induced nucleus pulposus cell oxidative damage via depressing mitophagy. Oxid. Med. Cell. Longev. 2021, 2021, 6192498. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | p Value C7 vs. C14 | Log2 C7/C14 | p Value PD7 vs. C7 | Log2 PD7/C7 | Location |
|---|---|---|---|---|---|---|
| TF | transferrin | 1.35 × 10−07 | 3.98 | 8.78 × 10−06 | 2.67 | Extracellular Space |
| COL6A3 | collagen type VI alpha 3 chain | 1.16 × 10−03 | 4.71 | 2.64 × 10−03 | 2.28 | Extracellular Space |
| STOML2 | stomatin like 2 | 2.77 × 10−02 | 1.36 | 5.00 × 10−05 | 2.13 | Plasma Membrane |
| FBN1 | fibrillin 1 | 3.61 × 10−02 | 2.01 | 1.11 × 10−01 | 1.93 | Extracellular Space |
| INS | insulin | 1.42 × 10−02 | 1.53 | 4.84 × 10−04 | 1.56 | Extracellular Space |
| SLC1A5 | solute carrier family 1 member 5 | 7.57 × 10−04 | 1.54 | 7.41 × 10−04 | 1.46 | Plasma Membrane |
| TPBG | trophoblast glycoprotein | 2.28 × 10−03 | 1.20 | 5.37 × 10−04 | 1.45 | Plasma Membrane |
| MFAP2 | microfibril associated protein 2 | 1.31 × 10−02 | 1.37 | 8.77 × 10−03 | 1.42 | Extracellular Space |
| GPNMB | glycoprotein nmb | 1.88 × 10−02 | 1.17 | 1.45 × 10−02 | 1.36 | Plasma Membrane |
| MIF | macrophage migration inhibitory factor | 2.26 × 10−02 | −0.49 | 5.21 × 10−03 | 0.94 | Extracellular Space |
| RAP2B | RAP2B. member of RAS oncogene family | 7.48 × 10−03 | 0.94 | 3.60 × 10−03 | 0.93 | Plasma Membrane |
| MME | membrane metalloendopeptidase | 5.19 × 10−02 | 0.76 | 5.59 × 10−02 | 0.89 | Plasma Membrane |
| CCDC47 | coiled-coil domain containing 47 | 7.94 × 10−02 | 0.91 | 4.60 × 10−02 | 0.72 | Extracellular Space |
| LAMTOR1 | late endosomal/lysosomal adaptor. MAPK and MTOR activator 1 | 1.78 × 10−02 | 1.31 | 1.46 × 10−01 | 0.70 | Plasma Membrane |
| BSG | basigin (Ok blood group) | 8.96 × 10−01 | 0.03 | 5.41 × 10−04 | 0.69 | Plasma Membrane |
| EMILIN2 | elastin microfibril interfacer 2 | 2.38 × 10−03 | 1.33 | 2.57 × 10−02 | 0.69 | Extracellular Space |
| MANF | mesencephalic astrocyte derived neurotrophic factor | 8.26 × 10−02 | 0.32 | 7.10 × 10−03 | 0.68 | Extracellular Space |
| ITGAV | integrin subunit alpha V | 6.55 × 10−03 | 0.50 | 2.10 × 10−02 | 0.63 | Plasma Membrane |
| LOXL1 | lysyl oxidase like 1 | 8.17 × 10−01 | 0.05 | 2.39 × 10−03 | 0.62 | Extracellular Space |
| CD248 | CD248 molecule | 3.71 × 10−03 | 0.81 | 6.14 × 10−02 | 0.62 | Plasma Membrane |
| SCARB2 | scavenger receptor class B member 2 | 3.63 × 10−03 | 1.10 | 1.65 × 10−01 | 0.60 | Plasma Membrane |
| NT5E | 5’-nucleotidase ecto | 3.15 × 10−05 | 1.10 | 9.54 × 10−02 | 0.59 | Plasma Membrane |
| ARL8B | ADP ribosylation factor like GTPase 8B | 9.04 × 10−02 | 0.66 | 1.07 × 10−01 | 0.57 | Plasma Membrane |
| THY1 | Thy-1 cell surface antigen | 4.21 × 10−04 | 1.30 | 1.61 × 10−01 | 0.55 | Plasma Membrane |
| COL6A2 | collagen type VI alpha 2 chain | 1.60 × 10−02 | 0.85 | 1.22 × 10−01 | 0.52 | Extracellular Space |
| LAMP1 | lysosomal associated membrane protein 1 | 2.76 × 10−01 | 0.36 | 5.61 × 10−02 | 0.52 | Plasma Membrane |
| ESYT2 | extended synaptotagmin 2 | 2.21 × 10−02 | 0.66 | 1.22 × 10−01 | 0.50 | Plasma Membrane |
| CD81 | CD81 molecule | 4.55 × 10−03 | 0.88 | 1.43 × 10−01 | 0.46 | Plasma Membrane |
| CD44 | CD44 molecule (Indian blood group) | 1.80 × 10−02 | 0.82 | 2.69 × 10−01 | 0.44 | Plasma Membrane |
| EMILIN1 | elastin microfibril interfacer 1 | 1.91 × 10−02 | 0.72 | 5.22 × 10−02 | 0.36 | Extracellular Space |
| HLA-A | major histocompatibility complex. class I. A | 3.99 × 10−03 | 0.89 | 2.65 × 10−01 | 0.35 | Plasma Membrane |
| MARCKS | myristoylated alanine rich protein kinase C substrate | 9.03 × 10−02 | −0.26 | 9.51 × 10−04 | −0.77 | Plasma Membrane |
| LAMP2 | lysosomal associated membrane protein 2 | 7.14 × 10−01 | 0.07 | 1.63 × 10−02 | −0.77 | Plasma Membrane |
| PLPP3 | phospholipid phosphatase 3 | 8.12 × 10−01 | 0.03 | 2.23 × 10−02 | −0.85 | Plasma Membrane |
| FN1 | fibronectin 1 | 2.20 × 10−09 | −0.84 | 2.50 × 10−01 | −1.46 | Extracellular Space |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, G.; Rescigno, F.; Meloni, M.; Baron, G.; Aldini, G.; Carini, M.; D’Amato, A. Oxidative Stress Modulation by Carnosine in Scaffold Free Human Dermis Spheroids Model: A Proteomic Study. Int. J. Mol. Sci. 2022, 23, 1468. https://doi.org/10.3390/ijms23031468
Aiello G, Rescigno F, Meloni M, Baron G, Aldini G, Carini M, D’Amato A. Oxidative Stress Modulation by Carnosine in Scaffold Free Human Dermis Spheroids Model: A Proteomic Study. International Journal of Molecular Sciences. 2022; 23(3):1468. https://doi.org/10.3390/ijms23031468
Chicago/Turabian StyleAiello, Gilda, Francesca Rescigno, Marisa Meloni, Giovanna Baron, Giancarlo Aldini, Marina Carini, and Alfonsina D’Amato. 2022. "Oxidative Stress Modulation by Carnosine in Scaffold Free Human Dermis Spheroids Model: A Proteomic Study" International Journal of Molecular Sciences 23, no. 3: 1468. https://doi.org/10.3390/ijms23031468