1. Introduction
Long-lasting protective immunity against a pathogen, in a host, is most often achieved through the development of pathogen-specific immune memory T cells that can be restimulated during the early phase of the infection. The functional properties of such memory T cells are orchestrated by cellular and molecular regulators, which define their phenotype and might also regulate their plasticity (i.e., the extent to which their phenotype can still be influenced by the pathogen) [
1]. The functional characteristics of memory T cells such as their plasticity, their ability to be recalled, and the duration of their persistence have significant implications for vaccination strategies. Indeed, for vaccines to be effective the immune memory needs to be resilent to manipulation by the pathogen during infection, so that the host is protected from pathogen-mediated manipulation of the burgeoning immune response, and persistence and recall functions of the immune response is not compromised during infection [
1].
The complex biology of parasites has often, over evolutionary timeframes, allowed them to develop specific mechanisms to manipulate the host’s immune system in order survive long-term. For example,
S. mansoni secretes immunomodulatory molecules able to impact DC functions [
2] and affect the type of T helper responses by inducing nonspecific suppressive pathways during different life stages [
3]. Indeed,
S. mansoni secretory excretory antigens (SEA), produced in the egg-stage of the parasite, have the ability to manipulate human or mouse DCs to suppress classical maturation of co-stimulatory ligand expression. These manipulated DCs induce a Th2 response in vitro following a transfer to a naïve host under MyD88
- and IL-4-independent conditions [
4]. Molecules having an effect on this pathway have been identified, including
S. mansoni Omega-1, resulting in the induction of a Th2 type immune response [
5,
6]. While the mechanisms of T cell manipulation by
S. mansoni eggs are well understood for naïve T cells, it is unclear whether this manipulation takes place against fully differentiated immune memory T cells with an established T helper phenotype bias. In turn, this understanding of whether parasites are able to influence an established immune memory response during the recall phase is critical to vaccination strategies relying on immune memory T cells able to withstand pathogen manipulation.
In this study, we utilise a model based on OT-II cells and transgenic
S. mansoni parasites to investigate whether fully differentiated Th1, Th2 and Th17 immune memory cells can withstand immune manipulation by
S. mansoni parasites following in vivo challenge. This work may also have important implications for the use of parasite-derived molecules as therapeutic agents [
7,
8,
9,
10].
2. Materials and Methods
2.1. Experimental Mice
Six- to eight-week-old female BALB/c and OT-II mice were purchased from the Walter and Eliza Hall Institute of Medical Research (WEHI, Melbourne, Australia) and were used in the present study to maintain the life cycle of S. mansoni and as a source of naïve OT-II T cells, respectively. All the experiments related to mice were conducted subject to approval from the Animal Ethical Committee of the University of Melbourne (Ethics ID: 1312952).
2.2. Transduction of S. mansoni Eggs Using Lentivirus Vector
We followed the egg-transduction method of Hagen et al. [
11]. Depending on the purpose, a range of 8000 to 12,000 eggs were used for viral transfection and untranduced (wild type) eggs were also included as a control. The eggs were cultured with 500 μL complete DMEM in the presence of 100 μL virus particles at a titter of 25 × 10
3 pfu per ml of OVA-encoding pGIPZ lentiviral vector containing CMV promoter or left untreated in complete DMEM. The OVA expressing construct was designed so that parasite-expressed OVA has an identical protein sequence as native OVA [
12]. The OVA transduced parasite eggs where previously characterized extensively and were shown to stimulate OT-II cells [
12].
2.3. Cell Culture
Spleen, maxillary and inguinal lymph nodes of BALB/c and OT-II mice were collected and pooled in RPMI-1640 (Invitrogen, Life Technologies, Waltham, MA, USA). The tissues were crushed using forceps and filtered through a 70 µm nylon cell strainer. The cells were centrifuged at 276× g for 6 min. After aspirating the supernatants, the red blood cell pellets were lysed using distilled water for 9 s, followed by addition of 10% of 10× PBS to stop the lysis reaction. The cell suspension was centrifuged at 276× g for 6 min. Supernatants were removed, and the cells were resuspended in fresh complete RPMI-1640 medium ((Invitrogen, Life Technologies) supplemented with 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin and 10% v/v heat-inactivated FCS, and 50 μM 2-mercaptoethanol). When required, cell concentrations were estimated using a hemocytometer. In all experiments we used pools of cells prepared from spleen and lymph nodes.
2.4. In Vitro Polarization of OT-II T Cells
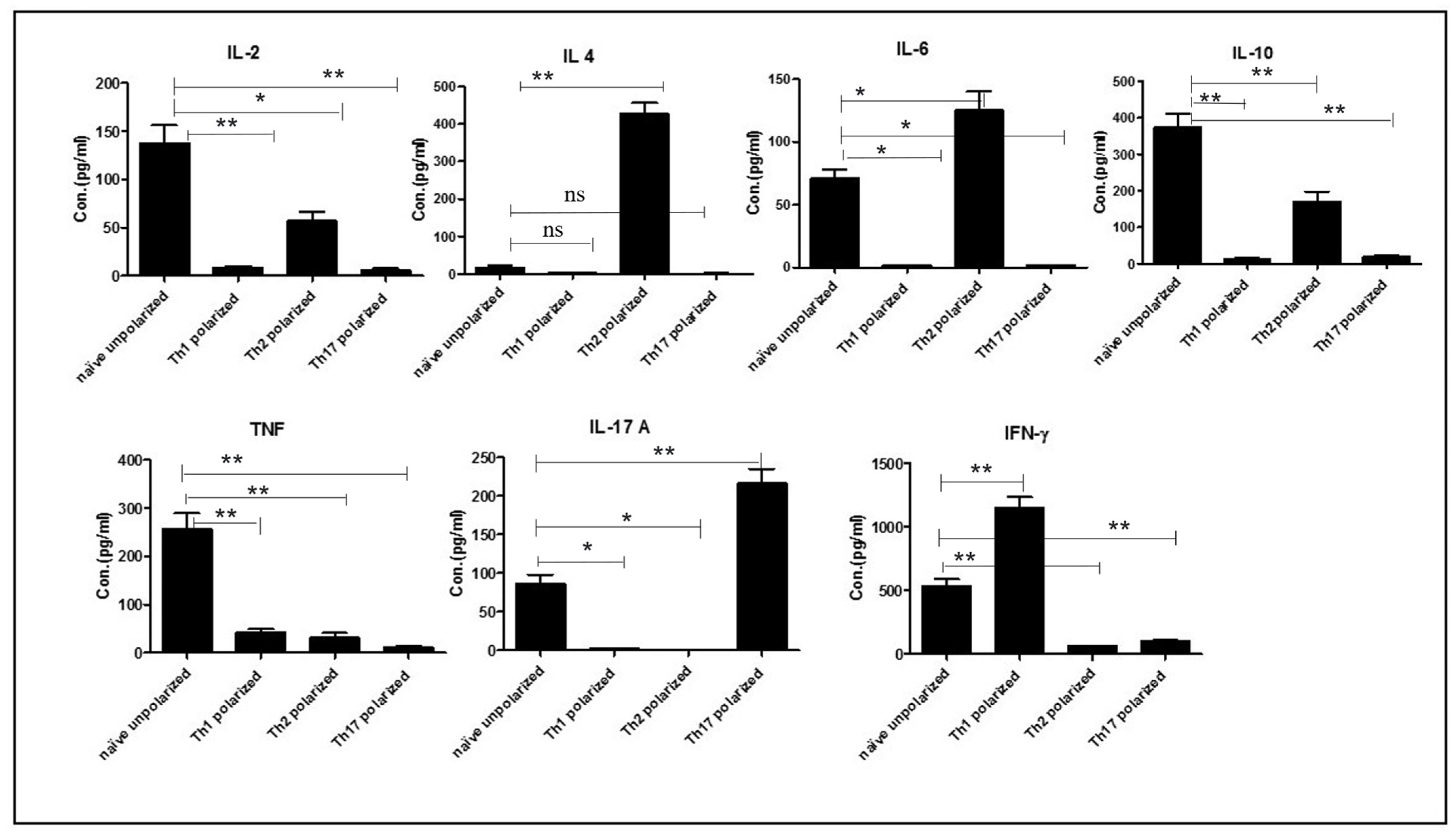
Naïve OT-II cells prepared as above, were polarized into three lineages using differentiating cytokines as follows: 2 × 105 cells per well were cultured for 4 days and polarized into Th1 (using 10 μg/mL OVA peptide, 10 ng/mL of IL-2), Th2 (using 10 μg OVA peptide and 10 ng/mL of IL-2, 10 ng/mL of IL-4 and 10 μg/mL of anti-IFN-γ), and Th17 (using 10 μg/mL OVA peptide, 10 μg/mL of anti-IL-4, 10 μg/mL of anti-IFN-γ antibodies, 5 ng/mL of anti-TGFβ and 20 ng/mL of IL-6). Naïve cells were stimulated with 10 μg/mL of OVA peptide only, to get unpolarized cells. Cells were further stimulated for two days using 4.5 µg/mL of IL-7 (PeproTech, Rocky Hill, NJ, USA) to generate memory cells. Th-polarised cells, 105, per well were stimulated with OVA peptide (10 μg/mL) for 72 h. The degree of polarization was determined based on the cytokine signatures of the differentiated cells using the mouse Th1, Th2, and Th17 cytometric bead array (BD Bioscience, North Brunswick Township, NJ, USA) as per manufacturer’s instructions.
2.5. Adoptive Transfer of OT-II T Cells to Congenic Mice and Parasite Injection
A total of 2 × 106 carboxyfluorescein-diacetate-succinimidyl-ester (CFSE; Bio Legend, San Diego, CA, USA) labelled OT-II T cells, which express CD45.2, were adoptively transferred into CD45.1 congenic mice. After 48 h, mice were injected with different antigens depending on the purpose of the experiment, including 1000 transduced or untransduced S. mansoni eggs suspended in 50 µL of PBS through intravenously (IV), or 20 µg/mL of OVA peptide as a positive control through the same route. Similarly, unchallenged OT-II T cells were used as a negative control. After 7 days, mice were culled to extract OT-II T cells from spleen for further analysis.
2.6. T Cell Proliferation Assay
Spleen, maxillary, axillary and inguinal lymph nodes of OT-II mice were collected and pooled for cell extraction using complete RPMI-1640 medium (Invitrogen, Life Technologies). Single cell suspensions were obtained by passing the tissue through a 70 µm nylon cell strainer. Red blood cells were lysed by water and lysis was stopped using 10× PBS, as described above. Cells were then resuspended in complete RPMI-1640 medium. For proliferation assays, cells were stained with CFSE according to manufacturer’s instruction. A total of 2 × 105 labelled cells per well were co-incubated with 25, 50, 100, or 200 transduced or un-transduced S. mansoni eggs for 4–5 days in 96 well plates at 37 °C, 5% CO2. After four days culturing, OT-II cells were collected and washed with FACS buffer (10% FCS in PBS). For proliferation analysis, cells were stained with PE-anti-mouse CD4 (Bio Legend) following an FcγR-surface block (anti-FcγR, clone 2.4G2-16, WEHI Monoclonal Facility). To exclude dead cells, 7-amino-actinomycin D (1 µg/mL; Sigma Aldrich, Steinheim, Germany) was added to cell suspension prior data acquisition using The BD FACSVerse™ flow cytometer (BD Bioscience). The data was analysed using flowjo version 10 (BD Bioscience, Ashland, OR, USA) gating on CD4+ T cells having CFSElow phenotypes. In the standard technique for cell proliferation the presence of low levels of CFSE indicate that the cells have proliferated since the dye has been diluted during consecutive cell divisions. The results were expressed as % CFSElow cells in the total CD4 population.
2.7. Cytokine Assay
Cell culture supernatants from T cell proliferation assays were analysed for cytokine production by mouse Th1, Th2, and Th17 cytometric bead array (BD Bioscience) as per the manufacturer’s instructions. According to the manufacturer, the detection limits for each cytokine were: IL-2 (0.1 pg/mL), IL-4 (0.03 pg/mL), IL-6 (1.4 pg/mL), IFN-γ (0.5 pg/mL), TNF (0.9 pg/mL), IL-17 A (0.8 pg/mL) and IL-10 (16.8 pg/mL).
2.8. Statistical Analysis
Data was computed and analysed using the single cell analysis software: flowjo version 10. All statistical analyses were performed using one-way ANOVA followed by t-test using Graph Pad Prism version 7.00 for Windows (GraphPad Software, La Jolla, CA, USA). In all the analyses, the confidence level was held at 95% and p < 0.05 was required for significance.
4. Discussion
Following infection of a vaccinated animal with a pathogen possessing immunomodulatory properties, the quality of the immune memory induced by the vaccine and how it can be affected during a recall response is very important to the disease outcome. Indeed, parasites develop specific tactics to subvert the recall of an immune response, which influences the effectiveness of vaccination. The functional persistence and recall of memory CD4
+ T cells allow the adjustment of the host’s immune response continuously, following exposure to different pathogenic infections [
1]. To this end, understanding the development of immune memory resilience by Th polarised cells in the face of immune modulatory pathogens is very important for the development of effective vaccination strategies towards these immune modulatory pathogens.
In this study, we first demonstrated that OT-II cells could be fully differentiated into Th1, Th2 and Th17 cells since these cells produced their signature cytokines following in vitro treatment.
Previous finding suggest that a predominant Th1 response is induced in vitro following exposure to
S. mansoni eggs. This result appears in contradiction with the observation that
S. mansoni eggs induce a Th2 response during in vivo infection. One possible explanation for this discrepancy is that not all cells that were implicated in the induction of the expected Th2 response in vivo are present in vitro. For example, it is possible that some of the critical DC populations might not be present in sufficient numbers in the OT-II-based system. In addition, the OT-II cells can be stimulated not only by B cells but also by a range of other APCs present in cell cultures [
12]. Another possibility is that OT-II cells are intrinsically biased towards Th1 responses. This has been suggested previously [
13], at least when the OT-II cells are stimulated with B cells as APCs.
We subsequently demonstrated that these in vitro polarised memory Th cells can recognize OVA protein expressed by
S. mansoni eggs and can respond by both proliferating and secreting specific cytokines such as IFN-γ, IL-17A, TNF, IL-10 and IL-6. Although we did not explicitly test whether OVA produced by the transduced parasites is glycosylated, we previously showed that it had an identical molecular weight to native OVA [
12] and the fact that it is recognized by OT-II cells suggests that it is processed by APCs in a similar way to native OVA. We also showed that OVA protein expressed by
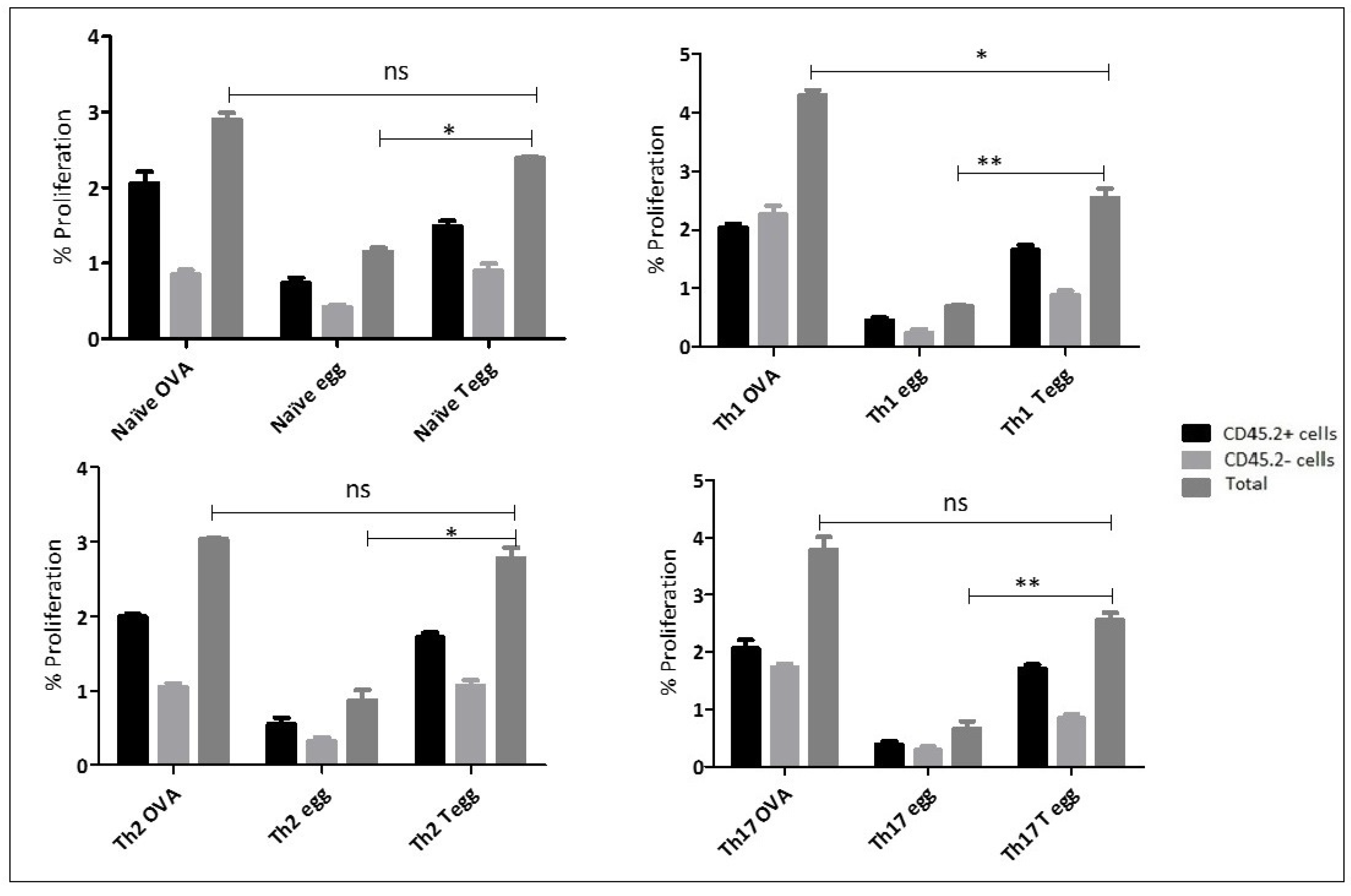
S. mansoni parasite eggs is recognized by polarised T helper cells in vivo. Indeed, these cells proliferate significantly more in vivo when re-stimulated with OVA-transduced eggs compared to untransduced eggs. The only exception was Th2-polarised cells, which do appear to have a high background level of proliferation even when only stimulated with untransduced eggs and as a result, there is no significant difference with between the proliferation induced by transduced and untransduced eggs. The reason for this elevated background proliferation is currently unknown.
In the present study, under both in vivo and ex vivo stimulation conditions, memory T cells showed a significant proliferative response to OVA expressing
S. mansoni eggs suggesting that they are able to not only survive but also continues to produce their cytokine signatures in the presence of the OVA antigen for extended periods of time. This is important, because CD4
+ T cells require a prolonged exposure to the antigen for their differentiation into effector cells and memory development and survival [
14,
15,
16]. Despite the long-term survival of memory cells in the presence of IL-7, there is a specific difference in the mechanism of their survival [
17]. For example, engagement of MHC class II and T cell receptor signalling have implications in the functional capacities and long-term survival of memory CD4
+ T cells [
1]. The functional capacity of these memory CD4
+ T cells is not irreversibly committed but it can be activated differentially during the recall immune response by changing the nature of the cytokine mediated signals present during the recall phase of the response [
18]. Similarly, the pattern of gene expression by memory Th1 and Th2 cells have been found to develop functional flexibility. For example, Th1 and Th2 were cloned under the same or opposite polarising conditions and under the same polarising conditions, cells retained the capacity to produce originally imprinted cytokines, but when restimulated under opposite polarising conditions, cells produce alternative cytokines.
The capacity of T helper memory cells to retain the ability to produce their cytokine signatures was confirmed, and this study showed that immune memory resilience can indeed be measured in this way, and that the in vitro generated polarised Th cells remain unaffected by the parasite eggs. This despite the fact that
S. mansoni eggs are well known for their ability to modulate the immune response towards a Th2 response following infection. The present finding is consistent with previous studies which showed that Th polarised cells can develop some degree of resilience to manipulation [
19,
20]. However, such studies were performed under in vitro conditions and it can therefore not be excluded that other in vivo controlling factors such as secretion of effector cytokines which have a regulatory effect in relation to the differentiation of CD4
+ T memory cells under in vivo conditions. More importantly, in the case of such parasites which causes complex immune responses, an in vivo study is more important and valid than in vitro studies because the presence of DCs which plays a significant role in the initiation and polarisation of T cells responses and can be affected by the presence of
S. mansoni eggs [
21]. These DCs and the microenvironment they are in, might not be present under in vitro conditions.
The cytokine environment at the time of primary stimulation can alter the outcome of memory CD4
+ T cell responses such as either promoting Th1 memory CD4
+ T cells with proinflammatory responses or Th2 type memory CD4
+ T cell responses [
20,
22]. Until the discovery of two other CD4
+ T cell lineages, Th17, and Treg cells, the differentiation of naïve CD4
+ T cells into specific lineages with specific effector function was considered an irreversible endpoints [
23]. In contrast to the Th1 and Th2 phenotypes the Th17 cells are less stable and this leads to a higher degree of flexibility during their differentiation and this phenomenon has its own biological implication [
23]. Indeed, the flexibility of these cells in terms of commitment to a particular phenotype might lead to lack of immune memory resilience in the face of an immunomodulatory pathogen. For example, it was reported that there is a cross-regulation between Th1, Th2, and Th17 during
S. mansoni infection, with Th1 cytokines (such as IFN-γ), and Th2 cytokines (such as IL-4) inhibiting the production of Th17 cytokines (such as IL-17) [
24].
It is well-established that following the acute phase of infection or during the first six weeks of infection, the immune response to
S. mansoni is predominantly Th1, characterised by the abundant production of TNF-α, and IFN-γ [
25]. However, soon after these 6 weeks, the parasite starts laying eggs, and the disease progresses into different organs mainly the liver and caused granulomas due to the immune response induced by the deposited
S. mansoni eggs. At this stage Th2 immune response to egg antigens predominates and T cells produce high level of IL-4, IL-5, IL-13, and IL-10 [
26]. The host-parasite interaction is very complex as they can involve both immunomodulatory mechanisms from the parasite and countermeasures from the host to resist these immunomodulatory effects. Following differentiation of naïve T cells in vitro into Th1, Th2, and Th17 phenotypes, we tested the degree of plasticity and resilience in the face of a parasite challenge after the polarised cells were transferred into recipient mice for in vivo study. The results showed that, Th1, Th2, and Th17 memory cells continued to produce their signature cytokines, when challenged by the parasite, suggesting that the parasite eggs were unable to manipulate the immune response of the differentiated T cells. At first sight this seems to contradict the well-established observations that the initial Th1 response against the parasite during the acute phase morphs into a Th2 response during the egg stage of the parasite. However, there are several major differences between our experiments and these observations, which may explain this apparent discrepancy.
First, we use fully differentiated memory cells and we do not know for sure in how far these can be compared to the memory cells present following the acute phase of the parasite or more importantly the vaccination using an effective adjuvant. Indeed, it is possible that the memory cells likely to be produced during the acute phase might not be as differentiated as these that we produce artificially in vitro with a cocktail of cytokines and antibodies to cytokines. If the cells generated following the acute phase of the parasitemia are not as differentiated, they may be less resilient to manipulation by the parasite, and hence could be manipulated during the egg-production phase. Secondly, unlike in our case, the antigen specificity of the Th2 immune response induced during the egg-phase could potentially be different to the antigen specificity of the immune response against the acute phase. Hence, the cells that are Th2 during the egg-phase of the infection could (at least theoretically) be different from these of the acute phase. In that case, there would be no need for the acute-phase memory cells to change their phenotype, they would just be superseded by the cells of the egg-phase. Of course, in our experiments this is not an option as all cells are specific for OVA and therefore, we are only considering a change in phenotype during the manipulation. This is a much greater ask of the parasite’s ability to manipulate the immune response. However, our experiments are still pertinent for vaccine development as they demonstrate that it might be possible to establish an immune memory response against an antigen that is resilient to manipulation and thus it is worthwhile to pursue this goal identifying both suitable antigens and effective adjuvants that promote full the differentiation of memory Th cells. Such vaccines would be able to withstand manipulation by the S. mansoni and hence protect the vaccinated host. Whether this can also be achieved for other pathogens remains to be explored.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




