
Therapeutic Potential of Human Fetal Mesenchymal Stem Cells in Musculoskeletal Disorders: A Narrative Review

 ,
,  ,
,
Abstract
:1. Introduction
2. Biological Feature of Human Fetal Mesenchymal Stem Cells (hfMSCs)
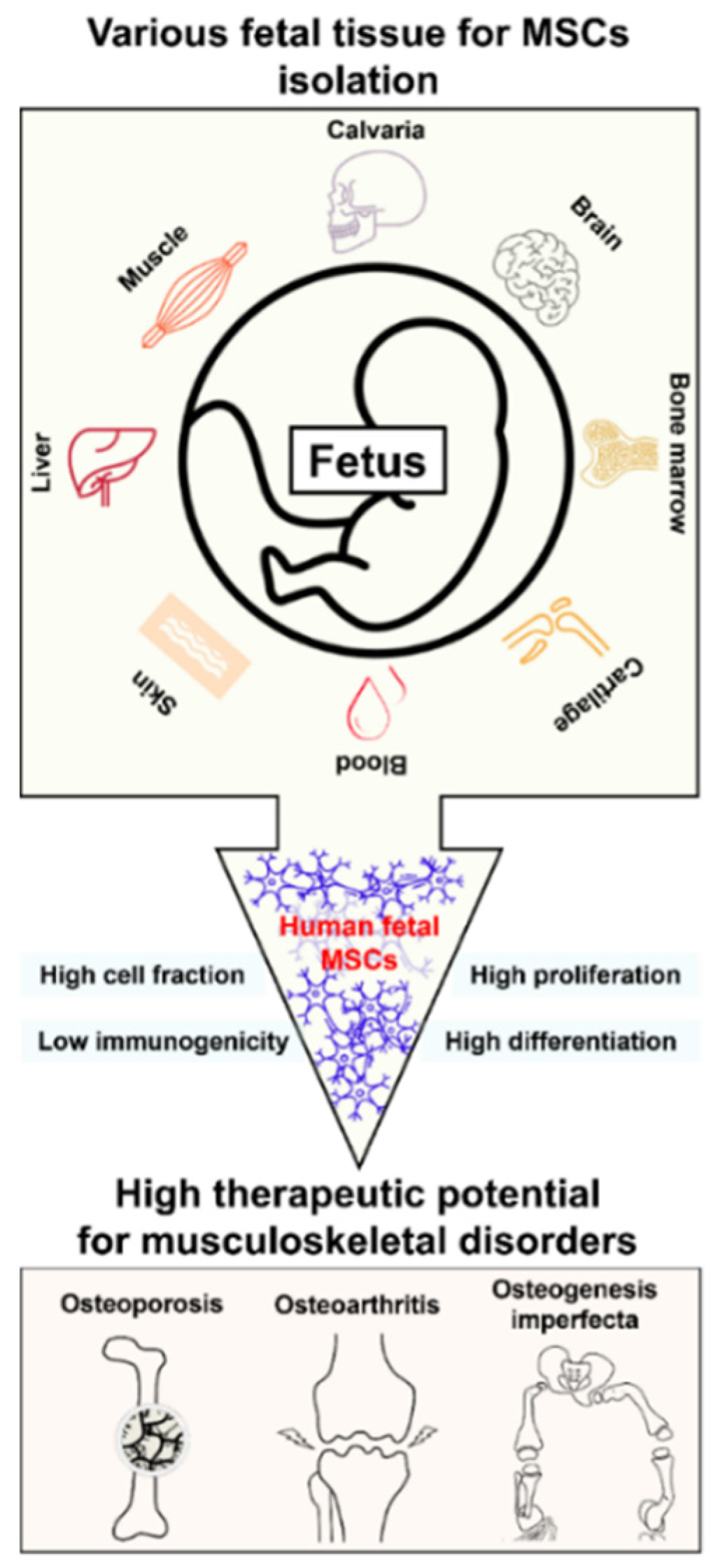
2.1. Sources of hfMSCs
2.2. Biological Components of hfMSCs
2.3. Immunomodulation of hfMSCs
2.4. Summary
3. Preclinical and Clinical Investigation of hfMSCs
3.1. Feasibility of hfMSCs for the Treatment of Musculoskeletal Disorders
3.2. Preclinical Research of hfMSCs
3.3. Clinical Trials Involving hfMSCs
4. Future Prospects and Conclusions
4.1. Future Prospects of hfMSCs
4.2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Miguel, M.P.; Fuentes-Julián, S.; Blázquez-Martínez, A.; Pascual, C.Y.; Aller, M.A.; Arias, J.; Arnalich-Montiel, F. Immunosuppressive properties of mesenchymal stem cells: Advances and applications. Curr. Mol. Med. 2012, 12, 574–591. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar] [PubMed]
- Kadiyala, S.; Young, R.G.; Thiede, M.A.; Bruder, S.P. Culture expanded canine mesenchymal stem cells possess osteochondrogenic potential in vivo and in vitro. Cell Transpl. 1997, 6, 125–134. [Google Scholar] [CrossRef]
- Erices, A.; Conget, P.; Minguell, J.J. Mesenchymal progenitor cells in human umbilical cord blood. Br. J. Haematol. 2000, 109, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.T.; Southerland, S.S.; Souza, J.; Calcutt, A.F.; Cartledge, R.G. Cells isolated from adult human skeletal muscle capable of differentiating into multiple mesodermal phenotypes. Am. Surg. 1999, 65, 22–26. [Google Scholar] [PubMed]
- De Bari, C.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef]
- In’t Anker, P.S.; Scherjon, S.A.; van der Keur, C.K.; Noort, W.A.; Claas, F.H.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 2003, 102, 1548–1549. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.J.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- O’Donoghue, K.; Chan, J. Human fetal mesenchymal stem cells. Curr. Stem Cell Res. Ther. 2006, 1, 371–386. [Google Scholar] [CrossRef]
- Götherström, C.; Ringdén, O.; Tammik, C.; Zetterberg, E.; Westgren, M.; Le Blanc, K. Immunologic properties of human fetal mesenchymal stem cells. Am. J. Obstet. Gynecol. 2004, 190, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.M.; Gibran, N.S. Mesenchymal stem cells: Paracrine signaling and differentiation during cutaneous wound repair. Exp. Cell Res. 2010, 316, 2213–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campagnoli, C.; Roberts, I.A.; Kumar, S.; Bennett, P.R.; Bellantuono, I.; Fisk, N.M. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 2001, 98, 2396–2402. [Google Scholar] [CrossRef] [PubMed]
- Götherström, C.; Ringdén, O.; Westgren, M.; Tammik, C.; Le Blanc, K. Immunomodulatory effects of human fetal liver-derived mesenchymal stem cells. Bone Marrow Transpl. 2003, 32, 265–272. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Teoh, S.H.; Chong, M.S.; Schantz, J.T.; Fisk, N.M.; Choolani, M.A.; Chan, J. Superior osteogenic capacity for bone tissue engineering of fetal compared with perinatal and adult mesenchymal stem cells. Stem Cells 2009, 27, 126–137. [Google Scholar] [CrossRef]
- Guillot, P.V.; De Bari, C.; Dell’Accio, F.; Kurata, H.; Polak, J.; Fisk, N.M. Comparative osteogenic transcription profiling of various fetal and adult mesenchymal stem cell sources. Differentiation 2008, 76, 946–957. [Google Scholar] [CrossRef]
- Yudoh, K.; Matsuno, H.; Osada, R.; Nakazawa, F.; Katayama, R.; Kimura, T. Decreased cellular activity and replicative capacity of osteoblastic cells isolated from the periarticular bone of rheumatoid arthritis patients compared with osteoarthritis patients. Arthritis Rheum. 2000, 43, 2178–2188. [Google Scholar] [CrossRef]
- In’t Anker, P.S.; Noort, W.A.; Scherjon, S.A.; van der Keur, C.K.; Kruisselbrink, A.B.; van Bezooijen, R.L.; Beekhuizen, W.; Willemze, R.; Kanhai, H.H.; Fibbe, W.E. Mesenchymal stem cells in human second-trimester bone marrow, liver, lung, and spleen exhibit a similar immunophenotype but a heterogeneous mutilineage differentiation potential. Haematologica 2003, 88, 845–852. [Google Scholar]
- Fan, C.G.; Tang, F.W.; Zhang, Q.J.; Lu, S.H.; Liu, H.Y.; Zhao, Z.M.; Liu, B.; Han, Z.B.; Han, Z.C. Characterization and neural differentiation of fetal lung mesenchymal stem cells. Cell Transpl. 2005, 14, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Liao, L.; Wang, Q.; Ma, L.; Ma, G.; Jiang, X.; Zhao, R.C. Isolation and identification of mesenchymal stem cells from human fetal pancreas. J. Lab. Clin. Med. 2003, 141, 342–349. [Google Scholar] [CrossRef]
- Zhang, L.; Hong, T.P.; Hu, J.; Liu, Y.N.; Wu, Y.H.; Li, L.S. Nestin-positive progenitor cells isolated from human fetal pancreas have phenotypic markers identical to mesenchymal stem cells. World J. Gastroenterol. 2005, 11, 2906–2911. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liao, L.; Cao, Y.; Jiang, X.; Zhao, R.C. Establishment and properties of fetal dermis-derived mesenchymal stem cell lines: Plasticity in vitro and hematopoietic protection in vivo. Bone Marrow Transpl. 2005, 36, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzhaninova, A.A.; Gornostaeva, S.N.; Goldshtein, D.V. Isolation and phenotypical characterization of mesenchymal stem cells from human fetal thymus. Bull. Exp. Biol. Med. 2005, 139, 134–140. [Google Scholar] [CrossRef]
- Kaviani, A.; Perry, T.E.; Dzakovic, A.; Jennings, R.W.; Ziegler, M.M.; Fauza, D.O. The amniotic fluid as a source of cells for fetal tissue engineering. J. Pediatr. Surg. 2001, 36, 1662–1665. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.S.; Lee, J.L.; Chang, Y.J.; Hwang, S.M. Isolation of human multipotent mesenchymal stem cells from second-triester amniotic fluid using a novel two-stage culture protocol. Hum. Reprod. 2004, 19, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Prusa, A.R.; Marton, E.; Rosner, M.; Bettelheim, D.; Lubec, G.; Pollack, A.; Bernaschek, G.; Hengstschläger, M. Neurogenic cells in human amniotic fluid. Am. J. Obstet. Gynecol. 2004, 191, 309–314. [Google Scholar] [CrossRef]
- O’Donoghue, K.; Fisk, N.M. Fetal stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 853–875. [Google Scholar] [CrossRef]
- Zhao, P.; Ise, H.; Hongo, M.; Ota, M.; Konishi, I.; Nikaido, T. Human amniotic mesenchymal cells have some characteristics of cardiomyocytes. Transplantation 2005, 79, 528–535. [Google Scholar] [CrossRef]
- Ryan, J.M.; Barry, F.P.; Murphy, J.M.; Mahon, B.P. Mesenchymal stem cells avoid allogeneic rejection. J. Inflamm. 2005, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Choo, S.Y. The HLA system: Genetics, immunology, clinical testing, and clinical implications. Yonsei Med. J. 2007, 48, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, W.M. The mother-child union: The case of missing self and protection of the fetus. Proc. Natl Acad. Sci. USA 1997, 94, 5998–6000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagar, R.; Walther-Jallow, L.; David, A.L.; Götherström, C.; Westgren, M. Fetal mesenchymal stromal cells: An opportunity for prenatal cellular therapy. Curr. Stem Cell Rep. 2018, 4, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Shao, J.Z.; Xiang, L.X.; Dong, X.J.; Zhang, G.R. Mesenchymal stem cells: A promising candidate in regenerative medicine. Int. J. Biochem. Cell Biol. 2008, 40, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Da Meirelles, L.S.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef]
- Le Blanc, K.; Tammik, L.; Sundberg, B.; Haynesworth, S.E.; Ringdén, O. Mesenchymal stem cells inhibit and stimulate mixed lymphocyte cultures and mitogenic responses independently of the major histocompatibility complex. Scand. J. Immunol. 2003, 57, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.D.; Whartenby, K.A. Mesenchymal stem cells: Emerging mechanisms of immunomodulation and therapy. World J. Stem Cells 2014, 6, 526–539. [Google Scholar] [CrossRef]
- Fibbe, W.E. Mesenchymal stem cells. A potential source for skeletal repair. Ann. Rheum. Dis. 2002, 61 (Suppl. 2), ii29–ii31. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.; O’Donoghue, K.; Kennea, N.L.; de la Fuente, J.; Kumar, S.; Morgan, J.; Fisk, N. Myogenic potential of fetal mesenchymal stem cells. Ann. Acad. Med. Singap. 2003, 32 (Suppl. 5), S11–S13. [Google Scholar]
- Chan, J.; O’Donoghue, K.; Gavina, M.; Torrente, Y.; Kennea, N.; Mehmet, H.; Stewart, H.; Watt, D.J.; Morgan, J.E.; Fisk, N.M. Galectin-1 induces skeletal muscle differentiation in human fetal mesenchymal stem cells and increases muscle regeneration. Stem Cells 2006, 24, 1879–1891. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Teoh, S.H.; Chong, M.S.; Lee, E.S.; Tan, L.G.; Mattar, C.N.; Fisk, N.M.; Choolani, M.; Chan, J. Neo-vascularization and bone formation mediated by fetal mesenchymal stem cell tissue-engineered bone grafts in critical-size femoral defects. Biomaterials 2010, 31, 608–620. [Google Scholar] [CrossRef]
- Morganstein, D.L.; Wu, P.; Mane, M.R.; Fisk, N.M.; White, R.; Parker, M.G. Human fetal mesenchymal stem cells differentiation into brown and white adipocytes: A role for ERRα in human UCP1 expression. Cell Res. 2010, 20, 434–444. [Google Scholar] [CrossRef]
- Choi, W.H.; Kim, H.R.; Lee, S.J.; Jeong, N.; Park, S.R.; Choi, B.H.; Min, B.H. Fetal cartilage-derived cells have stem cell properties and are a highly potent cell source for cartilage regeneration. Cell Transplant. 2016, 25, 449–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, M.A.; Wynn, R.F.; Jowitt, S.N.; Wraith, J.E.; Fairbairn, L.J.; Bellantuono, I. Study of telomere length reveals rapid aging of human marrow stromal cells following in vitro expansion. Stem Cells 2004, 22, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Mezey, E. The fate of neural crest stem cells: Nature vs nurture. Mol. Psychiatry 2003, 8, 128–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, S.A.; Deamond, S.F.; Ts’o, P.O. In vitro senescence of Syrian hamster mesenchymal cells of fetal to aged adult origin. Inverse relationship between in vivo donor age and in vitro proliferative capacity. Mech. Ageing Dev. 1986, 34, 151–173. [Google Scholar] [CrossRef]
- Banfi, A.; Bianchi, G.; Notaro, R.; Luzzatto, L.; Cancedda, R.; Quarto, R. Replicative aging and gene expression in long-term cultures of human bone marrow stromal cells. Tissue Eng. 2002, 8, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells-current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Prusa, A.R.; Hengstschläger, M. Amniotic fluid cells and human stem cell research: A new connection. Signature 2002, 8, 253–257. [Google Scholar]
- Sellam, J.; Berenbaum, F. The role of synovitis in pathophysiology and clinical symptoms of osteoarthritis. Nat. Rev. Rheumatol. 2010, 6, 625–635. [Google Scholar] [CrossRef]
- Le Blanc, K.; Götherström, C.; Ringdén, O.; Hassan, M.; McMahon, R.; Horwitz, E.; Anneren, G.; Axelsson, O.; Nunn, J.; Ewald, U.; et al. Fetal mesenchymal stem-cell engraftment in bone after in utero transplantation in a patient with severe osteogenesis imperfecta. Transplantation 2005, 79, 1607–1614. [Google Scholar] [CrossRef]
- Götherström, C.; Westgren, M.; Shaw, S.W.S.; Åström, E.; Biswas, A.; Byers, P.H.; Mattar, C.N.; Graham, G.E.; Taslimi, J.; Ewald, U.; et al. Pre- and postnatal transplantation of fetal mesenchymal stem cells in osteogenesis imperfecta: Two-center experience. Stem Cells Transl. Med. 2014, 3, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lee, W.Y.W.; Huang, B.; Zhang, J.F.; Wu, T.Y.; Jiang, X.; Wang, C.C.; Li, G. Secretome of human fetal mesenchymal stem cell ameliorates replicative senescence. Stem Cells Dev. 2016, 25, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, B.; Sun, Y.; Wu, T.; Liu, Y.; Zhang, J.; Lee, W.Y.; Pan, X.; Chai, Y.; Li, G. Human fetal mesenchymal stem cell secretome enhances bone consolidation in distraction osteogenesis. Stem Cell Res. Ther. 2016, 7, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arjmand, B.; Goodarzi, P.; Aghayan, H.R.; Payab, M.; Rahim, F.; Alavi-Moghadam, S.; Mohamadi-Jahani, F.; Larijani, B. Co-transplantation of human fetal mesenchymal and hematopoietic stem cells in type 1 diabetic mice model. Front. Endocrinol. 2019, 10, 761. [Google Scholar] [CrossRef] [Green Version]
- Ramkisoensing, A.A.; Pijnappels, D.A.; Askar, S.F.A.; Passier, R.; Swildens, J.; Goumans, M.J.; Schutte, C.I.; de Vries, A.A.F.; Scherjon, S.; Mummery, C.L.; et al. Human embryonic and fetal mesenchymal stem cells differentiate toward three different cardiac lineages in contrast to their adult counterparts. PLoS ONE 2011, 6, e24164. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Ao, Q.; Wang, X.; Cao, Y.; Liu, Y.; Zheng, S.G.; Tian, X. Mesenchymal stem cell-derived exosome: A promising biological tool in nanomedicine. Front. Pharmacol. 2020, 11, 590470. [Google Scholar] [CrossRef]
- Anderson, J.D.; Johansson, H.J.; Graham, C.S.; Vesterlund, M.; Pham, M.T.; Bramlett, C.S.; Montgomery, E.N.; Mellema, M.S.; Bardini, R.L.; Contreras, Z.; et al. Comprehensive proteomic analysis of mesenchymal stem cell exosomes reveals modulation of angiogenesis via nuclear factor-kappa B signaling. Stem Cells 2016, 34, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Yuan, O.D.; Lin, C.; Wagner, J.; Archard, J.A.; Deng, P.; Halmai, J.; Bauer, G.; Fink, K.D.; Fury, B.; Perotti, N.H.; et al. Exosome derived from human primed mesenchymal stem cells incduce mitosis and potentiate growth factor secretion. Stem Cells Dev. 2019, 28, 398–409. [Google Scholar] [CrossRef] [Green Version]
- Moisseiev, E.; Anderson, J.D.; Oltjen, S.; Goswami, M.; Zawadzki, R.J.; Nolta, J.A.; Park, S.S. Protective effect of intravitreal administration of exosomes derived from mesenchymal stem cells on retinal ischemia. Curr. Eye Res. 2017, 42, 1358–1367. [Google Scholar] [CrossRef]
- Deng, P.; Anderson, J.D.; Yu, A.S.; Annett, G.; Fink, K.D.; Nolta, J.A. Engineered BDNF producing cells as a potential treatment for neurologic disease. Expert Opin. Biol. Ther. 2016, 16, 1025–1033. [Google Scholar] [CrossRef] [Green Version]
- Showalter, M.R.; Wancewicz, B.; Fiehn, O.; Archard, J.A.; Clayton, S.; Wagner, J.; Deng, P.; Halmai, J.; Fink, K.D.; Bauer, G.; et al. Primed mesenchymal stem cells package exosomes with metabolites associated with immunomodulation. Biochem. Biophys. Res. Commun. 2019, 512, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Komaki, M.; Numata, Y.; Morioka, C.; Honda, I.; Tooi, M.; Yokoyama, N.; Ayame, H.; Iwasaki, K.; Taki, A.; Oshima, N.; et al. Exosomes of human placenta-derived mesenchymal stem cells stimulate angiogenesis. Stem Cell Res. Ther. 2017, 8, 219. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiao, Y.; Pan, Y.; Zhang, L.; Gong, H.; Qi, Y.; Wang, M.; Gong, H.; Shao, M.; Wang, X.; et al. Fetal dermal mesenchymal stem cell-derived exosomes accelerate cutaneous wound healing by activating notch signaling. Stem Cells Int. 2019, 2019, 2402916. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Herr, F.; Vernochet, A.; Mennesson, B.; Oberlin, E.; Durrbach, A. Human fetal liver mesenchymal stem cell-derived exosomes impair natural killer cell function. Stem Cells Dev. 2019, 28, 44–55. [Google Scholar] [CrossRef]
- Applegate, L.A.; Scaletta, C.; Hirt-Burri, N.; Raffoul, W.; Pioletti, D. Whole-cell bioprocessing of human fetal cells for tissue engineering of skin. Skin Pharmacol. Physiol. 2009, 22, 63–73. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Tissue Origin (Age of Donor) | Biological Properties | Phenotype (Positive) | Phenotype (Negative) | Proliferation | Ref. |
|---|---|---|---|---|---|
| Blood (Fetus in 16–26 weeks) | CD29, CD44, CD106, CD105, CD73, CD49b, vimentin, laminin, fibronectin | CD45, CD14, CD68, CD34, CD31, HLA-DR, type I collagen | Faster doubling time as every 24–30 h compared to adult No change until 20 passages | [13] | |
| Bone marrow | Adipogenic, osteogenic (Bone marrow derived MSCs > Liver derived MSCs) | CD29, CD44, CD106, CD105, CD73, CD49b, vimentin, laminin, fibronectin | CD45, CD14, CD68, CD34, CD31, HLA-DR, type I collagen | [13,18] | |
| Liver | CD29, CD44, CD54, CD106, CD105, CD73, CD49b, vimentin, laminin, fibronectin | CD45, CD14, CD68, CD34, CD31, HLA-DR, type I collagen | [18] | ||
| Lung | CD58, CD71, CD29, CD44, CD54, CD13, CD90, CD105, CD73, CD49e | CD45, CD14, CD31, CD50, CD106, CD11a, HLA-DR, | Stable until 40 passages | [18,19] | |
| Pancreas (Pregnancy in second trimester) | Osteogenic, adipogenic | CD29, CD44, CD13, CD90, CD147, vimentin, type I collagen | CD45, CD34, HLA-DR | Stable until 30 passages | [20,21] |
| Dermis | Bone, fat, nerve | CD90 | CD45, CD34, CD38, CD117, HLA-DR | [22] | |
| Thymus (Pregnancy in second trimester) | Myoblast, chondrogenic osteogenic, adipogenic | CD71, CD44, CD54, CD105, CD90, CD49b, vimentin | CD45, CD34, CD38, HLA Class I, HLA-DR | [23] | |
| Amniotic fluid (Pregnancy in second trimester) | Neural pathway, cardiomyocytes, osteogenic, adipogenic | CD29, CD44, CD105, CD73, CD90, OCT-4, vimentin, type I collagen | CD45, CD14, CD34, CD31, CD106, CD11a, CD13, CD117, HLA-DR | [8,24,25,26,28] | |
| Calvaria | Osteogenic, chondrogenic, adipogenic | CD105, CD90, CD44, CD29 | SSEA-3, TRA-1-81, CD34, CD45 | Author’s unpublished data |
| Biological Property | hfMSCs | Adult MSCs | Ref. |
|---|---|---|---|
| Immune response | HLA-DR * expression after 7 d exposure to IFN-γ | HLA-DR expression after 1 d exposure to IFN-γ | [11,12] |
| Immunogenic | Less | more | [32] |
| Proliferation (Population doubling time) | 24–30 h | 48–72 h | [13,14] |
| Telomere length | Longer | Shorter | [10] |
| Osteogenic Differentiation | Higher | Lower | [15,16] |
| Stemness (Colony-forming) | hfMSCs two times higher than adult MSCs | [15] | |
| Year | Patient | Mutation | Source | OI Phenotype | Cell Number (×106) | Outcome | Ref. | |
|---|---|---|---|---|---|---|---|---|
| Prenatal | Postnatal | |||||||
| 2005 | A | COL1A2, Gly33743Asp | Fetal liver (10 weeks) | III/IV | 6.5 hfMSCs at 32 weeks | [50] | ||
| 2014 | B | COL1A2, Gly33743Asp; Gly913Asp | Fetal liver (7 weeks 3 days and 10 weeks) | III | 6.5 hfMSCs at 31 weeks (5/kg) | 42 at 8 years and 2 months (2.8/kg) | No new fractures, improved growth velocity | [51] |
| C | COL1A2, Gly33743Asp; Gly130Asp | IV | 4 hfMSCs at 31 weeks (30/kg) | 88 at 19 months and 11 days (10/kg) | No new fractures, improved growth velocity | |||
| D | COL1A2, Gly33743Asp; Gly915Asp | II/III | None | None | Deceased at 5 months of age | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, I.; Rim, J.; Lee, J.; Jang, I.; Jung, B.; Kim, K.; Lee, S. Therapeutic Potential of Human Fetal Mesenchymal Stem Cells in Musculoskeletal Disorders: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 1439. https://doi.org/10.3390/ijms23031439
Song I, Rim J, Lee J, Jang I, Jung B, Kim K, Lee S. Therapeutic Potential of Human Fetal Mesenchymal Stem Cells in Musculoskeletal Disorders: A Narrative Review. International Journal of Molecular Sciences. 2022; 23(3):1439. https://doi.org/10.3390/ijms23031439
Chicago/Turabian StyleSong, Insun, Jongseop Rim, Jaemin Lee, Inseok Jang, Bosung Jung, Kisoo Kim, and Soonchul Lee. 2022. "Therapeutic Potential of Human Fetal Mesenchymal Stem Cells in Musculoskeletal Disorders: A Narrative Review" International Journal of Molecular Sciences 23, no. 3: 1439. https://doi.org/10.3390/ijms23031439