
The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases

 , , , ,
, , , ,
Abstract
:
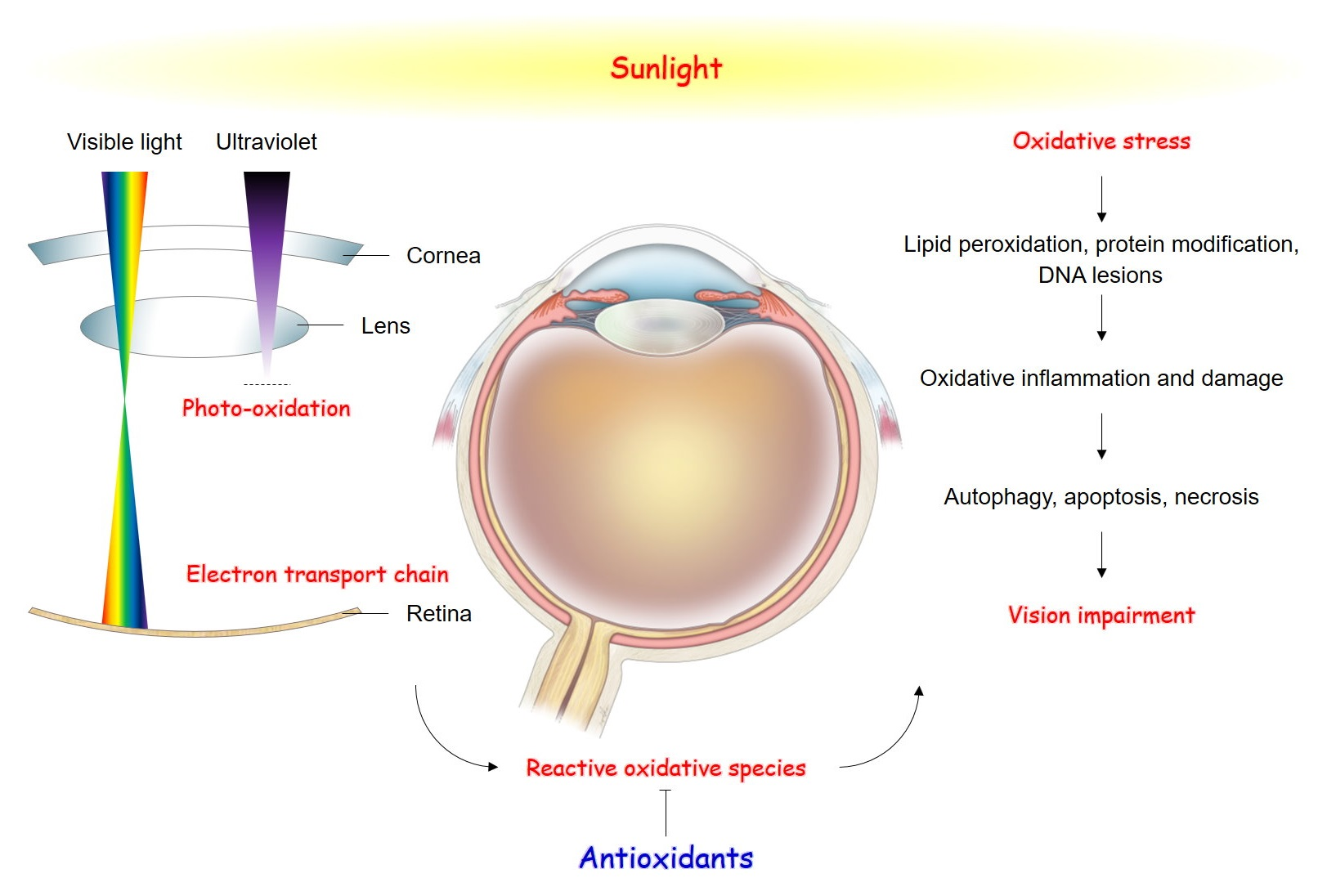
1. Introduction
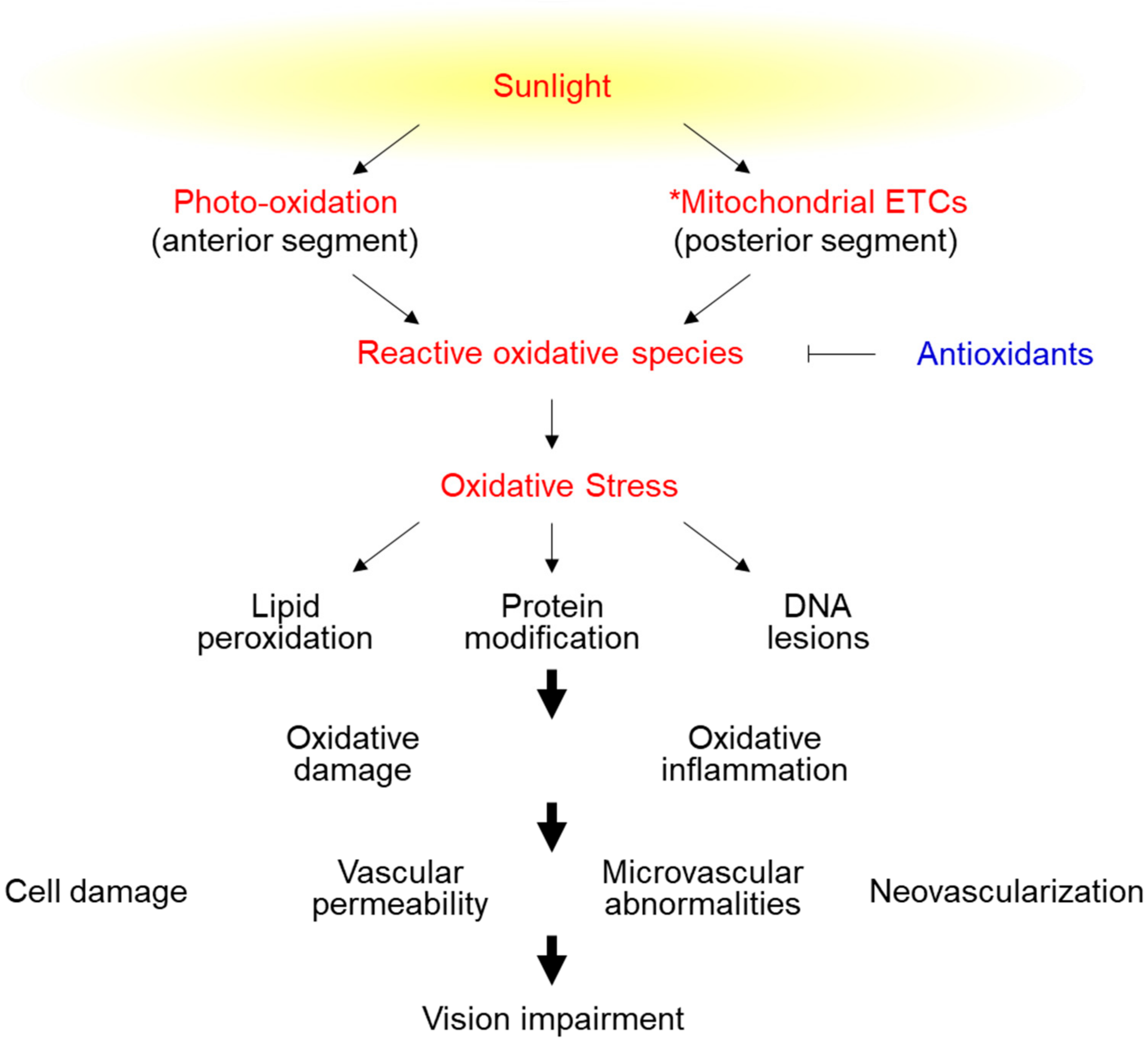
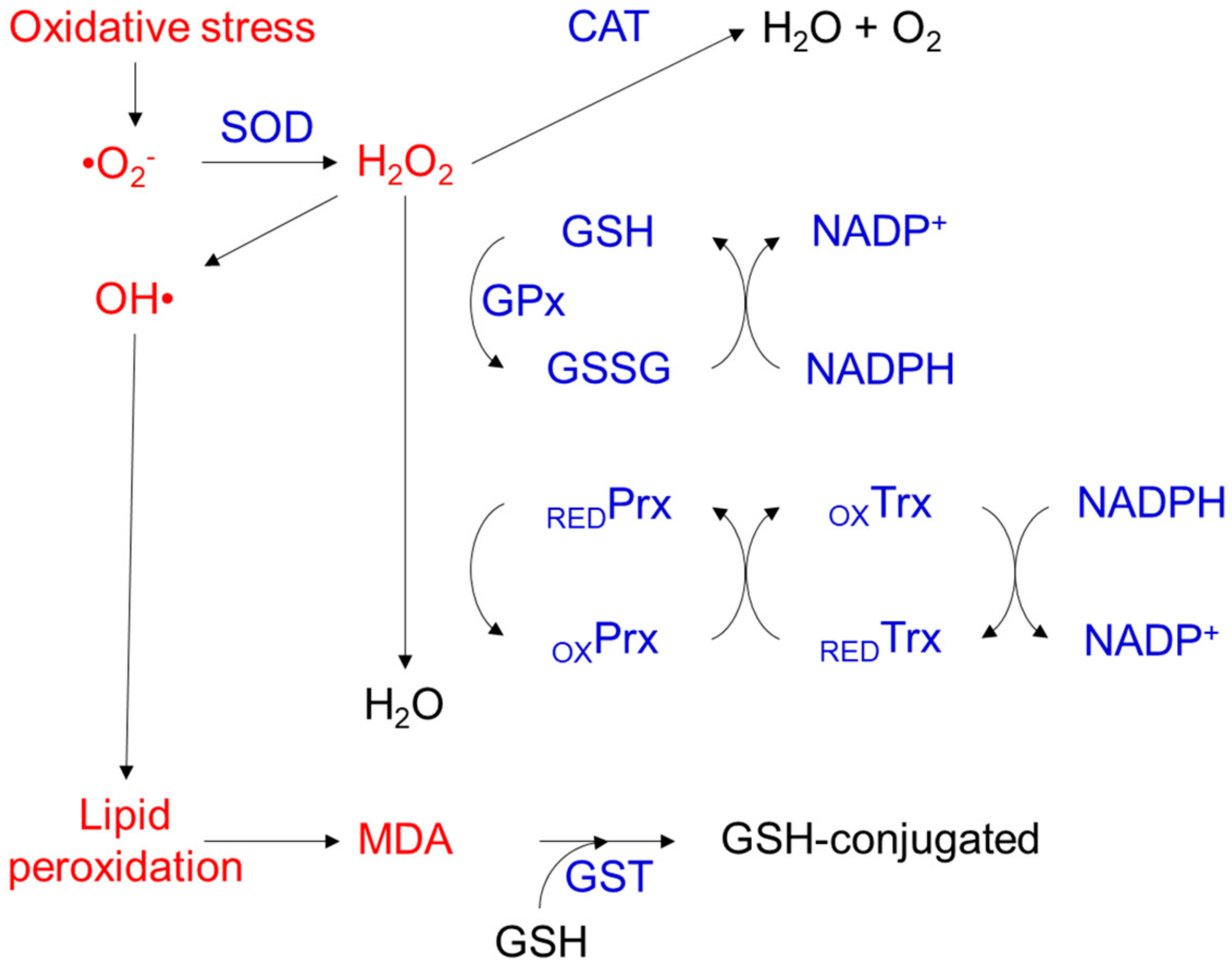
2. Molecular Aspects of Oxidative Stress-Induced Cell Damage
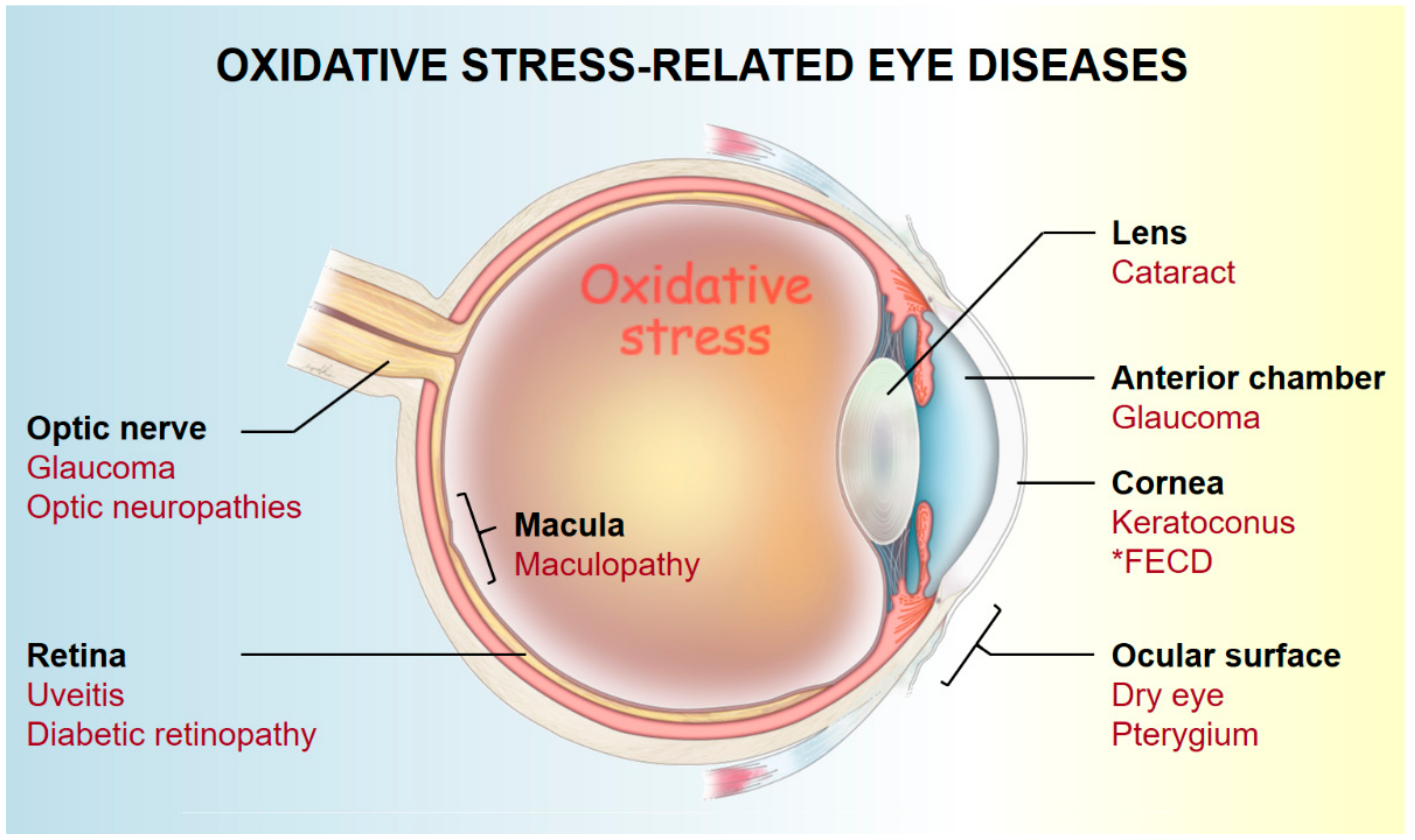
3. Pathogenesis of Oxidative Stress-Related Eye Diseases
3.1. Ocular Surface (Cornea and Conjunctiva)
3.2. Anterior Chamber
3.3. Lens
3.4. Retina and Optic Nerve
3.5. Others (Oxidative Stress-Related Surgical Complications)
4. Antioxidant Imbalance, Biomarkers, and the Potential of Predictive Diagnostics in Oxidative Stress-Related Ocular Diseases
4.1. Keratoconus (KC)
4.1.1. Cornea
4.1.2. Tear Film
4.1.3. Serum
4.2. Senile Cataract
4.2.1. Lens
4.2.2. Aqueous Humor
4.2.3. Serum
4.3. Age-Related Macular Degeneration (AMD)
4.3.1. Retina
4.3.2. Serum
4.4. Glaucoma
4.4.1. Aqueous Humor
4.4.2. Serum
4.5. The Potential of Predictive Diagnostics for Oxidative Stress-Related Eye Diseases
5. Natural Antioxidant Therapy for Oxidative Stress-Related Eye Diseases
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harman, D. Aging: A Theory Based on Free Radical and Radiation Chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aruoma, O.I. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.L.; Lim, H.W.; Mohammad, T.F. Sunscreens and Photoaging: A Review of Current Literature. Am. J. Clin. Dermatol. 2021, 22, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Zinflou, C.; Rochette, P.J. Ultraviolet A-induced oxidation in cornea: Characterization of the early oxidation-related events. Free Radic. Biol. Med. 2017, 108, 118–128. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxidative Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.Y.; Liu, P.K.; Wen, Y.T.; Quinn, P.M.J.; Levi, S.R.; Wang, N.K.; Tsai, R.K. Role of Oxidative Stress in Ocular Diseases Associated with Retinal Ganglion Cells Degeneration. Antioxidants 2021, 10, 1948. [Google Scholar] [CrossRef]
- Sunkireddy, P.; Jha, S.N.; Kanwar, J.R.; Yadav, S.C. Natural antioxidant biomolecules promises future nanomedicine based therapy for cataract. Colloids Surf. B Biointerfaces 2013, 112, 554–562. [Google Scholar] [CrossRef]
- Fuentes-León, F.; Peres de Oliveira, A.; Quintero-Ruiz, N.; Munford, V.; Satoru Kajitani, G.; Coimbra Brum, A.; Schuch, A.P.; Colepicolo, P.; Sánchez-Lamar, A.; Menck, C.F.M. DNA Damage Induced by Late Spring Sunlight in Antarctica. Photochem. Photobiol. 2020, 96, 1215–1220. [Google Scholar] [CrossRef]
- Natoli, R.; Jiao, H.; Barnett, N.L.; Fernando, N.; Valter, K.; Provis, J.M.; Rutar, M. A model of progressive photo-oxidative degeneration and inflammation in the pigmented C57BL/6J mouse retina. Exp. Eye Res. 2016, 147, 114–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.I.; Jang, Y.P. The Protective Effect of Brown-, Gray-, and Blue-Tinted Lenses against Blue LED Light-Induced Cell Death in A2E-Laden Human Retinal Pigment Epithelial Cells. Ophthalmic Res. 2017, 57, 118–124. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Saccà, S.C.; Cutolo, C.A.; Ferrari, D.; Corazza, P.; Traverso, C.E. The Eye, Oxidative Damage and Polyunsaturated Fatty Acids. Nutrients 2018, 10, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxidative Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochkov, V.N.; Oskolkova, O.V.; Birukov, K.G.; Levonen, A.L.; Binder, C.J.; Stöckl, J. Generation and biological activities of oxidized phospholipids. Antioxid. Redox Signal. 2010, 12, 1009–1059. [Google Scholar] [CrossRef] [Green Version]
- Di Gioia, M.; Zanoni, I. Dooming Phagocyte Responses: Inflammatory Effects of Endogenous Oxidized Phospholipids. Front. Endocrinol. 2021, 12, 626842. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G.; Grune, T.; Sereikaite, J. Role of Oxidative, Nitrative, and Chlorinative Protein Modifications in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2018, 2018, 3267898. [Google Scholar] [CrossRef]
- Shokolenko, I.; Venediktova, N.; Bochkareva, A.; Wilson, G.L.; Alexeyev, M.F. Oxidative stress induces degradation of mitochondrial DNA. Nucleic Acids Res. 2009, 37, 2539–2548. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.D.; Dizdaroglu, M.; Cooke, M.S. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res. 2004, 567, 1–61. [Google Scholar] [CrossRef]
- Roos, W.P.; Kaina, B. DNA damage-induced cell death by apoptosis. Trends Mol. Med. 2006, 12, 440–450. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.R.; Crawford, M.J.; Chaturvedi, M.M.; Jain, S.K.; Aggarwal, B.B.; Al-Ubaidi, M.R.; Agarwal, N. Photo-oxidative stress down-modulates the activity of nuclear factor-kappaB via involvement of caspase-1, leading to apoptosis of photoreceptor cells. J. Biol. Chem. 1999, 274, 3734–3743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitbaev, K.A.; Murkamilov, I.T.; Fomin, V.V. Molecular mechanisms of aging: The role of oxidative stress and epigenetic modifications. Adv. Gerontol. 2019, 32, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Nakka, V.P.; Prakash-Babu, P.; Vemuganti, R. Crosstalk between Endoplasmic Reticulum Stress, Oxidative Stress, and Autophagy: Potential Therapeutic Targets for Acute CNS Injuries. Mol. Neurobiol. 2016, 53, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klionsky, D.J.; Abeliovich, H.; Agostinis, P.; Agrawal, D.K.; Aliev, G.; Askew, D.; Baba, M.; Baehrecke, E.H.; Bahr, B.A.; Ballabio, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes. Autophagy 2008, 4, 151–175. [Google Scholar] [CrossRef]
- Hsueh, Y.J.; Meir, Y.J.; Yeh, L.K.; Wang, T.K.; Huang, C.C.; Lu, T.T.; Cheng, C.M.; Wu, W.C.; Chen, H.C. Topical Ascorbic Acid Ameliorates Oxidative Stress-Induced Corneal Endothelial Damage via Suppression of Apoptosis and Autophagic Flux Blockage. Cells 2020, 9, 943. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.Y.; Zhong, X.R.; Yu, S.H.; Zhang, L.C.; Liu, Y.N.; Zhang, Y.; Sun, L.K.; Su, J. p62 aggregates mediated Caspase 8 activation is responsible for progression of ovarian cancer. J. Cell. Mol. Med. 2019, 23, 4030–4042. [Google Scholar] [CrossRef]
- Laporte, C.; Kosta, A.; Klein, G.; Aubry, L.; Lam, D.; Tresse, E.; Luciani, M.F.; Golstein, P. A necrotic cell death model in a protist. Cell Death Differ. 2007, 14, 266–274. [Google Scholar] [CrossRef]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding oxidants and antioxidants: Classical team with new players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.S.; Singh, S.P.; Singhal, P.; Horne, D.; Singhal, J.; Awasthi, S. Antioxidant role of glutathione S-transferases: 4-Hydroxynonenal, a key molecule in stress-mediated signaling. Toxicol. Appl. Pharmacol. 2015, 289, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. The antioxidants of human extracellular fluids. Arch. Biochem. Biophys. 1990, 280, 1–8. [Google Scholar] [CrossRef]
- Delamere, N.A. Ascorbic Acid and the Eye. In Subcellular Biochemistry: Ascorbic Acid: Biochemistry and Biomedical Cell Biology; Harris, J.R., Ed.; Springer: Boston, MA, USA, 1996; pp. 313–329. [Google Scholar]
- Bragt, P.C.; Bonta, I.L. Oxidant stress during inflammation: Anti-inflammatory effects of antioxidants. Agents Actions 1980, 10, 536–539. [Google Scholar] [CrossRef]
- Augustin, A.J.; Dick, H.B. Oxidative tissue damage after phacoemulsification: Influence of ophthalmic viscosurgical devices. J. Cataract Refract. Surg. 2004, 30, 424–427. [Google Scholar] [CrossRef]
- Özer, M.A.; Polat, N.; Özen, S.; Parlakpınar, H.; Ekici, K.; Polat, A.; Vardı, N.; Tanbek, K.; Yildiz, A. Effects of Molsidomine on Retinopathy and Oxidative Stress Induced by Radiotheraphy in Rat Eyes. Curr. Eye Res. 2017, 42, 803–809. [Google Scholar] [CrossRef]
- Bergandi, L.; Skorokhod, O.A.; Franzone, F.; La Grotta, R.; Schwarzer, E.; Nuzzi, R. Induction of oxidative stress in human aqueous and vitreous humors by Nd:YAG laser posterior capsulotomy. Int. J. Ophthalmol. 2018, 11, 1145–1151. [Google Scholar]
- Hull, D.S.; Green, K. Oxygen free radicals and corneal endothelium. Lens Eye Toxic. Res. 1989, 6, 87–91. [Google Scholar]
- Kowluru, R.A.; Chan, P.S. Oxidative stress and diabetic retinopathy. Exp. Diabetes Res. 2007, 2007, 43603. [Google Scholar] [CrossRef] [Green Version]
- Beatty, S.; Koh, H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.L. Oxidative stress and the eye. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Sacca, S.C.; Roszkowska, A.M.; Izzotti, A. Environmental light and endogenous antioxidants as the main determinants of non-cancer ocular diseases. Mutat. Res. 2013, 752, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Kau, H.C.; Tsai, C.C.; Lee, C.F.; Kao, S.C.; Hsu, W.M.; Liu, J.H.; Wei, Y.H. Increased oxidative DNA damage, 8-hydroxydeoxy- guanosine, in human pterygium. Eye 2006, 20, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Cimpean, A.M.; Sava, M.P.; Raica, M. DNA damage in human pterygium: One-shot multiple targets. Mol. Vis. 2013, 19, 348–356. [Google Scholar]
- Ozturk, B.T.; Yıldırım, M.S.; Zamani, A.; Bozkurt, B. K-ras oncogene mutation in pterygium. Eye 2017, 31, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Balci, M.; Sahin, S.; Mutlu, F.M.; Yağci, R.; Karanci, P.; Yildiz, M. Investigation of oxidative stress in pterygium tissue. Mol. Vis. 2011, 17, 443–447. [Google Scholar]
- Sebastiá, R.; Ventura, M.P.; Solari, H.P.; Antecka, E.; Orellana, M.E.; Burnier, M.N., Jr. Immunohistochemical detection of Hsp90 and Ki-67 in pterygium. Diagn. Pathol. 2013, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.C.; Kao, Y.H.; Hu, P.S.; Chen, J.H. Geldanamycin, a HSP90 inhibitor, attenuates the hypoxia-induced vascular endothelial growth factor expression in retinal pigment epithelium cells in vitro. Exp. Eye Res. 2007, 85, 721–731. [Google Scholar] [CrossRef]
- Zidi, S.; Bediar-Boulaneb, F.; Belguendouz, H.; Belkhelfa, M.; Medjeber, O.; Laouar, O.; Henchiri, C.; Touil-Boukoffa, C. Local pro-inflammatory cytokine and nitric oxide responses are elevated in patients with pterygium. Int. J. Immunopathol. Pharmacol. 2017, 30, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Cantú, E.; Zavala, J.; Valenzuela, J.; Valdez-García, J.E. Molecular Basis of Pterygium Development. Semin. Ophthalmol. 2016, 31, 567–583. [Google Scholar] [CrossRef] [PubMed]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, T.M.; Tomiyoshi, L.M.; Dias, A.C.; Roma, L.P.; Módulo, C.M.; Malki, L.T.; Filho, E.B.; Deminice, R.; Jordão, A.A., Jr.; Cunha, D.A.; et al. Age-dependent changes in rat lacrimal gland anti-oxidant and vesicular related protein expression profiles. Mol. Vis. 2012, 18, 194–202. [Google Scholar] [PubMed]
- Yoon, C.H.; Ryu, J.S.; Hwang, H.S.; Kim, M.K. Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model. Int. J. Mol. Sci. 2020, 21, 4169. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Ren, Y.; Reinach, P.S.; She, Y.; Xiao, B.; Hua, S.; Qu, J.; Chen, W. Reactive oxygen species activated NLRP3 inflammasomes prime environment-induced murine dry eye. Exp. Eye Res. 2014, 125, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lackner, E.M.; Matthaei, M.; Meng, H.; Ardjomand, N.; Eberhart, C.G.; Jun, A.S. Design and analysis of keratoconus tissue microarrays. Cornea 2014, 33, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, K.A.; Blasiak, J.; Szaflik, J.; Szaflik, J.P. Role of biochemical factors in the pathogenesis of keratoconus. Acta Biochim. Pol. 2014, 61, 55–62. [Google Scholar] [CrossRef] [Green Version]
- McKay, T.B.; Hjortdal, J.; Priyadarsini, S.; Karamichos, D. Acute hypoxia influences collagen and matrix metalloproteinase expression by human keratoconus cells in vitro. PLoS ONE 2017, 12, e0176017. [Google Scholar] [CrossRef] [Green Version]
- Vallabh, N.A.; Romano, V.; Willoughby, C.E. Mitochondrial dysfunction and oxidative stress in corneal disease. Mitochondrion 2017, 36, 103–113. [Google Scholar] [CrossRef]
- Chwa, M.; Atilano, S.R.; Hertzog, D.; Zheng, H.; Langberg, J.; Kim, D.W.; Kenney, M.C. Hypersensitive response to oxidative stress in keratoconus corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4361–4369. [Google Scholar] [CrossRef] [Green Version]
- Soiberman, U.; Foster, J.W.; Jun, A.S.; Chakravarti, S. Pathophysiology of Keratoconus: What Do We Know Today. Open Ophthalmol. J. 2017, 11, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chwa, M.; Atilano, S.R.; Reddy, V.; Jordan, N.; Kim, D.W.; Kenney, M.C. Increased stress-induced generation of reactive oxygen species and apoptosis in human keratoconus fibroblasts. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.M.; Jeyabalan, N.; Tripathi, R.; Panigrahi, T.; Johnson, P.J.; Ghosh, A.; Mohan, R.R. Autophagy in corneal health and disease: A concise review. Ocul. Surf. 2019, 17, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Shetty, R.; Sharma, A.; Pahuja, N.; Chevour, P.; Padmajan, N.; Dhamodaran, K.; Jayadev, C.; Nuijts, R.M.M.A.; Ghosh, A.; Nallathambi, J. Oxidative stress induces dysregulated autophagy in corneal epithelium of keratoconus patients. PLoS ONE 2017, 12, e0184628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serbecic, N.; Beutelspacher, S.C. Anti-oxidative vitamins prevent lipid-peroxidation and apoptosis in corneal endothelial cells. Cell Tissue Res. 2005, 320, 465–475. [Google Scholar] [CrossRef]
- Schmedt, T.; Silva, M.M.; Ziaei, A.; Jurkunas, U. Molecular bases of corneal endothelial dystrophies. Exp. Eye Res. 2012, 95, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Jurkunas, U.V.; Bitar, M.S.; Funaki, T.; Azizi, B. Evidence of oxidative stress in the pathogenesis of fuchs endothelial corneal dystrophy. Am. J. Pathol. 2010, 177, 2278–2289. [Google Scholar] [CrossRef]
- Cho, K.-S.; Lee, E.H.; Choi, J.-S.; Joo, C.-K. Reactive oxygen species-induced apoptosis and necrosis in bovine corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 911–919. [Google Scholar]
- Nanda, G.G.; Alone, D.P. REVIEW: Current understanding of the pathogenesis of Fuchs’ endothelial corneal dystrophy. Mol. Vis. 2019, 25, 295–310. [Google Scholar]
- Halilovic, A.; Schmedt, T.; Benischke, A.S.; Hamill, C.; Chen, Y.; Santos, J.H.; Jurkunas, U.V. Menadione-Induced DNA Damage Leads to Mitochondrial Dysfunction and Fragmentation During Rosette Formation in Fuchs Endothelial Corneal Dystrophy. Antioxid. Redox Signal. 2016, 24, 1072–1083. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Vojnovic, D.; Kochevar, I.E.; Jurkunas, U.V. UV-A Irradiation Activates Nrf2-Regulated Antioxidant Defense and Induces p53/Caspase3-Dependent Apoptosis in Corneal Endothelial Cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Lovatt, M.; Adnan, K.; Kocaba, V.; Dirisamer, M.; Peh, G.S.L.; Mehta, J.S. Peroxiredoxin-1 regulates lipid peroxidation in corneal endothelial cells. Redox Biol. 2020, 30, 101417. [Google Scholar] [CrossRef] [PubMed]
- Ziaei, A.; Schmedt, T.; Chen, Y.; Jurkunas, U.V. Sulforaphane decreases endothelial cell apoptosis in fuchs endothelial corneal dystrophy: A novel treatment. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6724–6734. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.C.; Campanelli, J.; Sande, P.; Sánez, D.A.; Keller Sarmiento, M.I.; Rosenstein, R.E. Retinal oxidative stress induced by high intraocular pressure. Free Radic. Biol. Med. 2004, 37, 803–812. [Google Scholar] [CrossRef]
- Izzotti, A.; Saccà, S.C.; Longobardi, M.; Cartiglia, C. Sensitivity of ocular anterior chamber tissues to oxidative damage and its relevance to the pathogenesis of glaucoma. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5251–5258. [Google Scholar] [CrossRef] [Green Version]
- Saccà, S.C.; Izzotti, A. Focus on molecular events in the anterior chamber leading to glaucoma. Cell Mol. Life Sci. 2014, 71, 2197–2218. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, H.; Liu, X.; Han, Y.; Pan, S.; Wang, Y. MiR-181a inhibits human trabecular meshwork cell apoptosis induced by H₂O₂ through the suppression of NF-κB and JNK pathways. Adv. Clin. Exp. Med. 2018, 27, 577–582. [Google Scholar] [CrossRef]
- Saccà, S.C.; Pulliero, A.; Izzotti, A. The dysfunction of the trabecular meshwork during glaucoma course. J. Cell. Physiol. 2015, 230, 510–525. [Google Scholar] [CrossRef]
- Pascolini, D.; Mariotti, S.P. Global estimates of visual impairment: 2010. Br. J. Ophthalmol. 2012, 96, 614. [Google Scholar] [CrossRef] [Green Version]
- Moreau, K.L.; King, J.A. Protein misfolding and aggregation in cataract disease and prospects for prevention. Trends Mol. Med. 2012, 18, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Kisic, B.; Miric, D.; Zoric, L.; Ilic, A.; Dragojevic, I. Antioxidant capacity of lenses with age-related cataract. Oxidative Med. Cell. Longev. 2012, 2012, 467130. [Google Scholar] [CrossRef] [PubMed]
- Spector, A. Review: Oxidative stress and disease. J. Ocul. Pharmacol. Ther. 2000, 16, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Kisic, B.; Miric, D.; Zoric, L.; Ilic, A. Role of lipid peroxidation in the pathogenesis of age-related cataract. In Lipid Peroxidation; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar]
- Miric, D.; Kisic, B.; Zoric, L.; Miric, B.; Mirkovic, M.; Mitic, R. Influence of cataract maturity on aqueous humor lipid peroxidation markers and antioxidant enzymes. Eye 2014, 28, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.F.; Guo, B.; Ye, M.J.; Liao, R.F.; Li, S.L. Protective Effect of Rutin Against H2O2-Induced Oxidative Stress and Apoptosis in Human Lens Epithelial Cells. Curr. Eye Res. 2016, 41, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Delamere, N.A.; Tamiya, S. Expression, regulation and function of Na,K-ATPase in the lens. Prog. Retin. Eye Res. 2004, 23, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, J.H.; Kang, S.S.; Hwang, S.B.; Tchah, H. Topical nerve growth factor attenuates streptozotocin-induced diabetic cataracts via polyol pathway inhibition and Na(+)/K(+)-ATPase upregulation. Exp. Eye Res. 2021, 202, 108319. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Lin, T.; Walker, G.B.; Kurji, K.; Fang, E.; Law, G.; Prasad, S.S.; Kojic, L.; Cao, S.; White, V.; Cui, J.Z.; et al. Parainflammation associated with advanced glycation endproduct stimulation of RPE in vitro: Implications for age-related degenerative diseases of the eye. Cytokine 2013, 62, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Sheu, S.J.; Chen, J.L.; Bee, Y.S.; Lin, S.H.; Shu, C.W. ERBB2-modulated ATG4B and autophagic cell death in human ARPE19 during oxidative stress. PLoS ONE 2019, 14, e0213932. [Google Scholar] [CrossRef]
- Park, C.; Lee, H.; Hong, S.H.; Kim, J.H.; Park, S.K.; Jeong, J.W.; Kim, G.Y.; Hyun, J.W.; Yun, S.J.; Kim, B.W.; et al. Protective effect of diphlorethohydroxycarmalol against oxidative stress-induced DNA damage and apoptosis in retinal pigment epithelial cells. Cutan. Ocul. Toxicol. 2019, 38, 298–308. [Google Scholar] [CrossRef]
- Jarrett, S.G.; Boulton, M.E. Consequences of oxidative stress in age-related macular degeneration. Mol. Asp. Med. 2012, 33, 399–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of Mitochondrial DNA Damage in ROS-Mediated Pathogenesis of Age-Related Macular Degeneration (AMD). Int. J. Mol. Sci. 2019, 20, 2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrysostomou, V.; Rezania, F.; Trounce, I.A.; Crowston, J.G. Oxidative stress and mitochondrial dysfunction in glaucoma. Curr. Opin. Pharmacol. 2013, 13, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G. Multifactorial Pathogenic Processes of Retinal Ganglion Cell Degeneration in Glaucoma towards Multi-Target Strategies for Broader Treatment Effects. Cells 2021, 10, 1372. [Google Scholar] [CrossRef]
- Lin, W.J.; Kuang, H.Y. Oxidative stress induces autophagy in response to multiple noxious stimuli in retinal ganglion cells. Autophagy 2014, 10, 1692–1701. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Ren, Y.L.; Zhai, J.; Zhou, X.Y.; Wu, J. Down-regulated LAMA4 inhibits oxidative stress-induced apoptosis of retinal ganglion cells through the MAPK signaling pathway in rats with glaucoma. Cell Cycle 2019, 18, 932–948. [Google Scholar] [CrossRef]
- Wei, T.; Kang, Q.; Ma, B.; Gao, S.; Li, X.; Liu, Y. Activation of autophagy and paraptosis in retinal ganglion cells after retinal ischemia and reperfusion injury in rats. Exp. Ther. Med. 2015, 9, 476–482. [Google Scholar] [CrossRef]
- Russo, R.; Varano, G.P.; Adornetto, A.; Nazio, F.; Tettamanti, G.; Girardello, R.; Cianfanelli, V.; Cavaliere, F.; Morrone, L.A.; Corasaniti, M.T.; et al. Rapamycin and fasting sustain autophagy response activated by ischemia/reperfusion injury and promote retinal ganglion cell survival. Cell Death Dis. 2018, 9, 981. [Google Scholar] [CrossRef]
- Kitaoka, Y.; Munemasa, Y.; Kojima, K.; Hirano, A.; Ueno, S.; Takagi, H. Axonal protection by Nmnat3 overexpression with involvement of autophagy in optic nerve degeneration. Cell Death Dis. 2013, 4, e860. [Google Scholar] [CrossRef]
- Adornetto, A.; Parisi, V.; Morrone, L.A.; Corasaniti, M.T.; Bagetta, G.; Tonin, P.; Russo, R. The Role of Autophagy in Glaucomatous Optic Neuropathy. Front. Cell Dev. Biol. 2020, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Ventura, A.S.; Wälti, R.; Böhnke, M. Corneal thickness and endothelial density before and after cataract surgery. Br. J. Ophthalmol. 2001, 85, 18–20. [Google Scholar] [CrossRef] [Green Version]
- Suranyi, E.; Berta, A.; Modis, L.; Szalai, E.; Damjanovich, J. Does beta-ray emitting therapy of ciliary body tumors decrease central corneal endothelial cell density? Eur. J. Ophthalmol. 2013, 23, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Slomovic, A.R.; Parrish, R.K., II; Forster, R.K.; Cubillas, A. Neodymium-YAG Laser Posterior Capsulotomy: Central Corneal Endothelial Cell Density. Arch. Ophthalmol. 1986, 104, 536–538. [Google Scholar] [CrossRef] [PubMed]
- Ishii, N.; Yamaguchi, T.; Yazu, H.; Satake, Y.; Yoshida, A.; Shimazaki, J. Factors associated with graft survival and endothelial cell density after Descemet’s stripping automated endothelial keratoplasty. Sci. Rep. 2016, 6, 25276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lass, J.H.; Sugar, A.; Benetz, B.A.; Beck, R.W.; Dontchev, M.; Gal, R.L.; Kollman, C.; Gross, R.; Heck, E.; Holland, E.J. Endothelial cell density to predict endothelial graft failure after penetrating keratoplasty. Arch. Ophthalmol. 2010, 128, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Kettesy, B.; Nemeth, G.; Kemeny-Beke, A.; Berta, A.; Modis, L. Assessment of endothelial cell density and corneal thickness in corneal grafts an average of 5 years after penetrating keratoplasty. Wien. Klin. Wochenschr. 2014, 126, 286–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockmann, T.; Pilger, D.; Brockmann, C.; Maier, A.-K.B.; Bertelmann, E.; Torun, N. Predictive factors for clinical outcomes after primary Descemet’s membrane endothelial keratoplasty for Fuchs’ endothelial dystrophy. Curr. Eye Res. 2019, 44, 147–153. [Google Scholar] [CrossRef]
- Price, M.O.; Lass, J.H.; Price, F.W. Clinical Factors for Early and Late Endothelial Cell Loss After Corneal Transplantation. Curr. Ophthalmol. Rep. 2018, 6, 191–199. [Google Scholar] [CrossRef]
- Yazu, H.; Yamaguchi, T.; Tsubota, K.; Shimazaki, J. Clinical Factors for Rapid Endothelial Cell Loss after Corneal Transplantation: Novel Findings from the Aqueous Humor. Curr. Ophthalmol. Rep. 2019, 7, 89–97. [Google Scholar] [CrossRef]
- Moschos, M.M.; Chatziralli, I.P.; Sergentanis, T.N. Viscoat versus Visthesia during phacoemulsification cataract surgery: Corneal and foveal changes. BMC Ophthalmol. 2011, 11, 9. [Google Scholar] [CrossRef] [Green Version]
- Geffen, N.; Topaz, M.; Kredy-Farhan, L.; Barequet, I.S.; Farzam, N.; Assia, E.I.; Savion, N. Phacoemulsification-induced injury in corneal endothelial cells mediated by apoptosis: In vitro model. J. Cataract Refract. Surg. 2008, 34, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Rosado-Adames, N.; Afshari, N.A. The changing fate of the corneal endothelium in cataract surgery. Curr. Opin. Ophthalmol. 2012, 23, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Bourne, W.M.; Nelson, L.R.; Hodge, D.O. Continued endothelial cell loss ten years after lens implantation. Ophthalmology 1994, 101, 1014–1022. [Google Scholar] [CrossRef]
- Yamazoe, K.; Yamaguchi, T.; Hotta, K.; Satake, Y.; Konomi, K.; Den, S.; Shimazaki, J. Outcomes of cataract surgery in eyes with a low corneal endothelial cell density. J. Cataract Refract. Surg. 2011, 37, 2130–2136. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-C.; Huang, C.-W.; Yeh, L.-K.; Hsiao, F.-C.; Hsueh, Y.-J.; Meir, Y.-J.J.; Chen, K.-J.; Cheng, C.-M.; Wu, W.-C. Accelerated Corneal Endothelial Cell Loss after Phacoemulsification in Patients with Mildly Low Endothelial Cell Density. J. Clin. Med. 2021, 10, 2270. [Google Scholar] [CrossRef]
- Wojcik, K.A.; Kaminska, A.; Blasiak, J.; Szaflik, J.; Szaflik, J.P. Oxidative stress in the pathogenesis of keratoconus and Fuchs endothelial corneal dystrophy. Int. J. Mol. Sci. 2013, 14, 19294–19308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Mehta, G.; Vasiliou, V. Antioxidant defenses in the ocular surface. Ocul. Surf. 2009, 7, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Umapathy, A.; Donaldson, P.; Lim, J. Antioxidant delivery pathways in the anterior eye. Biomed Res. Int. 2013, 2013, 207250. [Google Scholar] [CrossRef] [Green Version]
- Arnal, E.; Peris-Martínez, C.; Menezo, J.L.; Johnsen-Soriano, S.; Romero, F.J. Oxidative stress in keratoconus? Investig. Ophthalmol. Vis. Sci. 2011, 52, 8592–8597. [Google Scholar] [CrossRef] [Green Version]
- Kenney, M.C.; Chwa, M.; Atilano, S.R.; Tran, A.; Carballo, M.; Saghizadeh, M.; Vasiliou, V.; Adachi, W.; Brown, D.J. Increased levels of catalase and cathepsin V/L2 but decreased TIMP-1 in keratoconus corneas: Evidence that oxidative stress plays a role in this disorder. Investig. Ophthalmol. Vis. Sci. 2005, 46, 823–832. [Google Scholar] [CrossRef]
- Behndig, A.; Karlsson, K.; Johansson, B.O.; Brännström, T.; Marklund, S.L. Superoxide dismutase isoenzymes in the normal and diseased human cornea. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2293–2296. [Google Scholar]
- Joseph, R.; Srivastava, O.P.; Pfister, R.R. Differential epithelial and stromal protein profiles in keratoconus and normal human corneas. Exp. Eye Res. 2011, 92, 282–298. [Google Scholar] [CrossRef]
- Gondhowiardjo, T.D.; van Haeringen, N.J.; Völker-Dieben, H.J.; Beekhuis, H.W.; Kok, J.H.; van Rij, G.; Pels, L.; Kijlstra, A. Analysis of corneal aldehyde dehydrogenase patterns in pathologic corneas. Cornea 1993, 12, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Caglayan, M.; Kocamıs, S.I.; Sarac, O.; Tatli Dogan, H.; Kosekahya, P.; Ayan, M.; Cagil, N. Investigation of Heme Oxygenase 2 Enzyme Protein Expression in Keratoconus and Normal Human Corneal Epithelium: An Immunohistochemical Study. Curr. Eye Res. 2019, 44, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Khalimov, A.R.; Khalimova, L.I.; Usubov, E.L. Local Antioxidant Status in Patients with Keratoconus after Ultraviolet Corneal Crosslinking. Ophthalmol. Russ. 2019, 16, 74–78. [Google Scholar] [CrossRef]
- Balmus, I.M.; Alexa, A.I.; Ciuntu, R.E.; Danielescu, C.; Stoica, B.; Cojocaru, S.I.; Ciobica, A.; Cantemir, A. Oxidative stress markers dynamics in keratoconus patients’ tears before and after corneal collagen crosslinking procedure. Exp. Eye Res. 2020, 190, 107897. [Google Scholar] [CrossRef]
- Choy, C.K.; Benzie, I.F.; Cho, P. Ascorbic acid concentration and total antioxidant activity of human tear fluid measured using the FRASC assay. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3293–3298. [Google Scholar]
- Gogia, R.; Richer, S.P.; Rose, R.C. Tear fluid content of electrochemically active components including water soluble antioxidants. Curr. Eye Res. 1998, 17, 257–263. [Google Scholar] [CrossRef]
- Saijyothi, A.V.; Fowjana, J.; Madhumathi, S.; Rajeshwari, M.; Thennarasu, M.; Prema, P.; Angayarkanni, N. Tear fluid small molecular antioxidants profiling shows lowered glutathione in keratoconus. Exp. Eye Res. 2012, 103, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.A.; Pye, D.C.; Willcox, M.D. Levels of lactoferrin, secretory IgA and serum albumin in the tear film of people with keratoconus. Exp. Eye Res. 2012, 96, 132–137. [Google Scholar] [CrossRef]
- Lema, I.; Brea, D.; Rodríguez-González, R.; Díez-Feijoo, E.; Sobrino, T. Proteomic analysis of the tear film in patients with keratoconus. Mol. Vis. 2010, 16, 2055–2061. [Google Scholar] [PubMed]
- Göncü, T.; Akal, A.; Adıbelli, F.M.; Çakmak, S.; Sezen, H.; Yılmaz, Ö.F. Tear Film and Serum Prolidase Activity and Oxidative Stress in Patients with Keratoconus. Cornea 2015, 34, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Kılıç, R.; Cumurcu, T.; Sancaktar, E.; Evliyaoğlu, O.; Sezer, H. Systemic Prolidase Activity and Oxidative Stress in Keratoconus. Curr. Eye Res. 2016, 41, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Toprak, I.; Kucukatay, V.; Yildirim, C.; Kilic-Toprak, E.; Kilic-Erkek, O. Increased systemic oxidative stress in patients with keratoconus. Eye 2014, 28, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Tekin, S.; Seven, E. Assessment of serum catalase, reduced glutathione, and superoxide dismutase activities and malondialdehyde levels in keratoconus patients. Eye 2021. [Google Scholar] [CrossRef]
- Cantemir, A.; Alexa, A.I.; Ciobica, A.; Balmus, I.M.; Antioch, I.; Stoica, B.; Chiselita, D.; Costin, D. Evaluation of Antioxidant Enzymes in Keratoconus. Rev. Chim. 2016, 67, 1538–1541. [Google Scholar]
- Abdul-Maksoud, R.S.; Fouad, R.A.; Elsayed, T.G.; Ibrahem, R.A.; Badawi, A.E. The impact of catalase and glutathione peroxidase-1 genetic polymorphisms on their enzyme activities among Egyptian patients with keratoconus. J. Gene Med. 2020, 22, e3192. [Google Scholar] [CrossRef]
- Ilhan, A.; Altun, S.; Durukan, I.; Yolcu, U.; Erdem, U. The association between genetic polymorphism of glutathione peroxidase 1 (rs1050450) and keratoconus in a Turkish population. Arq. Bras. Oftalmol. 2019, 82, 501–506. [Google Scholar] [CrossRef]
- Yari, D.; Saravani, R.; Saravani, S.; Ebrahimian, K.; Galavi, H.R. Genetic Polymorphisms of Catalase and Glutathione Peroxidase-1 in Keratoconus. Iran. J. Public Health 2018, 47, 1567–1574. [Google Scholar] [PubMed]
- Gulpamuk, B.; Koç, M.; Karatepe, M.S.; Yildiz, A.; Erel, O.; Neselioglu, S.; Yilmazbas, P. Novel Assay Assessment of Oxidative Stress Biomarkers in Patients with Keratoconus: Thiol-Disulfide Homeostasis. Curr. Eye Res. 2017, 42, 1215–1219. [Google Scholar] [CrossRef]
- Zarei-Ghanavati, S.; Yahaghi, B.; Hassanzadeh, S.; Mobarhan, M.G.; Hakimi, H.R.; Eghbali, P. Serum 25-Hydroxyvitamin D, Selenium, Zinc and Copper in Patients with Keratoconus. J. Curr. Ophthalmol. 2020, 32, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Bamdad, S.; Owji, N.; Bolkheir, A. Association Between Advanced Keratoconus and Serum Levels of Zinc, Calcium, Magnesium, Iron, Copper, and Selenium. Cornea 2018, 37, 1306–1310. [Google Scholar] [CrossRef] [PubMed]
- Katta, A.V.; Katkam, R.V.; Geetha, H. Lipid peroxidation and the total antioxidant status in the pathogenesis of age related and diabetic cataracts: A study on the lens and blood. J. Clin. Diagn. Res. 2013, 7, 978–981. [Google Scholar] [CrossRef] [PubMed]
- Babizhayev, M.A. Mitochondria induce oxidative stress, generation of reactive oxygen species and redox state unbalance of the eye lens leading to human cataract formation: Disruption of redox lens organization by phospholipid hydroperoxides as a common basis for cataract disease. Cell Biochem. Funct. 2011, 29, 183–206. [Google Scholar] [PubMed]
- Zhang, Y.; Zhang, L.; Sun, D.; Li, Z.; Wang, L.; Liu, P. Genetic polymorphisms of superoxide dismutases, catalase, and glutathione peroxidase in age-related cataract. Mol. Vis. 2011, 17, 2325–2332. [Google Scholar]
- Wang, X.; Sun, J.; Dang, G.F.; Gao, Y.; Duan, L.; Wu, X.Y. Antioxidant content and cytological examination of aqueous fluid from patients with age-related cataracts at different stages. Genet. Mol. Res. 2015, 14, 6251–6255. [Google Scholar] [CrossRef]
- Elmazar, H.M.; Elmadbouh, I.; Mandour, S.S.; Al Ariny, G.M.; Ibrahim, A.M. Association between cataract progression and ischemia-modified albumin in relation to oxidant-antioxidant profiles in the serum, aqueous humor, and lens. J. Cataract Refract. Surg. 2018, 44, 134–139. [Google Scholar] [CrossRef]
- Yanshole, V.V.; Yanshole, L.V.; Snytnikova, O.A.; Tsentalovich, Y.P. Quantitative metabolomic analysis of changes in the lens and aqueous humor under development of age-related nuclear cataract. Metabolomics 2019, 15, 29. [Google Scholar] [CrossRef]
- Krepler, K.; Schmid, R. Alpha-tocopherol in plasma, red blood cells and lenses with and without cataract. Am. J. Ophthalmol. 2005, 139, 266–270. [Google Scholar] [CrossRef]
- Zorić, L.; Aleksić, P.; Koraćević, D.; Trajković, G. The aqueous humour antioxidative capacity in different types and color of the age-related cataract. Vojn. Pregl. 2005, 62, 909–913. [Google Scholar] [CrossRef]
- Qin, Y.J.; Chan, S.O.; Lin, H.L.; Zhang, Y.Q.; Chen, Y.L.; Niu, Y.Y.; Xie, W.J.; Chu, W.K.; Pang, C.P.; Zhang, H.Y. Elevated level of uric acid in aqueous humour is associated with posterior subcapsular cataract in human lens. Clin. Exp. Ophthalmol. 2020, 48, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Canadananović, V.; Latinović, S.; Barišić, S.; Babić, N.; Jovanović, S. Age-related changes of vitamin C levels in aqueous humour. Vojn. Pregl. 2015, 72, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Selvi, R.; Angayarkanni, N.; Biswas, J.; Ramakrishnan, S. Total antioxidant capacity in Eales’ disease, uveitis & cataract. Indian J. Med. Res. 2011, 134, 83–90. [Google Scholar] [PubMed]
- Zhang, Y.; Jiang, W.; Xie, Z.; Wu, W.; Zhang, D. Vitamin E and risk of age-related cataract: A meta-analysis. Public Health Nutr. 2015, 18, 2804–2814. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Han, J.; Jiang, Y.; Zhang, D. Association of vitamin A and β-carotene with risk for age-related cataract: A meta-analysis. Nutrition 2014, 30, 1113–1121. [Google Scholar] [CrossRef]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [Green Version]
- Eamegdool, S.S.; Sitiwin, E.I.; Cioanca, A.V.; Madigan, M.C. Extracellular matrix and oxidative stress regulate human retinal pigment epithelium growth. Free Radic. Biol. Med. 2020, 146, 357–371. [Google Scholar] [CrossRef]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and its role in age-related macular degeneration. Cell Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [Green Version]
- Erie, J.C.; Good, J.A.; Butz, J.A.; Pulido, J.S. Reduced zinc and copper in the retinal pigment epithelium and choroid in age-related macular degeneration. Am. J. Ophthalmol. 2009, 147, 276–282.e1. [Google Scholar] [CrossRef]
- Armento, A.; Honisch, S.; Panagiotakopoulou, V.; Sonntag, I.; Jacob, A.; Bolz, S.; Kilger, E.; Deleidi, M.; Clark, S.; Ueffing, M. Loss of Complement Factor H impairs antioxidant capacity and energy metabolism of human RPE cells. Sci. Rep. 2020, 10, 10320. [Google Scholar] [CrossRef]
- Arunkumar, R.; Calvo, C.M.; Conrady, C.D.; Bernstein, P.S. What do we know about the macular pigment in AMD: The past, the present, and the future. Eye 2018, 32, 992–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totan, Y.; Yağci, R.; Bardak, Y.; Ozyurt, H.; Kendir, F.; Yilmaz, G.; Sahin, S.; Sahin Tiğ, U. Oxidative macromolecular damage in age-related macular degeneration. Curr. Eye Res. 2009, 34, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Mrowicka, M.; Mrowicki, J.; Szaflik, J.P.; Szaflik, M.; Ulinska, M.; Szaflik, J.; Majsterek, I. Analysis of antioxidative factors related to AMD risk development in the polish patients. Acta Ophthalmol. 2017, 95, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Zafrilla, P.; Losada, M.; Perez, A.; Caravaca, G.; Mulero, J. Biomarkers of oxidative stress in patients with wet age related macular degeneration. J. Nutr. Health Aging 2013, 17, 219–222. [Google Scholar] [CrossRef]
- Gale, C.R.; Hall, N.F.; Phillips, D.I.; Martyn, C.N. Lutein and zeaxanthin status and risk of age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2461–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merle, B.M.J.; Cougnard-Grégoire, A.; Korobelnik, J.F.; Schalch, W.; Etheve, S.; Rougier, M.B.; Féart, C.; Samieri, C.; Delyfer, M.N.; Delcourt, C. Plasma Lutein, a Nutritional Biomarker for Development of Advanced Age-Related Macular Degeneration: The Alienor Study. Nutrients 2021, 13, 2047. [Google Scholar] [CrossRef]
- Arıkan Yorgun, M.; Toklu, Y.; Altınkaynak, H.; Tanrıverdi, B.; Ergin, M.; Biçer, C. A Novel Tool for the Assessment Oxidative Stress in Age-Related Macular Degeneration: Thiol/Disulfide Homeostasis Revisited. Curr. Eye Res. 2016, 41, 1584–1589. [Google Scholar] [CrossRef]
- Nucci, C.; Di Pierro, D.; Varesi, C.; Ciuffoletti, E.; Russo, R.; Gentile, R.; Cedrone, C.; Pinazo Duran, M.D.; Coletta, M.; Mancino, R. Increased malondialdehyde concentration and reduced total antioxidant capacity in aqueous humor and blood samples from patients with glaucoma. Mol. Vis. 2013, 19, 1841–1846. [Google Scholar]
- Tang, B.; Li, S.; Cao, W.; Sun, X. The Association of Oxidative Stress Status with Open-Angle Glaucoma and Exfoliation Glaucoma: A Systematic Review and Meta-Analysis. J. Ophthalmol. 2019, 2019, 1803619. [Google Scholar] [CrossRef] [Green Version]
- Sacca, S.C.; Izzotti, A.; Rossi, P.; Traverso, C. Glaucomatous outflow pathway and oxidative stress. Exp. Eye Res. 2007, 84, 389–399. [Google Scholar] [CrossRef]
- Benoist d’Azy, C.; Pereira, B.; Chiambaretta, F.; Dutheil, F. Oxidative and Anti-Oxidative Stress Markers in Chronic Glaucoma: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0166915. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, M.; Costas-Rodríguez, M.; Lobo, L.; García, M.; González-Iglesias, H.; Pereiro, R.; Vanhaecke, F. Homeostatic alterations related to total antioxidant capacity, elemental concentrations and isotopic compositions in aqueous humor of glaucoma patients. Anal. Bioanal. Chem. 2021, 414, 515–524. [Google Scholar] [CrossRef]
- Ferreira, S.M.; Lerner, S.F.; Brunzini, R.; Evelson, P.A.; Llesuy, S.F. Oxidative stress markers in aqueous humor of glaucoma patients. Am. J. Ophthalmol. 2004, 137, 62–69. [Google Scholar] [CrossRef]
- Saccaà, S.C.; Izzotti, A. Oxidative Stress in Anterior Segment of Primary Open Angle Glaucoma. In Glaucoma—Current Clinical and Research Aspects; Gunvant, P., Ed.; InTech: Rijeka, Croatia, 2011; pp. 1–24. [Google Scholar]
- Li, S.; Shao, M.; Tang, B.; Zhang, A.; Cao, W.; Sun, X. The association between serum uric acid and glaucoma severity in primary angle closure glaucoma: A retrospective case-control study. Oncotarget 2017, 8, 2816–2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Shao, M.; Li, D.; Tang, B.; Cao, W.; Sun, X. Association of serum uric acid levels with primary open-angle glaucoma: A 5-year case-control study. Acta Ophthalmol. 2019, 97, e356–e363. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.E.; Tseng, V.L.; Yu, F.; Caprioli, J.; Coleman, A.L. Association of Dietary Fatty Acid Intake with Glaucoma in the United States. JAMA Ophthalmol. 2018, 136, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuki, K.; Murat, D.; Kimura, I.; Ohtake, Y.; Tsubota, K. Reduced-serum vitamin C and increased uric acid levels in normal-tension glaucoma. Graefes Arch. Clin. Exp. Ophthalmol. 2010, 248, 243–248. [Google Scholar] [CrossRef]
- Li, J.; Xu, F.; Zeng, R.; Gong, H.; Lan, Y. Plasma Homocysteine, Serum Folic Acid, Serum Vitamin B12, Serum Vitamin B6, MTHFR, and Risk of Normal-Tension Glaucoma. J. Glaucoma 2016, 25, e94–e98. [Google Scholar] [CrossRef]
- Ramdas, W.D.; Wolfs, R.C.; Kiefte-de Jong, J.C.; Hofman, A.; de Jong, P.T.; Vingerling, J.R.; Jansonius, N.M. Nutrient intake and risk of open-angle glaucoma: The Rotterdam Study. Eur. J. Epidemiol. 2012, 27, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.P.; Shen, M.Y.; Shen, G.L.; Qi, Q.R.; Sun, X.H. Melatonin concentrations in serum of primary glaucoma patients. Int. J. Ophthalmol. 2018, 11, 1337–1341. [Google Scholar]
- Mas Tur, V.; MacGregor, C.; Jayaswal, R.; O’Brart, D.; Maycock, N. A review of keratoconus: Diagnosis, pathophysiology, and genetics. Surv. Ophthalmol. 2017, 62, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Navel, V.; Malecaze, J.; Pereira, B.; Baker, J.S.; Malecaze, F.; Sapin, V.; Chiambaretta, F.; Dutheil, F. Oxidative and antioxidative stress markers in keratoconus: A systematic review and meta-analysis. Acta Ophthalmol. 2021, 99, e777–e794. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.G.; de Almeida Júnior, G.C.; Teixeira, R.M.; de Mattos, L.C.; Brandão de Mattos, C.C.; Castiglioni, L. Absence of the c.169+50delTAAACAG mutation of SOD1 gene in a sample of keratoconus patients in Brazilian population. BMC Res. Notes 2020, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Shinde, V.; Hu, N.; Mahale, A.; Maiti, G.; Daoud, Y.; Eberhart, C.G.; Maktabi, A.; Jun, A.S.; Al-Swailem, S.A.; Chakravarti, S. RNA sequencing of corneas from two keratoconus patient groups identifies potential biomarkers and decreased NRF2-antioxidant responses. Sci. Rep. 2020, 10, 9907. [Google Scholar] [CrossRef]
- Snytnikova, O.A.; Yanshole, L.V.; Iskakov, I.A.; Yanshole, V.V.; Chernykh, V.V.; Stepakov, D.A.; Novoselov, V.P.; Tsentalovich, Y.P. Quantitative metabolomic analysis of the human cornea and aqueous humor. Metabolomics 2017, 13, 152. [Google Scholar] [CrossRef]
- Tsao, Y.T.; Wu, W.C.; Chen, K.J.; Yeh, L.K.; Hwang, Y.S.; Hsueh, Y.J.; Chen, H.C.; Cheng, C.M. Analysis of aqueous humor total antioxidant capacity and its correlation with corneal endothelial health. Bioeng. Transl. Med. 2021, 6, e10199. [Google Scholar] [CrossRef]
- Haung, W.; Koralewska-Makár, A.; Bauer, B.; Akesson, B. Extracellular glutathione peroxidase and ascorbic acid in aqueous humor and serum of patients operated on for cataract. Clin. Chim. Acta 1997, 261, 117–130. [Google Scholar] [CrossRef]
- Lim, J.C.; Umapathy, A.; Grey, A.C.; Vaghefi, E.; Donaldson, P.J. Novel roles for the lens in preserving overall ocular health. Exp. Eye Res. 2017, 156, 117–123. [Google Scholar] [CrossRef]
- George, S.; Jyothi, M.; Mathew, B.; Shashidhar, S. Changes in glutathione, glutathione-linked enzymes and hexose monophosphate shunt enzymes in senile cataract. Indian J. Physiol. Pharmacol. 2003, 47, 191–196. [Google Scholar] [PubMed]
- Chen, X.; Meng, Y.; Li, J.; She, H.; Zhao, L.; Zhang, J.; Peng, Y.; Shang, K.; Zhang, Y.; Gu, X.; et al. Serum uric acid concentration is associated with hypertensive retinopathy in hypertensive chinese adults. BMC Ophthalmol. 2017, 17, 83. [Google Scholar] [CrossRef] [Green Version]
- Conrady, C.D.; Bell, J.P.; Besch, B.M.; Gorusupudi, A.; Farnsworth, K.; Ermakov, I.; Sharifzadeh, M.; Ermakova, M.; Gellermann, W.; Bernstein, P.S. Correlations Between Macular, Skin, and Serum Carotenoids. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3616–3627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engin, K.N.; Yemişci, B.; Yiğit, U.; Ağaçhan, A.; Coşkun, C. Variability of serum oxidative stress biomarkers relative to biochemical data and clinical parameters of glaucoma patients. Mol. Vis. 2010, 16, 1260–1271. [Google Scholar] [PubMed]
- Li, S.; Shao, M.; Li, Y.; Li, X.; Wan, Y.; Sun, X.; Cao, W. Relationship between Oxidative Stress Biomarkers and Visual Field Progression in Patients with Primary Angle Closure Glaucoma. Oxidative Med. Cell. Longev. 2020, 2020, 2701539. [Google Scholar] [CrossRef] [PubMed]
- Barros, P.S.; Safatle, A.M.; Queiroz, L.; Silva, V.V.; Barros, S.B. Blood and aqueous humour antioxidants in cataractous poodles. Can. J. Ophthalmol. 2004, 39, 19–24. [Google Scholar] [CrossRef]
- Brubaker, R.F.; Bourne, W.M.; Bachman, L.A.; McLaren, J.W. Ascorbic Acid Content of Human Corneal Epithelium. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1681–1683. [Google Scholar]
- Mancino, R.; Di Pierro, D.; Varesi, C.; Cerulli, A.; Feraco, A.; Cedrone, C.; Pinazo-Duran, M.D.; Coletta, M.; Nucci, C. Lipid peroxidation and total antioxidant capacity in vitreous, aqueous humor, and blood samples from patients with diabetic retinopathy. Mol. Vis. 2011, 17, 1298–1304. [Google Scholar]
- Beyazyıldız, E.; Cankaya, A.B.; Ergan, E.; Anayol, M.A.; Ozdamar, Y.; Sezer, S.; Tırhış, M.H.; Yılmazbaş, P.; Oztürk, F. Changes of total antioxidant capacity and total oxidant status of aqueous humor in diabetes patients and correlations with diabetic retinopathy. Int. J. Ophthalmol. 2013, 6, 531–536. [Google Scholar]
- Kirboga, K.; Ozec, A.V.; Kosker, M.; Dursun, A.; Toker, M.I.; Aydin, H.; Erdogan, H.; Topalkara, A.; Arici, M.K. The Association between Diabetic Retinopathy and Levels of Ischemia-Modified Albumin, Total Thiol, Total Antioxidant Capacity, and Total Oxidative Stress in Serum and Aqueous Humor. J. Ophthalmol. 2014, 2014, 820853. [Google Scholar] [CrossRef] [Green Version]
- Bozkurt, E.; Çakır, B.; Çelik, E.; Doğan, E.; Uçak, T.; Alagöz, G. Correlation of the aqueous humor total antioxidant capacity, total oxidant status, and levels of IL-6 and VEGF with diabetic retinopathy status. Arq. Bras. Oftalmol. 2019, 82, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Jacques, P.F.; Nadler, D.; Morrow, F.; Sulsky, S.I.; Shepard, D. Relationship in humans between ascorbic acid consumption and levels of total and reduced ascorbic acid in lens, aqueous humor, and plasma. Curr. Eye Res. 1991, 10, 751–759. [Google Scholar] [CrossRef]
- Takano, S.; Ishiwata, S.; Nakazawa, M.; Mizugaki, M.; Tamai, M. Determination of ascorbic acid in human vitreous humor by high-performance liquid chromatography with UV detection. Curr. Eye Res. 1997, 16, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Jabbehdari, S.; Handa, J.T. Oxidative stress as a therapeutic target for the prevention and treatment of early age-related macular degeneration. Surv. Ophthalmol. 2021, 66, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, H.; Alany, R.G.; Pierscionek, B. Age-related cataract and drug therapy: Opportunities and challenges for topical antioxidant delivery to the lens. J. Pharm. Pharmacol. 2015, 67, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Eghtedari, Y.; Oh, L.J.; Girolamo, N.D.; Watson, S.L. The role of topical N-acetylcysteine in ocular therapeutics. Surv. Ophthalmol. 2021; online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Harada, C.; Noro, T.; Kimura, A.; Guo, X.; Namekata, K.; Nakano, T.; Harada, T. Suppression of Oxidative Stress as Potential Therapeutic Approach for Normal Tension Glaucoma. Antioxidants 2020, 9, 874. [Google Scholar] [CrossRef]
- Age-Related Eye Disease Study Research Group. The Age-Related Eye Disease Study (AREDS): Design implications. AREDS report no. 1. Control Clin. Trials 1999, 20, 573–600. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0197245699000318 (accessed on 15 December 2021). [CrossRef]
- Evans, J. Antioxidant supplements to prevent or slow down the progression of AMD: A systematic review and meta-analysis. Eye 2008, 22, 751–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for preventing age-related macular degeneration. Cochrane Database Syst. Rev. 2017, 7, Cd000253. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Reiss, G.R.; Werness, P.G.; Zollman, P.E.; Brubaker, R.F. Ascorbic Acid Levels in the Aqueous Humor of Nocturnal and Diurnal Mammals. Arch. Ophthalmol. 1986, 104, 753–755. [Google Scholar] [CrossRef]
- Ringvold, A. The Significance of Ascorbate in the Aqueous Humour Protection Against UV-A and UV-B. Exp. Eye Res. 1996, 62, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Zhu, B.-Z.; Frei, B. Potential Antiatherogenic Mechanisms of Ascorbate (Vitamin C) and α-Tocopherol (Vitamin E). Circ. Res. 2000, 87, 349–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, R.L. The effect of ascorbate on wound healing. Int. Ophthalmol. Clin. 2000, 40, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Gujral, G.S.; Askari, S.N.; Ahmad, S.; Zakir, S.M.; Saluja, K. Topical vitamin C, vitamin E, and acetylcysteine as corneal wound healing agents: A comparative study. Indian J. Ophthalmol. 2020, 68, 2935–2939. [Google Scholar]
- Padua, I.R.M.; Valdetaro, G.P.; Lima, T.B.; Kobashigawa, K.K.; Silva, P.E.S.; Aldrovani, M.; Padua, P.P.M.; Laus, J.L. Effects of intracameral ascorbic acid on the corneal endothelium of dogs undergoing phacoemulsification. Vet. Ophthalmol. 2018, 21, 151–159. [Google Scholar] [CrossRef]
- Lee, C.Y.; Chen, H.T.; Hsueh, Y.J.; Chen, H.C.; Huang, C.C.; Meir, Y.J.; Cheng, C.M.; Wu, W.C. Perioperative topical ascorbic acid for the prevention of phacoemulsification-related corneal endothelial damage: Two case reports and review of literature. World J. Clin. Cases 2019, 7, 642–649. [Google Scholar] [CrossRef]
- Kaur, J.; Kukreja, S.; Kaur, A.; Malhotra, N.; Kaur, R. The oxidative stress in cataract patients. J. Clin. Diagn. Res. 2012, 6, 1629. [Google Scholar] [CrossRef]
- Ravindran, R.D.; Vashist, P.; Gupta, S.K.; Young, I.S.; Maraini, G.; Camparini, M.; Jayanthi, R.; John, N.; Fitzpatrick, K.E.; Chakravarthy, U. Inverse association of vitamin C with cataract in older people in India. Ophthalmology 2011, 118, 1958–1965.e2. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y.; Hashizume, K.; Kishimoto, S.; Tezuka, Y.; Nishigori, H.; Yamamoto, N.; Kondo, Y.; Maruyama, N.; Ishigami, A.; Kurosaka, D. Effect of vitamin C depletion on UVR-B induced cataract in SMP30/GNL knockout mice. Exp. Eye Res. 2012, 94, 85–89. [Google Scholar] [CrossRef]
- Mathew, M.C.; Ervin, A.M.; Tao, J.; Davis, R.M. Antioxidant vitamin supplementation for preventing and slowing the progression of age-related cataract. Cochrane Database Syst. Rev. 2012, 6, CD004567. [Google Scholar] [CrossRef] [Green Version]
- Thiagarajan, R.; Manikandan, R. Antioxidants and cataract. Free. Radic. Res. 2013, 47, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.-H.; Jing, C.-X.; Pan, H.-W. Association of blood antioxidants and vitamins with risk of age-related cataract: A meta-analysis of observational studies. Am. J. Clin. Nutr. 2013, 98, 778–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Thomas, F.; Lang, J.C.; Chaum, E. Modulation of oxidative stress responses in the human retinal pigment epithelium following treatment with vitamin C. J. Cell. Physiol. 2011, 226, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Sant, D.W.; Camarena, V.; Mustafi, S.; Li, Y.; Wilkes, Z.; Van Booven, D.; Wen, R.; Wang, G. Ascorbate Suppresses VEGF Expression in Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3608–3618. [Google Scholar] [CrossRef] [Green Version]
- Ramdas, W.D.; Schouten, J.; Webers, C.A.B. The Effect of Vitamins on Glaucoma: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 359. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, F.C.; Chen, H.T.; Chen, K.J.; Hsueh, Y.J.; Meir, Y.J.; Lu, T.T.; Cheng, C.M.; Wu, W.C.; Chen, H.C. Accelerated corneal endothelial cell loss in two patients with granulomatosis with polyangiitis following phacoemulsification. BMC Ophthalmol. 2020, 20, 480. [Google Scholar] [CrossRef]
- Hsueh, Y.J.; Meir, Y.J.; Lai, J.Y.; Huang, C.C.; Lu, T.T.; Ma, D.H.; Cheng, C.M.; Wu, W.C.; Chen, H.C. Ascorbic acid ameliorates corneal endothelial dysfunction and enhances cell proliferation via the noncanonical GLUT1-ERK axis. Biomed. Pharmacother. 2021, 144, 112306. [Google Scholar] [CrossRef]
- Shen, H.; Ding, C.; Yuan, S.; Pan, T.; Li, D.; Li, H.; Huang, B.; Liu, Q. Vitamin C- and Valproic Acid-Induced Fetal RPE Stem-like Cells Recover Retinal Degeneration via Regulating SOX2. Mol. Ther. 2020, 28, 1645–1657. [Google Scholar] [CrossRef]
- Shima, N.; Kimoto, M.; Yamaguchi, M.; Yamagami, S. Increased proliferation and replicative lifespan of isolated human corneal endothelial cells with L-ascorbic acid 2-phosphate. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8711–8717. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.M.; Tsai, J.L.; Lin, S.D.; Lai, C.S.; Chang, C.C. Accelerated growth and prolonged lifespan of adipose tissue-derived human mesenchymal stem cells in a medium using reduced calcium and antioxidants. Stem Cells Dev. 2005, 14, 92–102. [Google Scholar] [CrossRef]
- Liu, T.M.; Yildirim, E.D.; Li, P.; Fang, H.T.; Denslin, V.; Kumar, V.; Loh, Y.H.; Lee, E.H.; Cool, S.M.; Teh, B.T.; et al. Ascorbate and Iron Are Required for the Specification and Long-Term Self-Renewal of Human Skeletal Mesenchymal Stromal Cells. Stem Cell Rep. 2020, 14, 210–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.H.; Kao, A.P.; Chang, C.C.; Lin, T.C.; Kuo, T.C. Lifespan Extension and Sustained Expression of Stem Cell Phenotype of Human Breast Epithelial Stem Cells in a Medium with Antioxidants. Stem Cells Int. 2016, 2016, 4591310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, S.; Hayashi, R.; Soma, T.; Kageyama, T.; Duncan, T.; Tsujikawa, M.; Nishida, K. Identification and potential application of human corneal endothelial progenitor cells. Stem Cells Dev. 2014, 23, 2190–2201. [Google Scholar] [CrossRef] [PubMed]
- Muz, O.E.; Orhan, C.; Erten, F.; Tuzcu, M.; Ozercan, I.H.; Singh, P.; Morde, A.; Padigaru, M.; Rai, D.; Sahin, K. A Novel Integrated Active Herbal Formulation Ameliorates Dry Eye Syndrome by Inhibiting Inflammation and Oxidative Stress and Enhancing Glycosylated Phosphoproteins in Rats. Pharmaceuticals 2020, 13, 295. [Google Scholar] [CrossRef] [PubMed]
- van Asten, F.; Chiu, C.Y.; Agrón, E.; Clemons, T.E.; Ratnapriya, R.; Swaroop, A.; Klein, M.L.; Fan, R.; Chew, E.Y. No CFH or ARMS2 Interaction with Omega-3 Fatty Acids, Low versus High Zinc, or β-Carotene versus Lutein and Zeaxanthin on Progression of Age-Related Macular Degeneration in the Age-Related Eye Disease Study 2: Age-Related Eye Disease Study 2 Report No. 18. Ophthalmology 2019, 126, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Lutein + zeaxanthin and omega-3 fatty acids for age-related macular degeneration: The Age-Related Eye Disease Study 2 (AREDS2) randomized clinical trial. JAMA 2013, 309, 2005–2015. [CrossRef] [PubMed]
- Lem, D.W.; Davey, P.G.; Gierhart, D.L.; Rosen, R.B. A Systematic Review of Carotenoids in the Management of Age-Related Macular Degeneration. Antioxidants 2021, 10, 1255. [Google Scholar] [CrossRef]
- Lem, D.W.; Gierhart, D.L.; Davey, P.G. A Systematic Review of Carotenoids in the Management of Diabetic Retinopathy. Nutrients 2021, 13, 2441. [Google Scholar] [CrossRef]
- Fitzpatrick, N.; Chachay, V.; Bowtell, J.; Jackman, S.; Capra, S.; Shore, A.; Briskey, D. An appraisal of trials investigating the effects on macular pigment optical density of lutein and zeaxanthin dietary interventions: A narrative review. Nutr. Rev. 2021; online ahead of print. [Google Scholar] [CrossRef]
- García-Layana, A.; Recalde, S.; Hernandez, M.; Abraldes, M.J.; Nascimento, J.; Hernández-Galilea, E.; Olmedilla-Alonso, B.; Escobar-Barranco, J.J.; Zapata, M.A.; Silva, R.; et al. A Randomized Study of Nutritional Supplementation in Patients with Unilateral Wet Age-Related Macular Degeneration. Nutrients 2021, 13, 1253. [Google Scholar] [CrossRef]
- Radkar, P.; Lakshmanan, P.S.; Mary, J.J.; Chaudhary, S.; Durairaj, S.K. A Novel Multi-Ingredient Supplement Reduces Inflammation of the Eye and Improves Production and Quality of Tears in Humans. Ophthalmol. Ther. 2021, 10, 581–599. [Google Scholar] [CrossRef] [PubMed]
- Oppenheimer, A. Turmeric (Curcumin) in Biliary Diseases. Lancet 1937, 229, 619–621. [Google Scholar] [CrossRef]
- Strimpakos, A.S.; Sharma, R.A. Curcumin: Preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid. Redox Signal. 2008, 10, 511–545. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, Y.W.; Hsu, S.K.; Chen, J.Y.; Lin, I.L.; Chen, K.J.; Lee, P.Y.; Ng, H.S.; Chiu, C.C.; Cheng, K.C. Curcumin Metabolite Tetrahydrocurcumin in the Treatment of Eye Diseases. Int. J. Mol. Sci. 2020, 22, 212. [Google Scholar] [CrossRef] [PubMed]
- Li, P.C.; Chen, S.C.; Hsueh, Y.J.; Shen, Y.C.; Tsai, M.Y.; Hsu, L.W.; Yeh, C.K.; Chen, H.C.; Huang, C.C. Gelatin scaffold with multifunctional curcumin-loaded lipid-PLGA hybrid microparticles for regenerating corneal endothelium. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 120, 111753. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, N.; Guha, R.; Chowdhury, S.; Nandi, S.; Konar, A.; Hazra, S. Curcumin nanoparticles inhibit corneal neovascularization. J. Mol. Med. 2015, 93, 1095–1106. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Li, J.M.; Qin, L. Suppression of corneal neovascularization by curcumin via inhibition of Wnt/β-catenin pathway activation. Int. J. Ophthalmol. 2017, 10, 1791–1797. [Google Scholar]
- Guo, C.; Li, M.; Qi, X.; Lin, G.; Cui, F.; Li, F.; Wu, X. Intranasal delivery of nanomicelle curcumin promotes corneal epithelial wound healing in streptozotocin-induced diabetic mice. Sci. Rep. 2016, 6, 29753. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.H.; Ko, Y.C.; Chang, Y.F.; Huang, S.H.; Liu, C.J. Thermosensitive chitosan-gelatin-based hydrogel containing curcumin-loaded nanoparticles and latanoprost as a dual-drug delivery system for glaucoma treatment. Exp. Eye Res. 2019, 179, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Munia, I.; Gafray, L.; Bringer, M.A.; Goldschmidt, P.; Proukhnitzky, L.; Jacquemot, N.; Cercy, C.; Ramchani Ben Otman, K.; Errera, M.H.; Ranchon-Cole, I. Cytoprotective Effects of Natural Highly Bio-Available Vegetable Derivatives on Human-Derived Retinal Cells. Nutrients 2020, 12, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muangnoi, C.; Sharif, U.; Ratnatilaka Na Bhuket, P.; Rojsitthisak, P.; Paraoan, L. Protective Effects of Curcumin Ester Prodrug, Curcumin Diethyl Disuccinate against H(2)O(2)-Induced Oxidative Stress in Human Retinal Pigment Epithelial Cells: Potential Therapeutic Avenues for Age-Related Macular Degeneration. Int. J. Mol. Sci. 2019, 20, 3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allegrini, D.; Raimondi, R.; Angi, M.; Ricciardelli, G.; Montericcio, A.; Borgia, A.; Romano, M.R. Curcuma-Based Nutritional Supplement in Patients with Neovascular Age-Related Macular Degeneration. J. Med. Food 2021, 24, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Allegri, P.; Mastromarino, A.; Neri, P. Management of chronic anterior uveitis relapses: Efficacy of oral phospholipidic curcumin treatment. Long-term follow-up. Clin. Ophthalmol. 2010, 4, 1201–1206. [Google Scholar] [PubMed] [Green Version]
- Miserocchi, E.; Giuffrè, C.; Cicinelli, M.V.; Marchese, A.; Gattinara, M.; Modorati, G.; Bandello, F. Oral phospholipidic curcumin in juvenile idiopathic arthritis-associated uveitis. Eur. J. Ophthalmol. 2020, 30, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Mazzolani, F.; Togni, S. Oral administration of a curcumin-phospholipid delivery system for the treatment of central serous chorioretinopathy: A 12-month follow-up study. Clin. Ophthalmol. 2013, 7, 939–945. [Google Scholar]
- Nebbioso, M.; Franzone, F.; Greco, A.; Gharbiya, M.; Bonfiglio, V.; Polimeni, A. Recent Advances and Disputes about Curcumin in Retinal Diseases. Clin. Ophthalmol. 2021, 15, 2553–2571. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Sample Source | Total Antioxidant Capacity/References (↓): Decreased (--): Not Significantly Different | Antioxidant Biomarkers * | References | |
|---|---|---|---|---|---|
| Enzymatic | Nonenzymatic | ||||
| Keratoconus | Cornea | (↓) [121] | SOD Catalase Quinone ALDH HO-2 | Glutathione | [63,122,123,124,125,126] |
| Tear film | (↓) [127] | SOD GPx | Uric acid Glutathione Tyrosine Lactoferrin | [128,129,130,131,132,133] | |
| Serum | (--) [134,135,136] | SOD Catalase GPx | Glutathione Thiol Copper Zinc Selenium | [137,138,139,140,141,142,143,144] | |
| Senile cataract | Lens | (↓) [145] | SOD GPx GR | Ascorbic acid Glutathione Ergothioneine α-tocopherol | [145,146,147,148,149,150,151] |
| Aqueous humor | (↓) [152] | -- | Ascorbic acid Uric acid Glutathione Ergothioneine | [150,153,154] | |
| Serum | (↓) [147,155] | SOD Catalase | Ascorbic acid Glutathione Tocopherol β-carotene | [145,155,156,157] | |
| Age-related macular degeneration | Retina | (--) | SOD Catalase PRDX3 GPx | Lutein Zeaxanthin | [158,159,160] |
| Serum | (↓) [161] | SOD Catalase GPx | Zeaxanthin Lutein Thiol | [162,163,164,165,166,167,168,169] | |
| Glaucoma | Aqueous humor | (↓) [170] (--) [171] | SOD GPx Catalase GST | Zinc | [170,171,172,173,174] |
| Serum | (↓) [170,171,173,175] | SOD | Uric acid Ascorbic acid Melatonin | [170,173,174,175,176,177,178,179,180,181,182,183] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsueh, Y.-J.; Chen, Y.-N.; Tsao, Y.-T.; Cheng, C.-M.; Wu, W.-C.; Chen, H.-C. The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases. Int. J. Mol. Sci. 2022, 23, 1255. https://doi.org/10.3390/ijms23031255
Hsueh Y-J, Chen Y-N, Tsao Y-T, Cheng C-M, Wu W-C, Chen H-C. The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases. International Journal of Molecular Sciences. 2022; 23(3):1255. https://doi.org/10.3390/ijms23031255
Chicago/Turabian StyleHsueh, Yi-Jen, Yen-Ning Chen, Yu-Ting Tsao, Chao-Min Cheng, Wei-Chi Wu, and Hung-Chi Chen. 2022. "The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases" International Journal of Molecular Sciences 23, no. 3: 1255. https://doi.org/10.3390/ijms23031255