
Genome-Wide Identification of the CER Gene Family and Significant Features in Climate Adaptation of Castanea mollissima


Abstract
:1. Introduction
2. Results
2.1. Analysis of the Thickness of the Cuticle in Wild Chinese Chestnut Samples Collected from Northern and Southern China
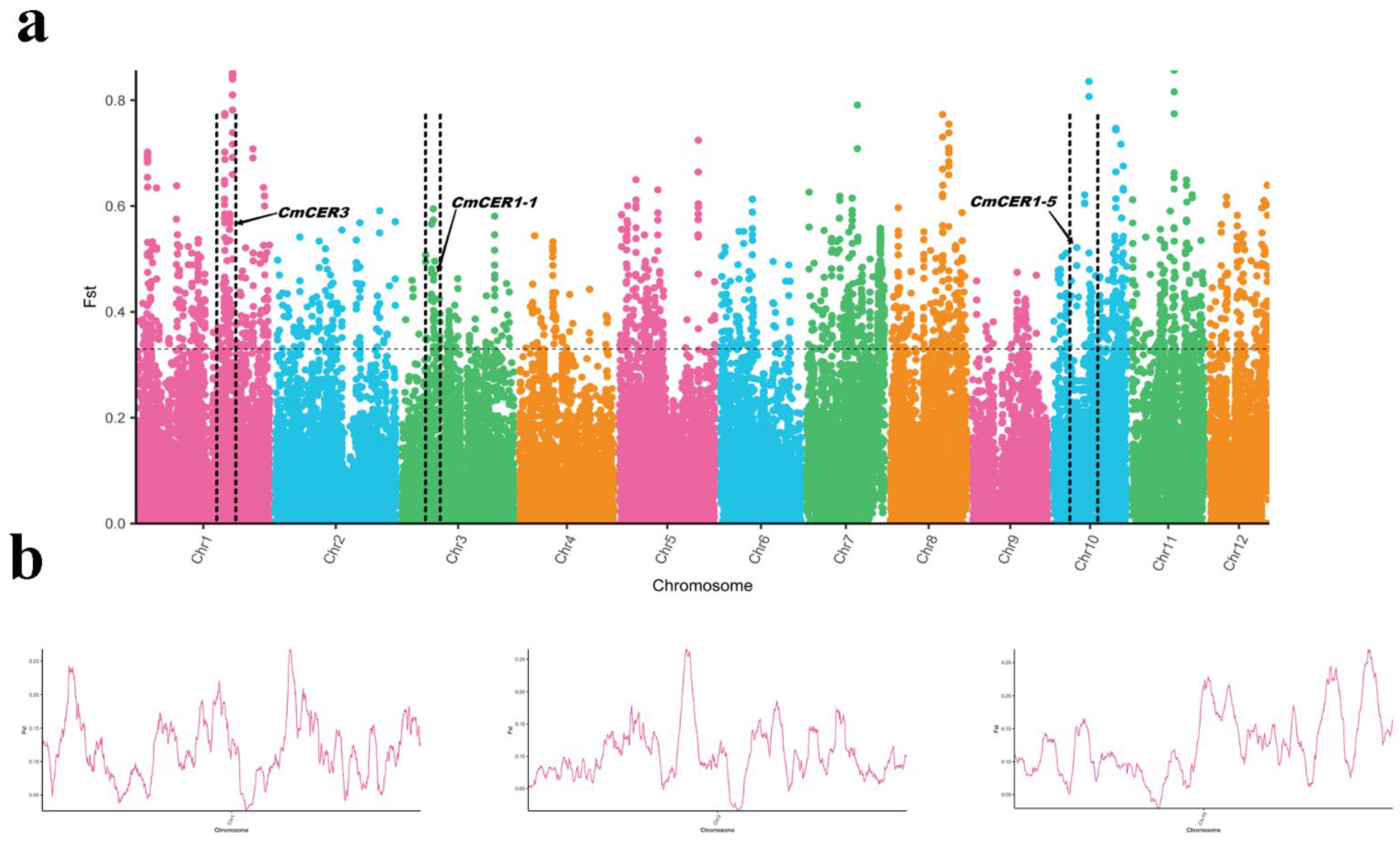
2.2. Genome-Wide Selective Sweep Analysis between the Populations of Wild Chinese Chestnut Collected from Northern and Southern China
2.3. Identification of CmCER Family Genes in Chinese Chestnut
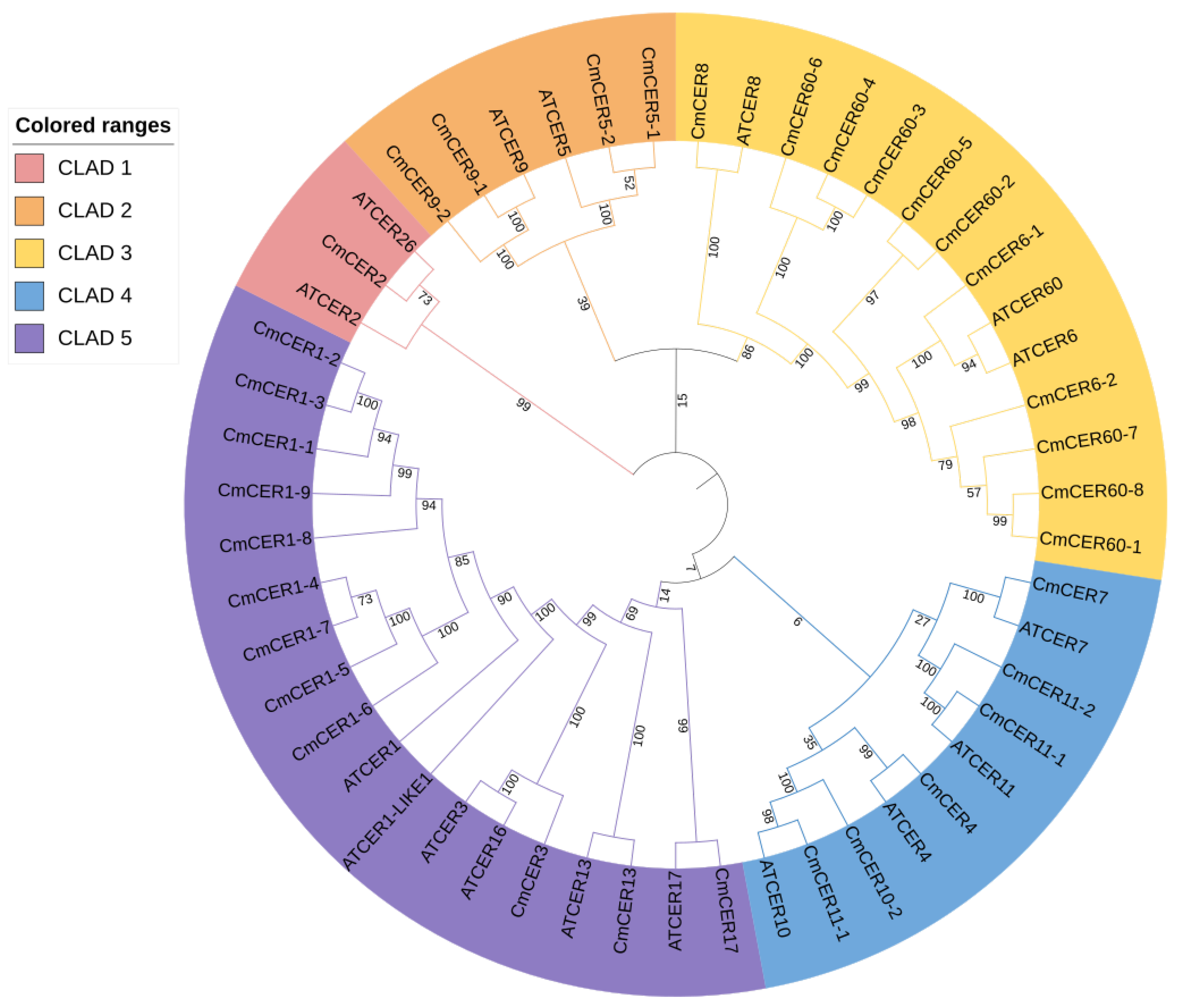
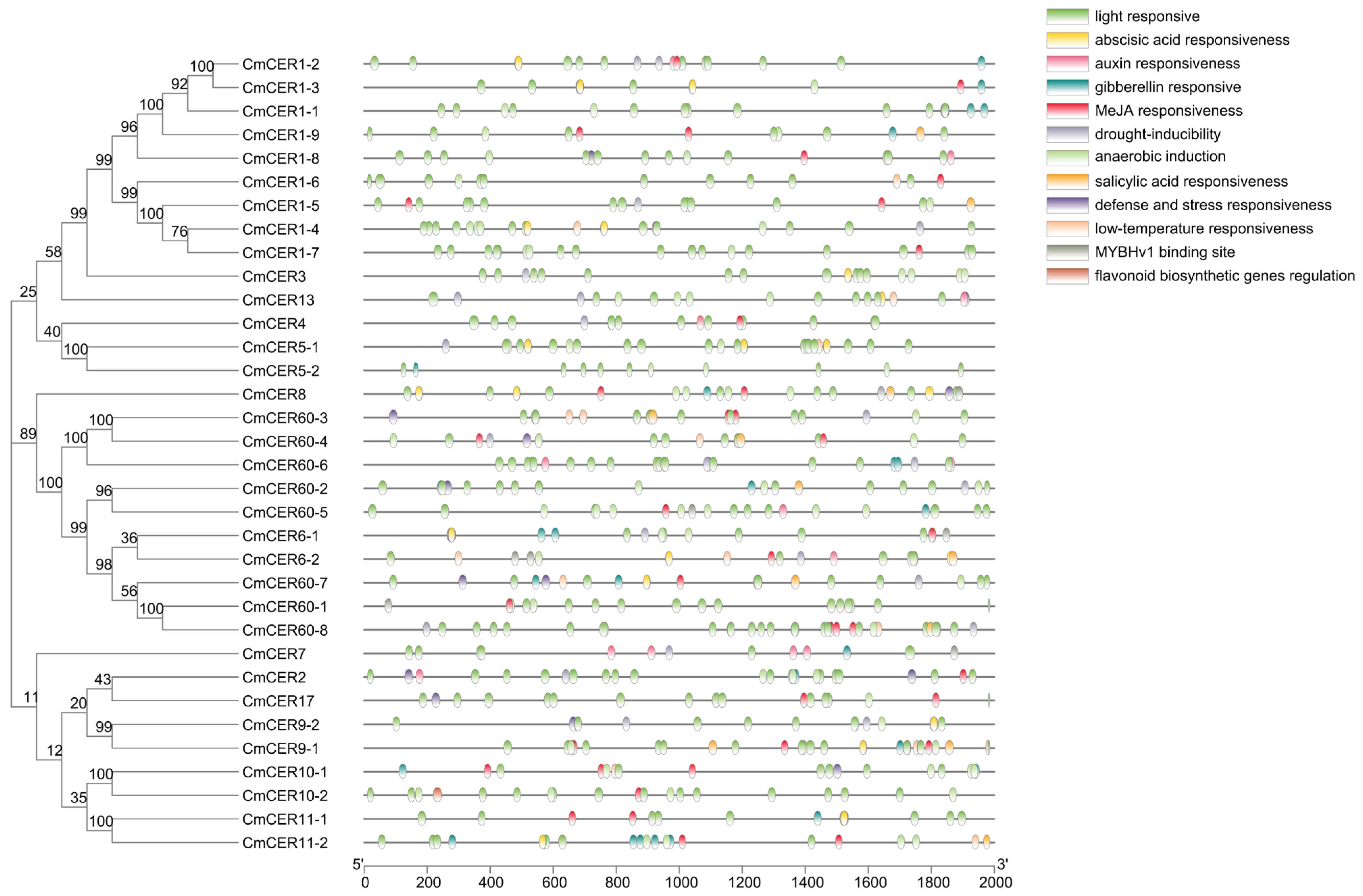
2.4. Phylogenetic Analysis of CmCER Proteins in Chinese Chestnut
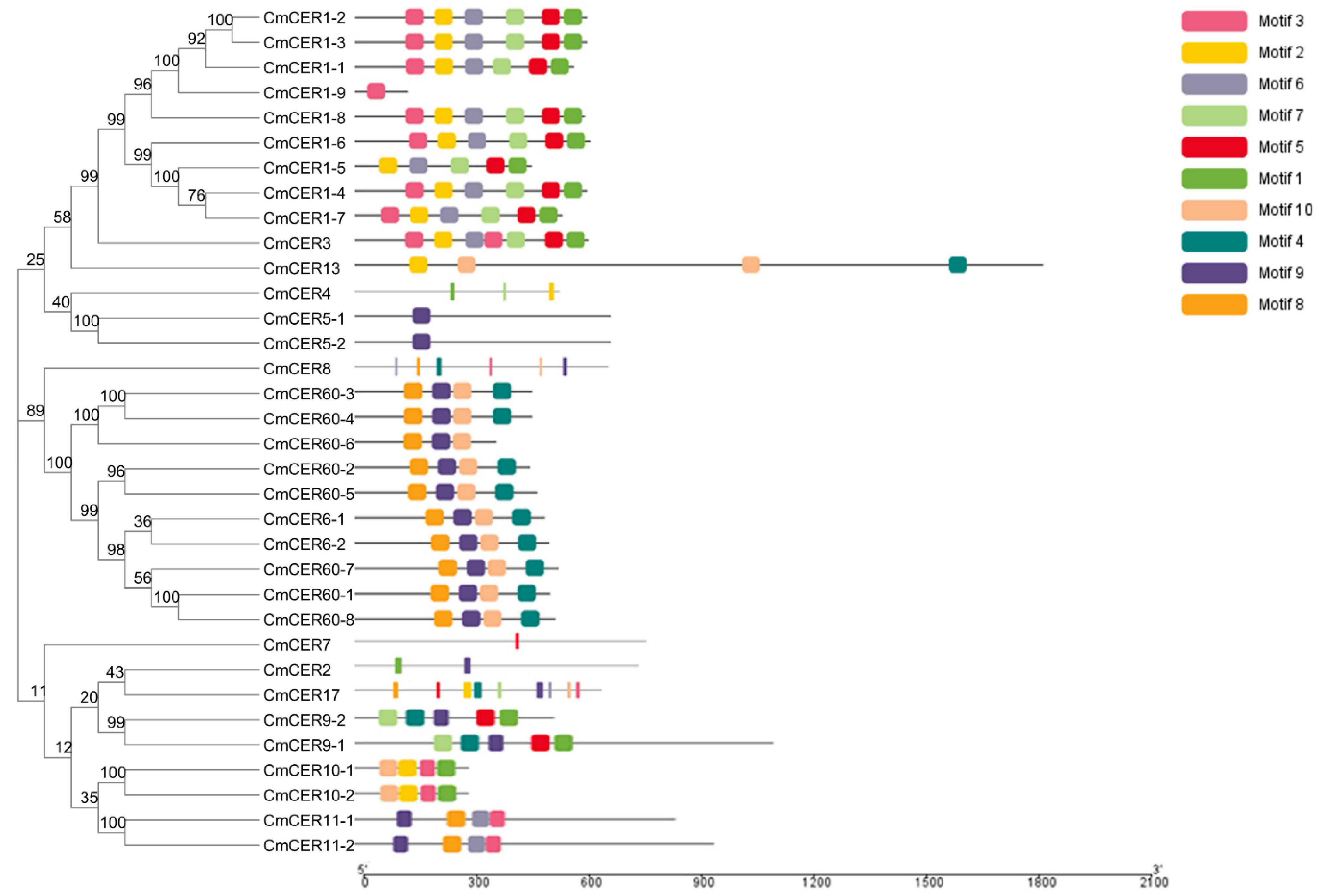
2.5. Analysis of Gene Motifs of CmCER Gene Family in Chinese Chestnut
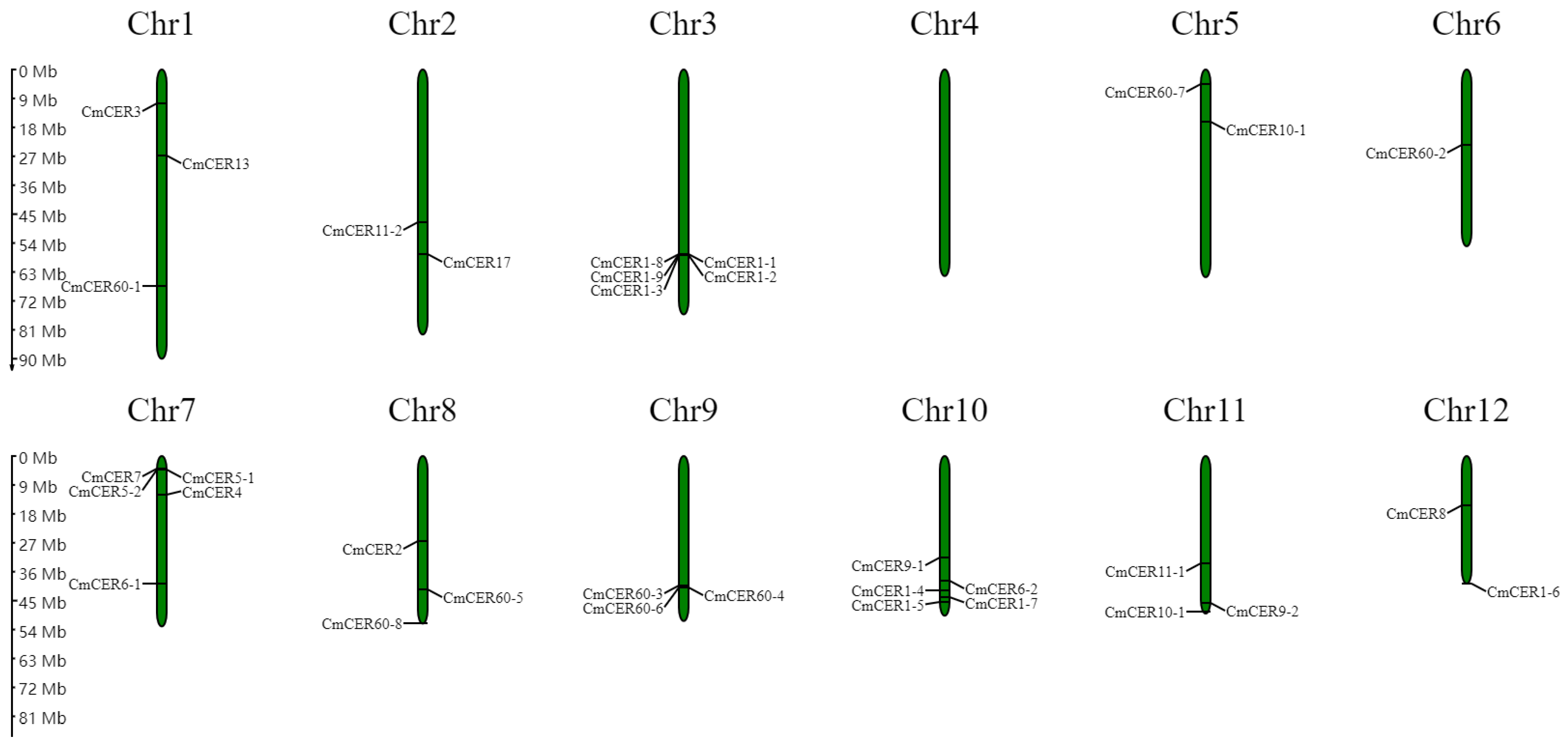
2.6. Chromosomal Localization of CmCER Gene in Chinese Chestnut
2.7. Chromosomal Localization of CmCER Gene in Chinese Chestnut
2.8. Changes of CmCER Gene Expression Level in Chinese Chestnut Seedlings under Drought Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Thickness Statistics of Wax on the Surface of Leaves of Wild and Cultivated Chestnut Species
4.3. Sample Sequencing
4.4. Selective Sweep
4.5. Identification of CER Gene Family Members and Analysis of Physicochemical Properties of Protein Sequences in Chinese Chestnut
4.6. Phylogenetic Analysis
4.7. Gene Structure and Motif Analysis
4.8. Chromosome Localization and Cis-Acting Element Analysis
4.9. qRT-PCR Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dai, A. Increasing Drought under Global Warming in Observations and Models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Zhang, Q.; Han, L.; Jia, J.; Song, L.; Wang, J. Management of Drought Risk under Global Warming. Appl. Clim. 2016, 125, 187–196. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER Encoding an AP2 Domain Transcription Factor Plays a Key Role in Developmental Change from Vegetative to Embryonic Phase. Plant Mol. Biol. 2010, 73, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.; Zeeman, S.C. Formation of Starch in Plant Cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef] [Green Version]
- Yeats, T.H.; Rose, J.K.C. The Formation and Function of Plant Cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuels, L.; Kunst, L.; Jetter, R. Sealing Plant Surfaces: Cuticular Wax Formation by Epidermal Cells. Annu. Rev. Plant Biol. 2008, 59, 683–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad Ahmad, H.; Wang, X.; Fiaz, S.; Mahmood-Ur-Rahman; Azhar Nadeem, M.; Aslam Khan, S.; Ahmar, S.; Azeem, F.; Shaheen, T.; Mora-Poblete, F. Comprehensive Genomics and Expression Analysis of Eceriferum (CER) Genes in Sunflower (Helianthus Annuus). Saudi J. Biol. Sci. 2021, 28, 6884–6896. [Google Scholar] [CrossRef]
- Li, N.; Li, X.Z.; Song, Y.Q.; Yang, S.T.; Li, L.L. Genome-Wide Identification, Characterization, and Expression Profiling of the ECERIFERUM (CER) Gene Family in Ziziphus Jujube. Russ. J. Plant Physiol. 2021, 68, 828–837. [Google Scholar] [CrossRef]
- Qi, C.; Jiang, H.; Zhao, X.; Mao, K.; Liu, H.; Li, Y.; Hao, Y. The Characterization, Authentication, and Gene Expression Pattern of the MdCER Family in Malus Domestica. Hortic. Plant J. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Rizwan, H.M.; Waheed, A.; Ma, S.; Li, J.; Arshad, M.B.; Irshad, M.; Li, B.; Yang, X.; Ali, A.; Ahmed, M.A.A.; et al. Comprehensive Genome-Wide Identification and Expression Profiling of Eceriferum (CER) Gene Family in Passion Fruit (Passiflora Edulis) Under Fusarium Kyushuense and Drought Stress Conditions. Front. Plant Sci. 2022, 13, 898307. [Google Scholar] [CrossRef]
- Aarts, M.G.M. Molecular Characterization of the CER1 Gene of Arabidopsis Lnvolved in Epicuticular Wax Biosynthesis and Pollen Fertility. Plant Cell 1995, 7, 2115–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, A.; Joubès, J. Arabidopsis Cuticular Waxes: Advances in Synthesis, Export and Regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, T.; Li, S.; Zhao, H.; Zhao, S.; Ma, C.; Jenks, M.A.; Lü, S. CER16 Inhibits Post-Transcriptional Gene Silencing of CER3 to Regulate Alkane Biosynthesis. Plant Physiol. 2020, 182, 1211–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Nikolau, B.J. Cloning and Characterization of CER2, an Arabidopsis Gene That Affects Cuticular Wax Accumulation. Plant Cell 1996, 8, 1291–1304. [Google Scholar] [CrossRef]
- Xia, Y.; Nikolau, B.J.; Schnable, P.S. Developmental and Hormonal Regulation of the Arabidopsis CER2 Gene That Codes for a Nuclear-Localized Protein Required for the Normal Accumulation of Cuticular Waxes. Plant Physiol. 1997, 115, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Haslam, T.M.; Mañas-Fernández, A.; Zhao, L.; Kunst, L. Arabidopsis ECERIFERUM2 Is a Component of the Fatty Acid Elongation Machinery Required for Fatty Acid Extension to Exceptional Lengths. Plant Physiol. 2012, 160, 1164–1174. [Google Scholar] [CrossRef] [Green Version]
- Haslam, T.M.; Haslam, R.; Thoraval, D.; Pascal, S.; Delude, C.; Domergue, F.; Fernández, A.M.; Beaudoin, F.; Napier, J.A.; Kunst, L.; et al. ECERIFERUM2-LIKE Proteins Have Unique Biochemical and Physiological Functions in Very-Long-Chain Fatty Acid Elongation. Plant Physiol. 2015, 167, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Pascal, S.; Bernard, A.; Sorel, M.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Domergue, F.; Joubès, J. The Arabidopsis Cer26 Mutant, like the Cer2 Mutant, Is Specifically Affected in the Very Long Chain Fatty Acid Elongation Process. Plant J. 2013, 73, 733–746. [Google Scholar] [CrossRef] [Green Version]
- Rowland, O.; Zheng, H.; Hepworth, S.R.; Lam, P.; Jetter, R.; Kunst, L. CER4 Encodes an Alcohol-Forming Fatty Acyl-Coenzyme A Reductase Involved in Cuticular Wax Production in Arabidopsis. Plant Physiol. 2006, 142, 866–877. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhao, H.; Kosma, D.K.; Tomasi, P.; Dyer, J.M.; Li, R.; Liu, X.; Wang, Z.; Parsons, E.P.; Jenks, M.A.; et al. The Acyl Desaturase CER17 Is Involved in Producing Wax Unsaturated Primary Alcohols and Cutin Monomers. Plant Physiol. 2017, 173, 1109–1124. [Google Scholar] [CrossRef]
- Bird, D.; Beisson, F.; Brigham, A.; Shin, J.; Greer, S.; Jetter, R.; Kunst, L.; Wu, X.; Yephremov, A.; Samuels, L. Characterization of Arabidopsis ABCG11/WBC11, an ATP Binding Cassette (ABC) Transporter That Is Required for Cuticular Lipid Secretion†: WBC11 Is Required for Cuticular Lipid Transport. Plant J. 2007, 52, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, A.; Mayfield, J.A.; Miley, N.L.; Chau, S.; Fischer, R.L.; Preuss, D. Alterations in CER6, a Gene Identical to CUT1, Differentially Affect Long-Chain Lipid Content on the Surface of Pollen and Stems. Plant Cell 2000, 12, 2001–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tresch, T.; Heilmann, M.; Christiansen, N.; Looser, R.; Grossmann, K. Inhibition of Saturated Very-Long-Chain Fatty Acid Biosynthesis by Mefluidide and Perfluidone, Selective Inhibitors of 3-Ketoacyl-CoA Synthases. Phytochemistry 2012, 76, 162–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Kunst, L. SUPERKILLER Complex Components Are Required for the RNA Exosome-Mediated Control of Cuticular Wax Biosynthesis in Arabidopsis Inflorescence Stems. Plant Physiol. 2016, 121, 960–973. [Google Scholar] [CrossRef] [Green Version]
- Lü, S.; Zhao, H.; Des Marais, D.L.; Parsons, E.P.; Wen, X.; Xu, X.; Bangarusamy, D.K.; Wang, G.; Rowland, O.; Juenger, T.; et al. Arabidopsis ECERIFERUM9 Involvement in Cuticle Formation and Maintenance of Plant Water Status. Plant Physiol. 2012, 159, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.Y.; Song, t.; Kosma, D.K.; Parsons, E.P.; Rowland, O.; Jenks, M.A. Arabidopsis CER8 Encodes LONG-CHAIN ACYL-COA SYNTHETASE 1 (LACS1) That Has Overlapping Functions with LACS2 in Plant Wax and Cutin Synthesis. Plant Cell 2009, 59, 553–564. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Yu, L.; Taji, T. ECERIFERUM 10 Encoding an Enoyl-CoA Reductase Plays a Crucial Role in Osmotolerance and Cuticular Wax Loading in Arabidopsis. Front. Plant Sci. 2022, 13, 12. [Google Scholar] [CrossRef]
- Shi, L.; Dean, G.H.; Zheng, H.; Meents, M.J.; Haslam, T.M.; Haughn, G.W.; Kunst, L. ECERIFERUM11/C-TERMINAL DOMAIN PHOSPHATASE-LIKE2 Affects Secretory Trafficking. Plant Physiol. 2019, 181, 901–915. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.; Kunst, L.; Jetter, R. Composition of Alkyl Esters in the Cuticular Wax on Inflorescence Stems of Arabidopsis Thaliana Cer Mutants: Esters in Stem Waxes of Arabidopsis Cer Mutants. Plant J. 2007, 50, 189–196. [Google Scholar] [CrossRef]
- Kenrick, P.; Crane, P.R. The Origin and Early Evolution of Plants on Land. Nature 1997, 389, 33–39. [Google Scholar] [CrossRef]
- Waters, E.R. Molecular Adaptation and the Origin of Land Plants. Mol. Phylogenetics Evol. 2003, 29, 456–463. [Google Scholar] [CrossRef]
- Pennisi, E. The Blue Revolution, Drop by Drop, Gene by Gene. Science 2008, 320, 5873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. China’s Success in Increasing per Capita Food Production. J. Exp. Bot. 2011, 62, 3707–3711. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, T.; Wynne Griffiths, D. The Effects of Stress on Plant Cuticular Waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Kim, H.U.; Suh, M.C. MYB94 and MYB96 Additively Activate Cuticular Wax Biosynthesis in Arabidopsis. Plant Cell Physiol. 2016, 57, 2300–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, P.J.; Lee, S.B.; Suh, M.C.; Park, M.-J.; Go, Y.S.; Park, C.-M. The MYB96 Transcription Factor Regulates Cuticular Wax Biosynthesis under Drought Conditions in Arabidopsis. Plant Cell 2011, 23, 1138–1152. [Google Scholar] [CrossRef] [Green Version]
- Patwari, P.; Salewski, V.; Gutbrod, K.; Kreszies, T.; Dresen-Scholz, B.; Peisker, H.; Steiner, U.; Meyer, A.J.; Schreiber, L.; Dörmann, P. Surface Wax Esters Contribute to Drought Tolerance in Arabidopsis. Plant J. 2019, 98, 727–744. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE Clade of AP2 Domain Transcription Factors Activates Wax Biosynthesis, Alters Cuticle Properties, and Confers Drought Tolerance When Overexpressed in Arabidopsis[W]. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.; Kovalchuk, N.; Langridge, P.; Tricker, P.J.; Lopato, S.; Borisjuk, N. The Impact of Drought on Wheat Leaf Cuticle Properties. BMC Plant Biol. 2017, 17, 85. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Liu, L.; Chen, Y.; Liu, T.; Jiang, Q.; Wei, Z.; Li, C.; Wang, Z. Tomato SlCER1–1 Catalyzes the Synthesis of Wax Alkanes, Increasing Drought Tolerance and Fruit Storability. Hortic. Res. 2022, 9, uhac004. [Google Scholar] [CrossRef]
- Dimopoulos, N.; Tindjau, R.; Wong, D.C.J.; Matzat, T.; Haslam, T.; Song, C.; Gambetta, G.A.; Kunst, L.; Castellarin, S.D. Drought Stress Modulates Cuticular Wax Composition of the Grape Berry. J. Exp. Bot. 2020, 71, 3126–3141. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Shi, S.; Lu, X.; Li, T.; Wang, J.; Liu, T.; Zhang, Q.; Sun, W.; Li, C.; Wang, Z.; et al. Expression Analysis and Functional Characterization of CER1 Family Genes Involved in Very-Long-Chain Alkanes Biosynthesis in Brachypodium Distachyon. Front. Plant Sci. 2019, 10, 1389. [Google Scholar] [CrossRef] [PubMed]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.; Wang, M.; Bi, Q.; Cui, Y.; Wang, L. Transcriptome and Physiological Analyses Provide Insights into the Leaf Epicuticular Wax Accumulation Mechanism in Yellowhorn. Hortic. Res. 2021, 8, 134. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortés, A.J.; This, D. Identification of an ERECTA Gene and Its Drought Adaptation Associations with Wild and Cultivated Common Bean. Plant Sci. 2016, 242, 250–259. [Google Scholar] [CrossRef]
- Cortés, A.J.; This, D.; Chavarro, C.; Madriñán, S.; Blair, M.W. Nucleotide Diversity Patterns at the Drought-Related DREB2 Encoding Genes in Wild and Cultivated Common Bean (Phaseolus Vulgaris L.). Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef]
- Fischer, I.; Camus-Kulandaivelu, L.; Allal, F.; Stephan, W. Adaptation to Drought in Two Wild Tomato Species: The Evolution of the Asr Gene Family. New Phytol. 2011, 190, 1032–1044. [Google Scholar] [CrossRef]
- Ai, W.; Liu, Y.; Mei, M.; Zhang, X.; Tan, E.; Liu, H.; Han, X.; Zhan, H.; Lu, X. A Chromosome-scale Genome Assembly of the Mongolian Oak (Quercus Mongolica). Mol. Ecol. Resour. 2022, 22, 2396–2410. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Number of Amino Acids | Molecular Weight/kD | Theoretical pI | Instability Index (II) | Aliphatic Index | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| CmCER1-1 | Cm03G01983 | 588 | 68.88 | 8.97 | 33.52 | 94.18 | Vacuole |
| CmCER1-2 | Cm03G01987 | 623 | 72.82 | 8.58 | 34.99 | 96.42 | Vacuole |
| CmCER1-3 | Cm03G01992 | 623 | 72.77 | 8.58 | 34.42 | 96.26 | Vacuole |
| CmCER1-4 | Cm10G01890 | 623 | 71.85 | 8.72 | 34.72 | 99.49 | Vacuole |
| CmCER1-5 | Cm10G02076 | 474 | 54.37 | 9.02 | 35.86 | 96.22 | Cell membrane Vacuole |
| CmCER1-6 | Cm12G02161 | 632 | 73.08 | 8.03 | 31.51 | 97.74 | Vacuole |
| CmCER1-7 | Cm10G01980 | 557 | 63.69 | 8.85 | 33.95 | 98.13 | Vacuole |
| CmCER1-8 | Cm03G01976 | 619 | 72.19 | 9.18 | 34.23 | 93.51 | Vacuole |
| CmCER1-9 | Cm03G01984 | 142 | 16.50 | 6.68 | 42.9 | 97.61 | Vacuole |
| CmCER2 | Cm08G01333 | 446 | 49.80 | 5.28 | 32.03 | 88.25 | Cytoplasm |
| CmCER3 | Cm01G00496 | 628 | 71.94 | 8.47 | 31.96 | 102.72 | Vacuole |
| CmCER4 | Cm07G00737 | 490 | 55.19 | 8.36 | 25.77 | 104.22 | Golgi apparatus |
| CmCER5-1 | Cm07G00263 | 696 | 77.79 | 9.35 | 48.11 | 93.99 | Cell membrane |
| CmCER5-2 | Cm07G00264 | 682 | 75.88 | 8.67 | 43.98 | 99.5 | Cell membrane |
| CmCER6-1 | Cm07G01950 | 496 | 55.85 | 9.01 | 40.14 | 99.11 | Cytoplasm |
| CmCER6-2 | Cm10G01697 | 507 | 57.07 | 9.29 | 37.49 | 94.97 | Cytoplasm |
| CmCER7 | Cm07G00259 | 457 | 49.88 | 5.94 | 53.18 | 76.21 | Chloroplast Mitochondrion |
| CmCER8 | Cm12G00788 | 654 | 73.85 | 6.36 | 29.42 | 86.73 | Peroxisome |
| CmCER9-1 | Cm10G01351 | 1109 | 123.64 | 5.85 | 36.29 | 109.1 | Cell membrane Nucleus |
| CmCER9-2 | Cm11G02224 | 534 | 58.97 | 6.11 | 35.25 | 106.84 | Cell membrane |
| CmCER10-1 | Cm05G00914 | 308 | 36.04 | 9.61 | 41.52 | 86.66 | Nucleus |
| CmCER10-2 | Cm11G02385 | 310 | 36.31 | 9.61 | 48.66 | 90.23 | Cell membrane Golgi apparatus |
| CmCER11-1 | Cm11G01507 | 854 | 95.40 | 6.35 | 51.84 | 80.68 | Nucleus |
| CmCER11-2 | Cm02G02052 | 952 | 106.28 | 5.93 | 56.97 | 80.09 | Chloroplast Nucleus |
| CmCER13 | Cm01G01426 | 1852 | 206.05 | 5.84 | 46.55 | 111.02 | Nucleus |
| CmCER17 | Cm02G02326 | 389 | 44.47 | 9.64 | 46.71 | 86.97 | Chloroplast Endoplasmic reticulum |
| CmCER60-1 | Cm01G03190 | 510 | 57.62 | 9.08 | 42.32 | 94.63 | Chloroplast Cytoplasm |
| CmCER60-2 | Cm06G01231 | 457 | 50.95 | 8.74 | 37.59 | 97.72 | Cytoplasm |
| CmCER60-3 | Cm09G01848 | 463 | 52.35 | 8.69 | 43.47 | 93.89 | Cytoplasm |
| CmCER60-4 | Cm09G01872 | 463 | 52.35 | 8.69 | 43.47 | 93.89 | Cytoplasm |
| CmCER60-5 | Cm08G02023 | 478 | 53.64 | 9.71 | 34.43 | 99.96 | Cytoplasm |
| CmCER60-6 | Cm09G01873 | 370 | 42.02 | 9.58 | 40.14 | 98.54 | Chloroplast Cytoplasm |
| CmCER60-7 | Cm05G00220 | 532 | 60.17 | 8.99 | 32.68 | 90.55 | Cytoplasm |
| CmCER60-8 | Cm08G02594 | 523 | 59.02 | 9.27 | 39.35 | 92.64 | Cytoplasm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Nie, X.; Liu, X.; Wang, B.; Liu, S.; Qin, L.; Xing, Y. Genome-Wide Identification of the CER Gene Family and Significant Features in Climate Adaptation of Castanea mollissima. Int. J. Mol. Sci. 2022, 23, 16202. https://doi.org/10.3390/ijms232416202
Zhao S, Nie X, Liu X, Wang B, Liu S, Qin L, Xing Y. Genome-Wide Identification of the CER Gene Family and Significant Features in Climate Adaptation of Castanea mollissima. International Journal of Molecular Sciences. 2022; 23(24):16202. https://doi.org/10.3390/ijms232416202
Chicago/Turabian StyleZhao, Shuqing, Xinghua Nie, Xueqing Liu, Biyao Wang, Song Liu, Ling Qin, and Yu Xing. 2022. "Genome-Wide Identification of the CER Gene Family and Significant Features in Climate Adaptation of Castanea mollissima" International Journal of Molecular Sciences 23, no. 24: 16202. https://doi.org/10.3390/ijms232416202