
Huperzine—A Improved Animal Behavior in Cuprizone-Induced Mouse Model by Alleviating Demyelination and Neuroinflammation
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
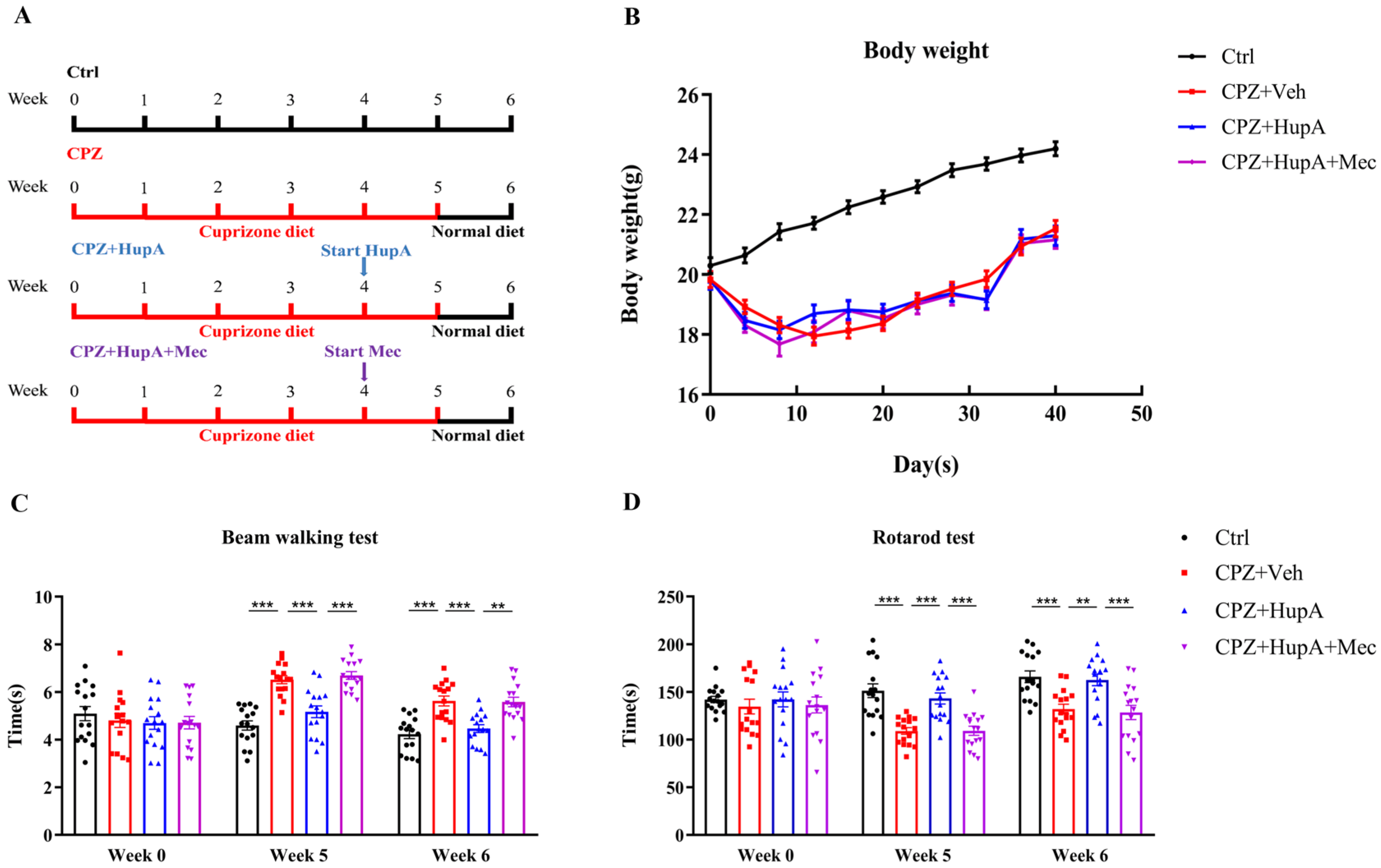
2.1. HupA Enhanced the Motor Coordination Function of Demyelinated Mice
2.2. HupA Diminished Anxiety and Ameliorates Spatial Memory Deficit in Demyelinated Mice
2.3. HupA Promoted the Expression of Myelin Basic Protein (MBP) in the Corpus Callosum and Increased the Quantity of the Myelinated Axons
2.4. HupA Did Not have an Impact on the Quantity of Oligodendrocyte Precursor Cells (OPCs) but Increased the Quantity of the Mature Oligodendrocytes In Vivo Settings
2.5. HupA Did Not Affect the Proliferation of OPCs but Promoted the Differentiation of OPCs In Vitro Settings
2.6. HupA Can Regulate the Activation of Microglia in the Corpus Callosum of Demyelinated Mice
2.7. HupA Down-Regulated and Up-Regulated the mRNA Expression of Pro-Inflammatory and Anti-Inflammatory Genes in the Microglia Respectively
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. HupA Treatment
4.3. Animal Behavior
4.4. BWT and Rotarod Test
4.5. Y Maze and OFT
4.6. Tissue Preparation
4.7. Western Blot
4.8. OPC Isolation and Culture
4.9. OPC Differentiation and CTG
4.9.1. ICC
4.9.2. IHC
4.9.3. LFB Staining
4.10. TEM
4.11. Microglia Isolation and Quantitative RT-PCR (qRT-PCR)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Cotsapas, C.; Mitrovic, M.; Hafler, D. Multiple sclerosis. Handb. Clin. Neurol. 2018, 148, 723–730. [Google Scholar] [PubMed]
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple Sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef] [PubMed]
- Sen, M.K.; Mahns, D.A.; Coorssen, J.R.; Shortland, P.J. Behavioural phenotypes in the cuprizone model of central nervous system demyelination. Neurosci. Biobehav. Rev. 2019, 107, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Lucchinetti, C.F.; Brück, W.; Rodriguez, M.; Lassmann, H. Distinct patterns of multiple sclerosis pathology indicates heterogeneity on pathogenesis. Brain Pathol. 1996, 6, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Lucchinetti, C.; Brück, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of multiple sclerosis lesions: Implications for the pathogenesis of demyelination. Ann. Neurol. 2000, 47, 707–717. [Google Scholar] [CrossRef]
- Lemus, H.N.; Warrington, A.E.; Rodriguez, M. Multiple Sclerosis: Mechanisms of Disease and Strategies for Myelin and Axonal Repair. Neurol. Clin. 2018, 36, 1–11. [Google Scholar] [CrossRef]
- Yin, J.J.; He, Y.; An, J.; Miao, Q.; Sui, R.X.; Wang, Q.; Yu, J.Z.; Xiao, B.G.; Ma, C.G. Dynamic Balance of Microglia and Astrocytes Involved in the Remyelinating Effect of Ginkgolide B. Front. Cell Neurosci. 2019, 13, 572. [Google Scholar] [CrossRef]
- Vaknine, S.; Soreq, H. Central and peripheral anti-inflammatory effects of acetylcholinesterase inhibitors. Neuropharmacology 2020, 168, 108020. [Google Scholar] [CrossRef]
- Tsai, S.J. Huperzine-A, a versatile herb, for the treatment of Alzheimer’s disease. J. Chin. Med. Assoc. 2019, 82, 750–751. [Google Scholar] [CrossRef]
- Yang, G.; Wang, Y.; Tian, J.; Liu, J.P. Huperzine A for Alzheimer’s disease: A systematic review and meta-analysis of randomized clinical trials. PLoS ONE 2013, 8, e74916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.R.; Tay, K.C.; Su, Y.X.; Wong, C.K.; Tan, W.N.; Khaw, K.Y. Potential of Naturally Derived Alkaloids as Multi-Targeted Therapeutic Agents for Neurodegenerative Diseases. Molecules 2021, 26, 728. [Google Scholar] [CrossRef] [PubMed]
- Nett, R.S.; Dho, Y.; Low, Y.Y.; Sattely, E.S. A metabolic regulon reveals early and late acting enzymes in neuroactive Lycopodium alkaloid biosynthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2102949118. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Y.; Chen, C.P.; Jinn, T.R. Traditional Chinese medicines and Alzheimer’s disease. Taiwan J. Obstet. Gynecol. 2011, 50, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, R.A.; Sarpong, R. Heathcock-inspired strategies for the synthesis of fawcettimine-type Lycopodium alkaloids. Chemistry 2014, 20, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.R.; Fang, R.; Houghton, P.J. Effect of Chinese Herbal Medicine on Alzheimer’s Disease. Int. Rev. Neurobiol. 2017, 135, 29–56. [Google Scholar] [PubMed]
- Coleman, B.R.; Ratcliffe, R.H.; Oguntayo, S.A.; Shi, X.; Doctor, B.P.; Gordon, R.K.; Nambiar, M.P. [+]-Huperzine A treatment protects against N-methyl-D-aspartate-induced seizure/status epilepticus in rats. Chem. Biol. Interact. 2008, 175, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Ghassab-Abdollahi, N.; Mobasseri, K.; Dehghani Ahmadabad, A.; Nadrian, H.; Mirghafourvand, M. The effects of Huperzine A on dementia and mild cognitive impairment: An overview of systematic reviews. Phytother. Res. 2021, 35, 4971–4987. [Google Scholar] [CrossRef]
- Bialer, M.; Johannessen, S.I.; Levy, R.H.; Perucca, E.; Tomson, T.; White, H.S. Progress report on new antiepileptic drugs: A summary of the Twelfth Eilat Conference (EILAT XII). Epilepsy Res. 2015, 111, 85–141. [Google Scholar] [CrossRef]
- Gersner, R.; Ekstein, D.; Dhamne, S.C.; Schachter, S.C.; Rotenberg, A. Huperzine A prophylaxis against pentylenetetrazole-induced seizures in rats is associated with increased cortical inhibition. Epilepsy Res. 2015, 117, 97–103. [Google Scholar] [CrossRef]
- Ruan, Q.; Liu, F.; Gao, Z.; Kong, D.; Hu, X.; Shi, D.; Bao, Z.; Yu, Z. The anti-inflamm-aging and hepatoprotective effects of huperzine A in D-galactose-treated rats. Mech. Ageing Dev. 2013, 134, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Desilets, A.R.; Gickas, J.J.; Dunican, K.C. Role of huperzine a in the treatment of Alzheimer’s disease. Ann. Pharmacother. 2009, 43, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, F.; Zheng, P.; Deng, W.; Yuan, J.; Peng, B.; Wang, R.; Liu, W.; Zhao, H.; Wang, Y.; et al. Huperzine A ameliorates experimental autoimmune encephalomyelitis via the suppression of T cell-mediated neuronal inflammation in mice. Exp. Neurol. 2012, 236, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, X.; Chen, Q.; Shu, L.; Wang, J.; Shan, G. Clinical efficacy and safety of huperzine Alpha in treatment of mild to moderate Alzheimer disease, a placebo-controlled, double-blind, randomized trial. Zhonghua Yi Xue Za Zhi 2002, 82, 941–944. [Google Scholar] [PubMed]
- Gul, A.; Bakht, J.; Mehmood, F. Huperzine-A response to cognitive impairment and task switching deficits in patients with Alzheimer’s disease. J. Chin. Med. Assoc. 2019, 82, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Koutsoudaki, P.N.; Skripuletz, T.; Gudi, V.; Moharregh-Khiabani, D.; Hildebrandt, H.; Trebst, C.; Stangel, M. Demyelination of the hippocampus is prominent in the cuprizone model. Neurosci. Lett 2009, 451, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Skripuletz, T.; Bussmann, J.H.; Gudi, V.; Koutsoudaki, P.N.; Pul, R.; Moharregh-Khiabani, D.; Lindner, M.; Stangel, M. Cerebellar cortical demyelination in the murine cuprizone model. Brain Pathol. 2010, 20, 301–312. [Google Scholar] [CrossRef]
- Blakemore, W.F. Demyelination of the superior cerebellar peduncle in the mouse induced by cuprizone. J. Neurol. Sci. 1973, 20, 63–72. [Google Scholar] [CrossRef]
- Gudi, V.; Moharregh-Khiabani, D.; Skripuletz, T.; Koutsoudaki, P.N.; Kotsiari, A.; Skuljec, J.; Trebst, C.; Stangel, M. Regional differences between grey and white matter in cuprizone induced demyelination. Brain Res. 2009, 1283, 127–138. [Google Scholar] [CrossRef]
- Matsushima, G.K.; Morell, P. The neurotoxicant, cuprizone, as a model to study demyelination and remyelination in the central nervous system. Brain Pathol. 2001, 11, 107–116. [Google Scholar] [CrossRef]
- Zhu, K.; Sun, J.; Kang, Z.; Zou, Z.; Wu, G.; Wang, J. Electroacupuncture Promotes Remyelination after Cuprizone Treatment by Enhancing Myelin Debris Clearance. Front. Neurosci. 2016, 10, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, W.; Liu, A.; Lu, J.; Zhang, W.; Tattersall, D.; Wang, J. An Alternative Cuprizone-Induced Demyelination and Remyelination Mouse Model. ASN Neuro 2017, 9, 1759091417725174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, C.B.; Sun, S.; Roholt, A.; Benson, E.; Edberg, D.; Medicetty, S.; Dutta, R.; Kidd, G.; Macklin, W.B.; Trapp, B. A mouse model for testing remyelinating therapies. Exp. Neurol. 2016, 283 Pt A, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Morell, P.; Barrett, C.V.; Mason, J.L.; Toews, A.D.; Hostettler, J.D.; Knapp, G.W.; Matsushima, G.K. Gene expression in brain during cuprizone-induced demyelination and remyelination. Mol. Cell Neurosci. 1998, 12, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Vega-Riquer, J.M.; Mendez-Victoriano, G.; Morales-Luckie, R.A.; Gonzalez-Perez, O. Five Decades of Cuprizone, an Updated Model to Replicate Demyelinating Diseases. Curr. Neuropharmacol. 2019, 17, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Zirngibl, M.; Assinck, P.; Sizov, A.; Caprariello, A.V.; Plemel, J.R. Oligodendrocyte death and myelin loss in the cuprizone model: An updated overview of the intrinsic and extrinsic causes of cuprizone demyelination. Mol. Neurodegener. 2022, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bi, X.; Adebiyi, O.; Wang, J.; Mooshekhian, A.; Cohen, J.; Wei, Z.; Wang, F.; Li, X.M. Venlafaxine Improves the Cognitive Impairment and Depression-Like Behaviors in a Cuprizone Mouse Model by Alleviating Demyelination and Neuroinflammation in the Brain. Front. Pharmacol. 2019, 10, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.Y.; Gu, L.H.; Ma, D.L.; Wang, M.Y.; Yang, C.C.; Zhang, L.; Li, X.M.; Zhang, J.W.; Li, L. Behavioral and neurobiological changes in a novel mouse model of schizophrenia induced by the combination of cuprizone and MK-801. Brain Res. Bull. 2021, 174, 141–152. [Google Scholar] [CrossRef]
- Liu, Q.; Lv, H.W.; Yang, S.; He, Y.Q.; Ma, Q.R.; Liu, J. NEP1-40 alleviates behavioral phenotypes and promote oligodendrocyte progenitor cell differentiation in the hippocampus of cuprizone-induced demyelination mouse model. Neurosci. Lett. 2020, 725, 134872. [Google Scholar] [CrossRef]
- Xu, Z.; Adilijiang, A.; Wang, W.; You, P.; Lin, D.; Li, X.; He, J. Arecoline attenuates memory impairment and demyelination in a cuprizone-induced mouse model of schizophrenia. Neuroreport 2019, 30, 134–138. [Google Scholar] [CrossRef]
- Martinsen, V.; Kursula, P. Multiple sclerosis and myelin basic protein: Insights into protein disorder and disease. Amino Acids 2022, 54, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Zeinali, H.; Baluchnejadmojarad, T.; Roghani, M. Diosgenin ameliorates cellular and molecular changes in multiple sclerosis in C57BL/6 mice. Mult. Scler. Relat. Disord. 2021, 55, 103211. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Shen, M.; Kitamura, N.; Sennari, Y.; Morita, K.; Tsukada, J.; Kanazawa, T.; Yoshida, Y. Mitogen-activated protein kinases are involved in cucurbitacin D-induced antitumor effects on adult T-cell leukemia cells. Investig. New Drugs 2021, 39, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Lin, J.; Wang, K.; Wei, M.; Chen, Q.; Wang, Y. Huperzine A protects neural stem cells against Aβ-induced apoptosis in a neural stem cells and microglia co-culture system. Int. J. Clin. Exp. Pathol. 2015, 8, 6425–6433. [Google Scholar] [PubMed]
- Young, K.; Morrison, H. Quantifying Microglia Morphology from Photomicrographs of Immunohistochemistry Prepared Tissue Using ImageJ. J. Vis. Exp. 2018, 136, e57648. [Google Scholar] [CrossRef] [Green Version]
- Monje, M. Myelin Plasticity and Nervous System Function. Annu. Rev. Neurosci. 2018, 41, 61–76. [Google Scholar] [CrossRef]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the Central Nervous System: Structure, Function, and Pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef]
- Bolino, A. Myelin Biology. Neurotherapeutics 2021, 18, 2169–2184. [Google Scholar] [CrossRef]
- Faissner, S.; Plemel, J.R.; Gold, R.; Yong, V.W. Progressive multiple sclerosis: From pathophysiology to therapeutic strategies. Nat. Rev. Drug Discov. 2019, 18, 905–922. [Google Scholar] [CrossRef]
- Klistorner, S.; Barnett, M.H.; Wasserthal, J.; Yiannikas, C.; Barton, J.; Parratt, J.; You, Y.; Graham, S.L.; Klistorner, A. Differentiating axonal loss and demyelination in chronic MS lesions: A novel approach using single streamline diffusivity analysis. PLoS ONE 2021, 16, e0244766. [Google Scholar] [CrossRef]
- Rodríguez Murúa, S.; Farez, M.F.; Quintana, F.J. The Immune Response in Multiple Sclerosis. Annu. Rev. Pathol. 2022, 17, 121–139. [Google Scholar] [CrossRef] [PubMed]
- Friedli, M.J.; Inestrosa, N.C. Huperzine A and Its Neuroprotective Molecular Signaling in Alzheimer’s Disease. Molecules 2021, 26, 6531. [Google Scholar] [CrossRef] [PubMed]
- Callizot, N.; Campanari, M.L.; Rouvière, L.; Jacquemot, G.; Henriques, A.; Garayev, E.; Poindron, P. Huperzia serrata Extract ‘NSP01’ With Neuroprotective Effects-Potential Synergies of Huperzine A and Polyphenols. Front. Pharmacol. 2021, 12, 681532. [Google Scholar] [CrossRef] [PubMed]
- Giazkoulidou, A.; Messinis, L.; Nasios, G. Cognitive functions and social cognition in multiple sclerosis: An overview. Hell. J. Nucl. Med. 2019, 22, 102–110. [Google Scholar] [PubMed]
- Oset, M.; Stasiolek, M.; Matysiak, M. Cognitive Dysfunction in the Early Stages of Multiple Sclerosis-How Much and How Important? Curr. Neurol. Neurosci. Rep. 2020, 20, 22. [Google Scholar] [CrossRef] [PubMed]
- De Meo, E.; Portaccio, E.; Giorgio, A.; Ruano, L.; Goretti, B.; Niccolai, C.; Patti, F.; Chisari, C.G.; Gallo, P.; Grossi, P.; et al. Identifying the Distinct Cognitive Phenotypes in Multiple Sclerosis. JAMA Neurol. 2021, 78, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Glukhovsky, L.; Kurz, D.; Brandstadter, R.; Leavitt, V.M.; Krieger, S.; Fabian, M.; Katz Sand, I.; Klineova, S.; Riley, C.S.; Lublin, F.D.; et al. Depression and cognitive function in early multiple sclerosis: Multitasking is more sensitive than traditional assessments. Mult. Scler. 2021, 27, 1276–1283. [Google Scholar] [CrossRef]
- Quik, M.; Wonnacott, S. α6β2* and α4β2* nicotinic acetylcholine receptors as drug targets for Parkinson’s disease. Pharmacol. Rev. 2011, 63, 938–966. [Google Scholar] [CrossRef] [Green Version]
- Hoskin, J.L.; Al-Hasan, Y.; Sabbagh, M.N. Nicotinic Acetylcholine Receptor Agonists for the Treatment of Alzheimer’s Dementia: An Update. Nicotine Tob. Res. 2019, 21, 370–376. [Google Scholar] [CrossRef]
- Koneczny, I.; Herbst, R. Myasthenia Gravis: Pathogenic Effects of Autoantibodies on Neuromuscular Architecture. Cells 2019, 8, 671. [Google Scholar] [CrossRef]
- Liu, Q.; Li, M.; Whiteaker, P.; Shi, F.D.; Morley, B.J.; Lukas, R.J. Attenuation in Nicotinic Acetylcholine Receptor α9 and α10 Subunit Double Knock-Out Mice of Experimental Autoimmune Encephalomyelitis. Biomolecules 2019, 9, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godin, J.R.; Roy, P.; Quadri, M.; Bagdas, D.; Toma, W.; Narendrula-Kotha, R.; Kishta, O.A.; Damaj, M.I.; Horenstein, N.A.; Papke, R.L.; et al. A silent agonist of α7 nicotinic acetylcholine receptors modulates inflammation ex vivo and attenuates EAE. Brain Behav. Immun. 2020, 87, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Ohba, T.; Yoshino, Y.; Ishisaka, M.; Abe, N.; Tsuruma, K.; Shimazawa, M.; Oyama, M.; Tabira, T.; Hara, H. Japanese Huperzia serrata extract and the constituent, huperzine A, ameliorate the scopolamine-induced cognitive impairment in mice. Biosci. Biotechnol. Biochem. 2015, 79, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.X.; Zhu, X.Q.; Chen, Y.; Wu, G.C.; Wang, J. Huperzine A inhibits CCL2 production in experimental autoimmune encephalomyelitis mice and in cultured astrocyte. Int. J. Immunopathol. Pharmacol. 2013, 26, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Kipp, M.; Nyamoya, S.; Hochstrasser, T.; Amor, S. Multiple sclerosis animal models: A clinical and histopathological perspective. Brain Pathol. 2017, 27, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Mann, T.; Joost, S.; Behrangi, N.; Frank, M.; Kipp, M. The Cuprizone Model: Dos and Donts. Cells 2020, 9, 843. [Google Scholar] [CrossRef] [Green Version]
- Toomey, L.M.; Papini, M.; Lins, B.; Wright, A.J.; Warnock, A.; McGonigle, T.; Hellewell, S.C.; Bartlett, C.A.; Anyaegbu, C.; Fitzgerald, M. Cuprizone feed formulation influences the extent of demyelinating disease pathology. Sci. Rep. 2021, 11, 22594. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.R.; Kang, Y.H.; Jeon, H.; Lee, S.; Park, S.J.; Song, D.Y.; Min, S.S.; Yoo, S.M.; Lee, M.S.; Lee, S.H. Differential Expression of miRNAs and Behavioral Change in the Cuprizone-Induced Demyelination Mouse Model. Int. J. Mol. Sci. 2020, 21, 646. [Google Scholar] [CrossRef] [Green Version]
- Snyder, C.N.; Brown, A.R.; Buffalari, D. Similar tests of anxiety-like behavior yield different results: Comparison of the open field and free exploratory rodent procedures. Physiol. Behav. 2021, 230, 113246. [Google Scholar] [CrossRef]
- Du, Y.; Liang, H.; Zhang, L.; Fu, F. Administration of Huperzine A exerts antidepressant-like activity in a rat model of post-stroke depression. Pharmacol. Biochem. Behav. 2017, 158, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yao, Y.; Hu, Y.; Yang, J.; Zhang, C.; He, Y.; Zhang, A.; Liu, X.; Zhang, C.; Gan, G. Valproic acid suppresses cuprizone-induced hippocampal demyelination and anxiety-like behavior by promoting cholesterol biosynthesis. Neurobiol. Dis. 2021, 158, 105489. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Guo, Y.E.; Fang, J.H.; Shi, C.J.; Suo, N.; Zhang, R.; Xie, X. Donepezil, a drug for Alzheimer’s disease, promotes oligodendrocyte generation and remyelination. Acta Pharmacol. Sin. 2019, 40, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Molina-Holgado, E.; Khorchid, A.; Liu, H.N.; Almazan, G. Regulation of muscarinic receptor function in developing oligodendrocytes by agonist exposure. Br. J. Pharmacol. 2003, 138, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, S.W.; Gregori, N.Z.; Carlson, N.; Gahring, L.C.; Noble, M. Neuronal nicotinic acetylcholine receptor expression by O2A/oligodendrocyte progenitor cells. Glia 2001, 33, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Masuda, T.; Wheeler, M.A.; Quintana, F.J. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu. Rev. Immunol. 2021, 39, 251–277. [Google Scholar] [CrossRef] [PubMed]
- Spiteri, A.G.; Wishart, C.L.; Pamphlett, R.; Locatelli, G.; King, N.J.C. Microglia and monocytes in inflammatory CNS disease: Integrating phenotype and function. Acta Neuropathol. 2022, 143, 179–224. [Google Scholar] [CrossRef]
- Lamberty, Y.; Gower, A.J.; Gobert, J.; Hanin, I.; Wulfert, E. Behavioural, biochemical and histological effects of AF64A following injection into the third ventricle of the mouse. Behav. Brain Res. 1992, 51, 165–177. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, H.; Jiang, W.; Xiao, L.; Yan, B.; He, J.; Wang, Y.; Bi, X.; Li, X.; Kong, J.; et al. Quetiapine alleviates the cuprizone-induced white matter pathology in the brain of C57BL/6 mouse. Schizophr. Res. 2008, 106, 182–191. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, D.; Sun, J.; Wang, Y.; Wu, S.; Wang, J. Huperzine—A Improved Animal Behavior in Cuprizone-Induced Mouse Model by Alleviating Demyelination and Neuroinflammation. Int. J. Mol. Sci. 2022, 23, 16182. https://doi.org/10.3390/ijms232416182
Zhang H, Wang D, Sun J, Wang Y, Wu S, Wang J. Huperzine—A Improved Animal Behavior in Cuprizone-Induced Mouse Model by Alleviating Demyelination and Neuroinflammation. International Journal of Molecular Sciences. 2022; 23(24):16182. https://doi.org/10.3390/ijms232416182
Chicago/Turabian StyleZhang, Hongyu, Danjie Wang, Jingxian Sun, Yumeng Wang, Shuai Wu, and Jun Wang. 2022. "Huperzine—A Improved Animal Behavior in Cuprizone-Induced Mouse Model by Alleviating Demyelination and Neuroinflammation" International Journal of Molecular Sciences 23, no. 24: 16182. https://doi.org/10.3390/ijms232416182