
Quercetin Ameliorates Testicular Damage in Zucker Diabetic Fatty Rats through Its Antioxidant, Anti-Inflammatory and Anti-Apoptotic Properties
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
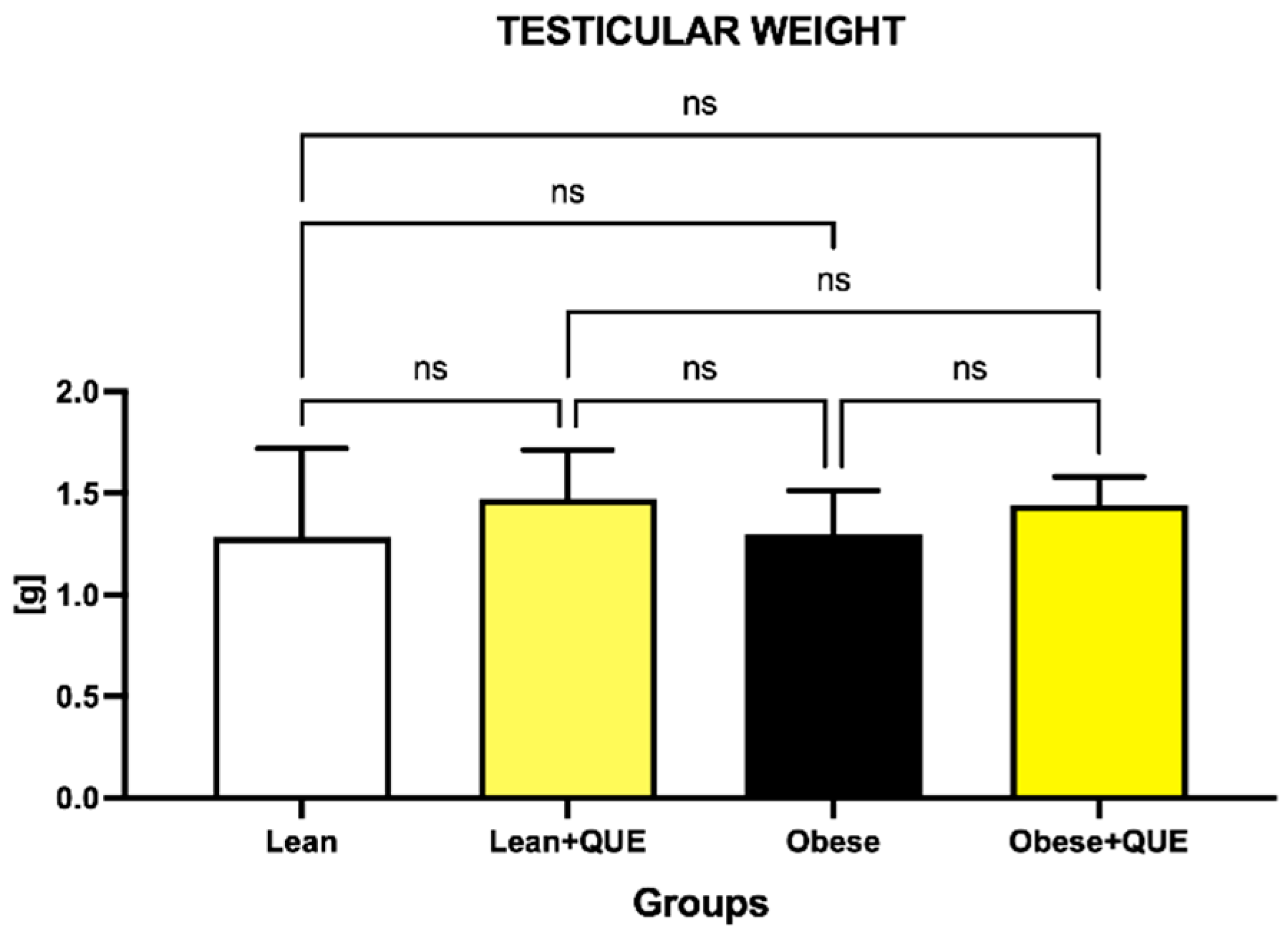
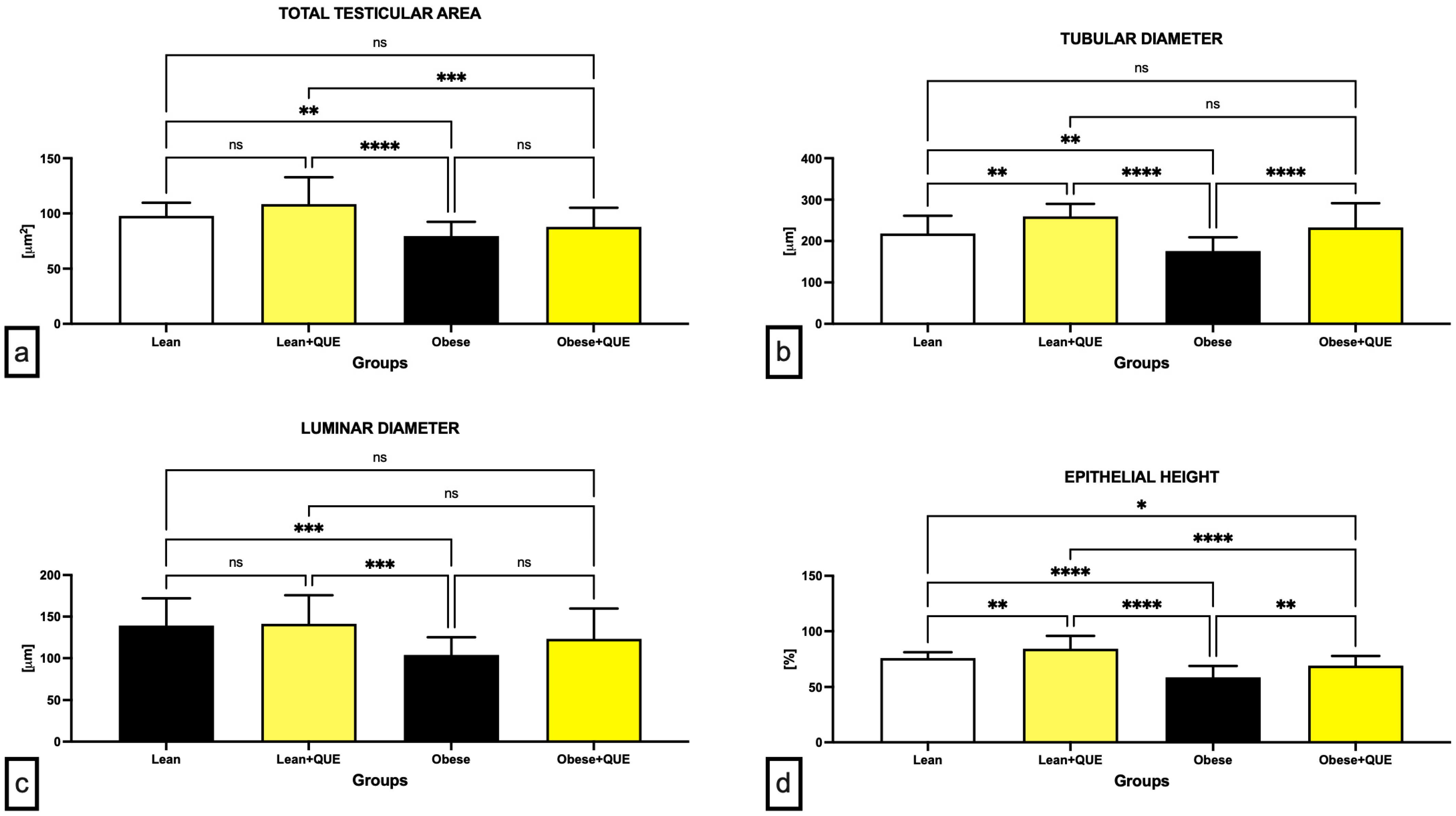
2.1. Histology and Morphometry
2.2. Oxidative Profile
2.3. Cytokines
2.4. Western Blot
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Histology and Morphometry
4.3. Tissue Lysis
4.4. Oxidative Profile Assessment
4.5. Cytokine Analysis
4.6. Western Blotting
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sakran, N.; Graham, Y.; Pintar, T.; Yang, W.; Kassir, R.; Willigendael, E.M.; Singhal, R.; Kooreman, Z.E.; Ramnarain, D.; Mahawar, K.; et al. The many faces of diabetes. Is there a need for re-classification? A narrative review. BMC Endocr. Disord. 2022, 22, 9. [Google Scholar] [CrossRef] [PubMed]
- Nisar, O.; Pervez, H.; Mandalia, B.; Waqas, M.; Sra, H.K. Type 3 Diabetes Mellitus: A Link Between Alzheimer’s Disease and Type 2 Diabetes Mellitus. Cureus 2020, 12, e11703. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.; Bain, S.; Kanamarlapudi, V. A Review of Current Trends with Type 2 Diabetes Epidemiology, Aetiology, Pathogenesis, Treatments and Future Perspectives. Diabetes, Metab. Syndr. Obes. Targets Ther. 2021, ume 14, 3567–3602. [Google Scholar] [CrossRef]
- Safiri, S.; Karamzad, N.; Kaufman, J.S.; Bell, A.W.; Nejadghaderi, S.A.; Sullman, M.J.M.; Moradi-Lakeh, M.; Collins, G.; Kolahi, A.-A. Prevalence, Deaths and Disability-Adjusted-Life-Years (DALYs) Due to Type 2 Diabetes and Its Attributable Risk Factors in 204 Countries and Territories, 1990-2019: Results from the Global Burden of Disease Study 2019. Front. Endocrinol. 2022, 13, 838027. [Google Scholar] [CrossRef] [PubMed]
- Chobot, A.; Górowska-Kowolik, K.; Sokołowska, M.; Jarosz-Chobot, P. Obesity and diabetes-Not only a simple link between two epidemics. Diabetes/Metabolism Res. Rev. 2018, 34, e3042. [Google Scholar] [CrossRef] [Green Version]
- Condorelli, R.A.; La Vignera, S.; Mongioì, L.M.; Alamo, A.; Calogero, A.E. Diabetes Mellitus and Infertility: Different Pathophysiological Effects in Type 1 and Type 2 on Sperm Function. Front. Endocrinol. 2018, 9, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leisegang, K.; Sengupta, P.; Agarwal, A.; Henkel, R. Obesity and male infertility: Mechanisms and management. Andrologia 2021, 53, e13617. [Google Scholar] [CrossRef]
- Zhong, O.; Ji, L.; Wang, J.; Lei, X.; Huang, H. Association of diabetes and obesity with sperm parameters and testosterone levels: A meta-analysis. Diabetol. Metab. Syndr. 2021, 13, 109. [Google Scholar] [CrossRef]
- Ye, R.-J.; Yang, J.-M.; Hai, D.-M.; Liu, N.; Ma, L.; Lan, X.-B.; Niu, J.-G.; Zheng, P.; Yu, J.-Q. Interplay between male reproductive system dysfunction and the therapeutic effect of flavonoids. Fitoterapia 2020, 147, 104756. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Zou, H.; Ye, H.; Kamaraj, R.; Zhang, T.; Zhang, J.; Pavek, P. A review on pharmacological activities and synergistic effect of quercetin with small molecule agents. Phytomedicine 2021, 92, 153736. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Tušimová, E.; Kováčik, A.; Paál, D.; Libová, L.; Lukáč, N. Protective Effects of Quercetin on Selected Oxidative Biomarkers in Bovine Spermatozoa Subjected to Ferrous Ascorbate. Reprod. Domest. Anim. 2016, 51, 524–537. [Google Scholar] [CrossRef]
- Cheraghi, E.; Sajadi, S.M.S.; Mehranjani, M.S. The effect of Quercetin on the quality of sperm parameters in frozen-thawed semen of patients with Asthenospermia. Andrologia 2021, 53, e14167. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, S.U. Quercetin and its role in biological functions: An updated review. EXCLI J. 2018, 17, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Coldiron, A.D.; Sanders, R.A.; Watkins, J.B. Effects of combined quercetin and coenzyme Q10 treatment on oxidative stress in normal and diabetic rats. J. Biochem. Mol. Toxicol. 2002, 16, 197–202. [Google Scholar] [CrossRef]
- Arya, A.; Al-Obaidi, M.M.J.; Shahid, N.; Bin Noordin, M.I.; Looi, C.Y.; Wong, W.F.; Khaing, S.L.; Mustafa, M.R. Synergistic effect of quercetin and quinic acid by alleviating structural degeneration in the liver, kidney and pancreas tissues of STZ-induced diabetic rats: A mechanistic study. Food Chem. Toxicol. 2014, 71, 183–196. [Google Scholar] [CrossRef]
- Kanter, M.; Aktas, C.; Erboga, M. Protective effects of quercetin against apoptosis and oxidative stress in streptozotocin-induced diabetic rat testis. Food Chem. Toxicol. 2012, 50, 719–725. [Google Scholar] [CrossRef]
- Ojo, O.O.; Olorunsogo, O.O. Quercetin and vitamin E attenuate diabetes-induced testicular anomaly in Wistar rats via the mitochondrial-mediated apoptotic pathway. Andrologia 2021, 53, e14185. [Google Scholar] [CrossRef]
- Hajiyeva, R.; Durmuş, S.; Tunçdemir, M.; Gelişgen, R.; Sözer, V.; Mirzataş, E.B.; Şimsek, G.; Uzun, H. The Role of Quercetin in Alleviating the Testicular Oxidative Stress and Apoptosis in Streptozotocin-Induced Diabetes. Cerrah-Med J. 2021, 45, 159–166. [Google Scholar] [CrossRef]
- Kottaisamy, C.P.D.; Raj, D.S.; Kumar, V.P.; Sankaran, U. Experimental animal models for diabetes and its related complications—A review. Lab. Anim. Res. 2021, 37, 23. [Google Scholar] [CrossRef]
- Pang, Y.-L.; Hu, J.-W.; Liu, G.-L.; Lu, S.-Y. Comparative medical characteristics of ZDF-T2DM rats during the course of development to late stage disease. Anim. Model. Exp. Med. 2018, 1, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Kováč, J.; Benko, F.; Ďuračka, M.; Varga, A.; Uličná, O.; Almášiová, V.; Capcarová, M.; Chomová, M. Characterization of the structural, oxidative, and immunological features of testis tissue from Zucker diabetic fatty rats. Open Life Sci. 2022, 17, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- Vendramini, V.; Cedenho, A.P.; Miraglia, S.M.; Spaine, D.M. Reproductive Function of the Male Obese Zucker Rats: Alteration in Sperm Production and Sperm DNA Damage. Reprod. Sci. 2013, 21, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, N.; Tsuji, M.; Tamura, M.; Kagawa, S. Transforming growth factor (beta1) in testes of aged and diabetic rats: Correlation with testicular function. Arch. Androl. 2001, 47, 217–226. [Google Scholar] [CrossRef]
- Vessal, M.; Hemmati, M.; Vasei, M. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2003, 135, 357–364. [Google Scholar] [CrossRef]
- Benko, F.; Chomová, M.; Uličná, O.; Ďuračka, M.; Kováč, J.; Tvrdá, E. The impact of diabetes mellitus type 2 on the steroidogenesis of male Zucker Diabetic Fatty rats. Physiol. Res. 2022, 71, 713–717. [Google Scholar]
- Khaki, A.; Fathiazad, F.; Nouri, M.; Khaki, A.; Maleki, N.A.; Khamnei, H.J.; Ahmadi, P. Beneficial effects of quercetin on sperm parameters in streptozotocin-induced diabetic male rats. Phytother. Res. 2010, 24, 1285–1291. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Zhai, Q.-Q.; Zhu, Y.-F.; Liu, B.-Y.; Xu, Y. Quercetin ameliorates testosterone secretion disorder by inhibiting endoplasmic reticulum stress through the miR-1306-5p/HSD17B7 axis in diabetic rats. Bosn. J. Basic Med. Sci. 2022, 22, 191–204. [Google Scholar] [CrossRef]
- Saito, M.; Ueno, M.; Ogino, S.; Kubo, K.; Nagata, J.; Takeuchi, M. High dose of Garcinia cambogia is effective in suppressing fat accumulation in developing male Zucker obese rats, but highly toxic to the testis. Food Chem. Toxicol. 2005, 43, 411–419. [Google Scholar] [CrossRef]
- Omar, S.S.; Aly, R.G.; Badae, N.M. Vitamin E improves testicular damage in streptozocin-induced diabetic rats, via increasing vascular endothelial growth factor and poly(ADP-ribose) polymerase-1. Andrologia 2018, 50, e12925. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Zrieq, R.; Al-Quraishy, S.; Moneim, A.E.A. Selenium Nanoparticles Attenuate Oxidative Stress and Testicular Damage in Streptozotocin-Induced Diabetic Rats. Molecules 2016, 21, 1517. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.; Coleman, E.; Dennis, J.; Akingbemi, B.; Schwartz, D.; Braden, T.; Judd, R.; Plaisance, E.; Stewart, L.K.; Morrison, E. Activation of PPAR by Rosiglitazone Does Not Negatively Impact Male Sex Steroid Hormones in Diabetic Rats. PPAR Res. 2009, 2009, 101857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yelumalai, S.; Giribabu, N.; Karim, K.; Omar, S.Z.; Bin Salleh, N. In vivo administration of quercetin ameliorates sperm oxidative stress, inflammation, preserves sperm morphology and functions in streptozotocin-nicotinamide induced adult male diabetic rats. Arch. Med. Sci. 2019, 15, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Taepongsorat, L.; Tangpraprutgul, P.; Kitana, N.; Malaivijitnond, S. Stimulating effects of quercetin on sperm quality and reproductive organs in adult male rats. Asian J. Androl. 2008, 10, 249–258. [Google Scholar] [CrossRef]
- Naseer, Z.; Ahmad, E.; Aksoy, M.; Epikmen, E. Impact of quercetin supplementation on testicular functions in summer heat-stressed rabbits. World Rabbit Sci. 2020, 28, 19–27. [Google Scholar] [CrossRef]
- Maresch, C.C.; Stute, D.C.; Alves, M.G.; Oliveira, P.F.; de Kretser, D.M.; Linn, T. Diabetes-induced hyperglycemia impairs male reproductive function: A systematic review. Hum. Reprod. Updat. 2017, 24, 86–105. [Google Scholar] [CrossRef] [Green Version]
- Sadek, K.M.; Lebda, M.A.; Nasr, S.M.; Shoukry, M. Spirulina platensis prevents hyperglycemia in rats15 by modulating gluconeogenesis and apoptosis via modification of oxidative stress and MAPK-16 pathways. Biomed. Pharmacother. 2017, 92, 1085–1094. [Google Scholar] [CrossRef]
- Moazamian, R.; Polhemus, A.; Connaughton, H.; Fraser, B.; Whiting, S.; Gharagozloo, P.; Aitken, R.J. Oxidative stress and human spermatozoa: Diagnostic and functional significance of aldehydes generated as a result of lipid peroxidation. Mol. Hum. Reprod. 2015, 21, 502–515. [Google Scholar] [CrossRef] [Green Version]
- Koh, P.-O. Streptozotocin-Induced Diabetes Increases Apoptosis through JNK Phosphorylation and Bax Activation in Rat Testes. J. Vet. Med. Sci. 2007, 69, 969–971. [Google Scholar] [CrossRef] [Green Version]
- Koh, P.-O. Streptozotocin-induced diabetes increases the interaction of Bad/Bcl-XL and decreases the binding of pBad/14–3–3 in rat testis. Life Sci. 2007, 81, 1079–1084. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Cabler, S.; McAlister, D.A.; Sabanegh, E.; Agarwal, A. The effect of obesity on sperm disorders and male infertility. Nat. Rev. Urol. 2010, 7, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Abu Bakar, A.B.; Ahmad, A.; Eleazu, C.O.; Mohamed, M. Oxidative Stress, NF-κB-Mediated Inflammation and Apoptosis in the Testes of Streptozotocin–Induced Diabetic Rats: Combined Protective Effects of Malaysian Propolis and Metformin. Antioxidants 2019, 8, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aprioku, J.S. Pharmacology of free radicals and the impact of reactive oxygen species on the testis. J. Reprod. Infertil. 2013, 14, 158–172. [Google Scholar] [PubMed]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, A.D.; Manson, J.E.; Rifai, N.; Buring, J.E.; Ridker, P.M. C-reactive protein, interleukin 6, and risk of developing type 2 diabetes mellitus. JAMA 2001, 286, 327–334. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef]
- El Husseny, M.W.; Mamdouh, M.; Shaban, S.; Ibrahim Abushouk, A.; Zaki, M.M.; Ahmed, O.M.; Abdel-Daim, M.M. Adipokines: Potential Therapeutic Targets for Vascular Dysfunction in Type II Diabetes Mellitus and Obesity. J. Diabetes Res. 2017, 2017, 8095926. [Google Scholar] [CrossRef]
- Hasan, H.; Bhushan, S.; Fijak, M.; Meinhardt, A. Mechanism of Inflammatory Associated Impairment of Sperm Function, Spermatogenesis and Steroidogenesis. Front. Endocrinol. 2022, 13, 897029. [Google Scholar] [CrossRef]
- Chojnacka, K.; Bilinska, B.; Mruk, D.D. Interleukin 1alpha-induced disruption of the Sertoli cell cytoskeleton affects gap junctional communication. Cell. Signal. 2016, 28, 469–480. [Google Scholar] [CrossRef]
- Zhang, H.; Yin, Y.; Wang, G.; Liu, Z.; Liu, L.; Sun, F. Interleukin-6 disrupts blood-testis barrier through inhibiting protein degradation or activating phosphorylated ERK in Sertoli cells. Sci. Rep. 2014, 4, 4260. [Google Scholar] [CrossRef] [Green Version]
- Boraschi, D.; Dinarello, C.A. IL-18 in autoimmunity: Review. Eur. Cytokine Netw. 2006, 17, 224–252. [Google Scholar] [PubMed]
- Havrylyuk, A.; Chopyak, V.; Boyko, Y.; Kril, I.; Kurpisz, M. Cytokines in the blood and semen of infertile patients. Cent. Eur. J. Immunol. 2015, 40, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Komsky, A.; Huleihel, M.; Ganaiem, M.; Kasterstein, E.; Komorovsky, D.; Bern, O.; Raziel, A.; Friedler, S.; Ron-El, R.; Strassburger, D. Presence of IL-18 in testicular tissue of fertile and infertile men. Andrologia 2012, 44, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Galvez, J.; de la Cruz, J.P.; Zarzuelo, A.; Sanchez de Medina, F.; Jimenez, J., Jr.; Sanchez de la Cuesta, F. Oral administration of quercitrin modifies intestinal oxidative status in rats. Gen. Pharmacol. 1994, 25, 1237–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comalada, M.; Camuesco, D.; Sierra, S.; Ballester, I.; Xaus, J.; Gálvez, J.; Zarzuelo, A. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-kappaB pathway. Eur. J. Immunol. 2005, 35, 584–592. [Google Scholar] [CrossRef]
- Rogler, G.; Brand, K.; Vogl, D.; Page, S.; Hofmeister, R.; Andus, T.; Knuechel, R.; Baeuerle, P.A.; Schölmerich, J.; Gross, V. Nuclear factor kappaB is activated in macrophages and epithelial cells of inflamed intestinal mucosa. Gastroenterology 1998, 115, 357–369. [Google Scholar] [CrossRef]
- Endale, M.; Park, S.C.; Kim, S.; Kim, S.H.; Yang, Y.; Cho, J.Y.; Rhee, M.H. Quercetin disrupts tyrosine-phosphorylated phosphatidylinositol 3-kinase and myeloid differentiation factor-88 association, and inhibits MAPK/AP-1 and IKK/NF-κB-induced inflammatory mediators production in RAW 264.7 cells. Immunobiology 2013, 218, 1452–1467. [Google Scholar] [CrossRef]
- Nikfarjam, B.A.; Hajiali, F.; Adineh, M.; Nassiri-Asl, M. Anti-inflammatory Effects of Quercetin and Vitexin on Activated Human Peripheral Blood Neutrophils. J. Pharmacopunct. 2017, 20, 127–131. [Google Scholar] [CrossRef]
- Ma, Z.; Ji, W.; Fu, Q.; Ma, S. Formononetin inhibited the inflammation of LPS-induced acute lung injury in mice associated with induction of PPAR gamma expression. Inflammation 2013, 36, 1560–1566. [Google Scholar] [CrossRef]
- Bhaskar, S.; Sudhakaran, P.R.; Helen, A. Quercetin attenuates atherosclerotic inflammation and adhesion molecule expression by modulating TLR-NF-κB signaling pathway. Cell. Immunol. 2016, 310, 131–140. [Google Scholar] [CrossRef]
- Pečivová, J.; Mačičková, T.; Sviteková, K.; Nosáľ, R. Quercetin inhibits degranulation and superoxide generation in PMA stimulated neutrophils. Interdiscip. Toxicol. 2012, 5, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Pfeuffer, M.; Auinger, A.; Bley, U.; Kraus-Stojanowic, I.; Laue, C.; Winkler, P.; Rüfer, C.; Frank, J.; Bösch-Saadatmandi, C.; Rimbach, G.; et al. Effect of quercetin on traits of the metabolic syndrome, endothelial function and inflammation in men with different APOE isoforms. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.R.; Lin, J.Y. Quercetin intraperitoneal administration ameliorates lipopolysaccharide-induced systemic inflammation in mice. Life Sci. 2015, 137, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Shaha, C.; Tripathi, R.; Mishra, D.P. Male germ cell apoptosis: Regulation and biology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1501–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, W.; Bai, Y.; Xu, L.; Liu, Y.; Yang, Z.; Gao, H.; Li, J. Curcumin Attenuates Testicular Injury in Rats with Streptozotocin-Induced Diabetes. Biomed Res. Int. 2018, 2018, 7468019. [Google Scholar] [CrossRef]
- Zhao, Y.; Tan, Y.; Dai, J.; Li, B.; Guo, L.; Cui, J.; Wang, G.; Shi, X.; Zhang, X.; Mellen, N.; et al. Exacerbation of diabetes-induced testicular apoptosis by zinc deficiency is most likely associated with oxidative stress, p38 MAPK activation, and p53 activation in mice. Toxicol. Lett. 2011, 200, 100–106. [Google Scholar] [CrossRef]
- He, W.; Liu, H.; Hu, L.; Wang, Y.; Huang, L.; Liang, A.; Wang, X.; Zhang, Q.; Chen, Y.; Cao, Y.; et al. Icariin improves testicular dysfunction via enhancing proliferation and inhibiting mitochondria-dependent apoptosis pathway in high-fat diet and streptozotocin-induced diabetic rats. Reprod. Biol. Endocrinol. 2021, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Kitamura, M. Anti-apoptotic effect of quercetin: Intervention in the JNK- and ERK-mediated apoptotic pathways. Kidney Int. 2000, 58, 1078–1087. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef]
- Nait Chabane, M.; Al Ahmad, A.; Peluso, J.; Muller, C.D.; Ubeaud, G. Quercetin and naringenin transport across human intestinal Caco-2 cells. J. Pharm. Pharmacol. 2009, 61, 1473–1483. [Google Scholar] [CrossRef]
- Li, H.; Li, M.; Fu, J.; Ao, H.; Wang, W.; Wang, X. Enhancement of oral bioavailability of quercetin by metabolic inhibitory nanosuspensions compared to conventional nanosuspensions. Drug Deliv. 2021, 28, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodi, F.; Jimenez, R.; Moreno, L.; Kroon, P.A.; Needs, P.W.; Hughes, D.A.; Santos-Buelga, C.; Gonzalez-Paramas, A.; Cogol-ludo, A.; Lopez-Sepulveda, R.; et al. Glucuronidated and sulfated metabolites of the flavonoid quercetin prevent endothelial dysfunction but lack direct vasorelaxant effects in rat aorta. Atherosclerosis 2009, 204, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Jasenovec, T.; Radosinska, D.; Kollarova, M.; Balis, P.; Ferenczyova, K.; Kalocayova, B.; Bartekova, M.; Tothova, L.; Radosinska, J. Beneficial Effect of Quercetin on Erythrocyte Properties in Type 2 Diabetic Rats. Molecules 2021, 26, 4868. [Google Scholar] [CrossRef]
- Ferenczyova, K.; Kalocayova, B.; Kindernay, L.; Jelemensky, M.; Balis, P.; Berenyiova, A.; Zemancikova, A.; Farkasova, V.; Sykora, M.; Tothova, L.; et al. Quercetin Exerts Age-Dependent Beneficial Effects on Blood Pressure and Vascular Function, But Is Inefficient in Preventing Myocardial Ischemia-Reperfusion Injury in Zucker Diabetic Fatty Rats. Molecules 2020, 25, 187. [Google Scholar] [CrossRef] [Green Version]
- Barteková, M.; Šimončíková, P.; Fogarassyová, M.; Ivanová, M.; Okruhlicová, Ľ.; Tribulová, N.; Dovinová, I.; Barančík, M. Quercetin Improves Postischemic Recovery of Heart Function in Doxorubicin-Treated Rats and Prevents Doxorubicin-Induced Matrix Metalloproteinase-2 Activation and Apoptosis Induction. Int. J. Mol. Sci. 2015, 16, 8168–8185. [Google Scholar] [CrossRef] [Green Version]
- Muller, C.H.; Lee, T.K.; Montano, M.A. Improved chemiluminescence assay for measuring antioxidant capacity of seminal plasma. Methods Mol. Biol. 2013, 927, 363–376. [Google Scholar] [CrossRef]
- Weber, D.; Davies, M.J.; Grune, T. Determination of protein carbonyls in plasma, cell extracts, tissue homogenates, isolated proteins: Focus on sample preparation and derivatization conditions. Redox Biol. 2015, 5, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Kovacik, A.; Tirpak, F.; Tomka, M.; Miskeje, M.; Tvrda, E.; Arvay, J.; Andreji, J.; Slanina, T.; Gabor, M.; Hleba, L.; et al. Trace elements content in semen and their interactions with sperm quality and RedOx status in freshwater fish Cyprinus carpio: A correlation study. J. Trace Elements Med. Biol. 2018, 50, 399–407. [Google Scholar] [CrossRef]
- Beers, R.F., Jr.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Benko, F.; Geschwandtnerová, A.; Žiarovská, J.; Lukáč, N.; Tvrdá, E. Expression of HSP90 Gene in the Cryopreserved Bovine Spermatozoa. Sci. Papers Anim. Sci. Biotechnol. 2021, 54, 73–77. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Protein | Antibody | Clonality/Isotype | Dilution | Blocking Solution | Source | ID | Manufacturer |
|---|---|---|---|---|---|---|---|
| BAX | anti-BAX antibody (BCL2-Associated X Protein) N-Term | Polyclonal/IgG | 1:1000 | 5% milk in TBS/0.1% Tween-20 | rabbit | #ABIN6990475 | Antibodies Online; Dunwoody, GA, USA |
| Bcl-2 | anti-Bcl-2 antibody (B-Cell CLL/lymphoma 2) N-Term | Polyclonal/IgG | 1:1000 | 5% milk in TBS/0.1% Tween-20 | rabbit | #ABIN2857047 | Antibodies Online; Dunwoody, GA, USA |
| p53 | anti-p53 antibody [PAb421] | Monoclonal/IgG | 1:1000 | 5% milk in TBS/0.1% Tween-20 | rabbit | #ab245685 | Abcam, Cambridge, UK |
| Caspase-3 | Caspase 3 (Cleaved Asp175) | Polyclonal/IgG | 1:1000 | 5% milk in TBS/0.1% Tween-20 | rabbit | #PA5-114687 | Invitrogen, Waltham, MA, USA |
| β-actin | beta Actin Polyclonal Antibody | Polyclonal/IgG | 1:1000 | 5% milk in TBS/0.1% Tween-20 | rabbit | #PA1-16889 | Invitrogen, Waltham, MA, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tvrdá, E.; Kováč, J.; Ferenczyová, K.; Kaločayová, B.; Ďuračka, M.; Benko, F.; Almášiová, V.; Barteková, M. Quercetin Ameliorates Testicular Damage in Zucker Diabetic Fatty Rats through Its Antioxidant, Anti-Inflammatory and Anti-Apoptotic Properties. Int. J. Mol. Sci. 2022, 23, 16056. https://doi.org/10.3390/ijms232416056
Tvrdá E, Kováč J, Ferenczyová K, Kaločayová B, Ďuračka M, Benko F, Almášiová V, Barteková M. Quercetin Ameliorates Testicular Damage in Zucker Diabetic Fatty Rats through Its Antioxidant, Anti-Inflammatory and Anti-Apoptotic Properties. International Journal of Molecular Sciences. 2022; 23(24):16056. https://doi.org/10.3390/ijms232416056
Chicago/Turabian StyleTvrdá, Eva, Ján Kováč, Kristína Ferenczyová, Barbora Kaločayová, Michal Ďuračka, Filip Benko, Viera Almášiová, and Monika Barteková. 2022. "Quercetin Ameliorates Testicular Damage in Zucker Diabetic Fatty Rats through Its Antioxidant, Anti-Inflammatory and Anti-Apoptotic Properties" International Journal of Molecular Sciences 23, no. 24: 16056. https://doi.org/10.3390/ijms232416056