

Characterisation of the Molecular Mechanism of Permeation of the Prodrug Me-5ALA across the Human Stratum Corneum Using Molecular Dynamics Simulations
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Predicted Free Energy Profiles
2.2. Diffusivity Profiles
2.3. Transdermal Permeability
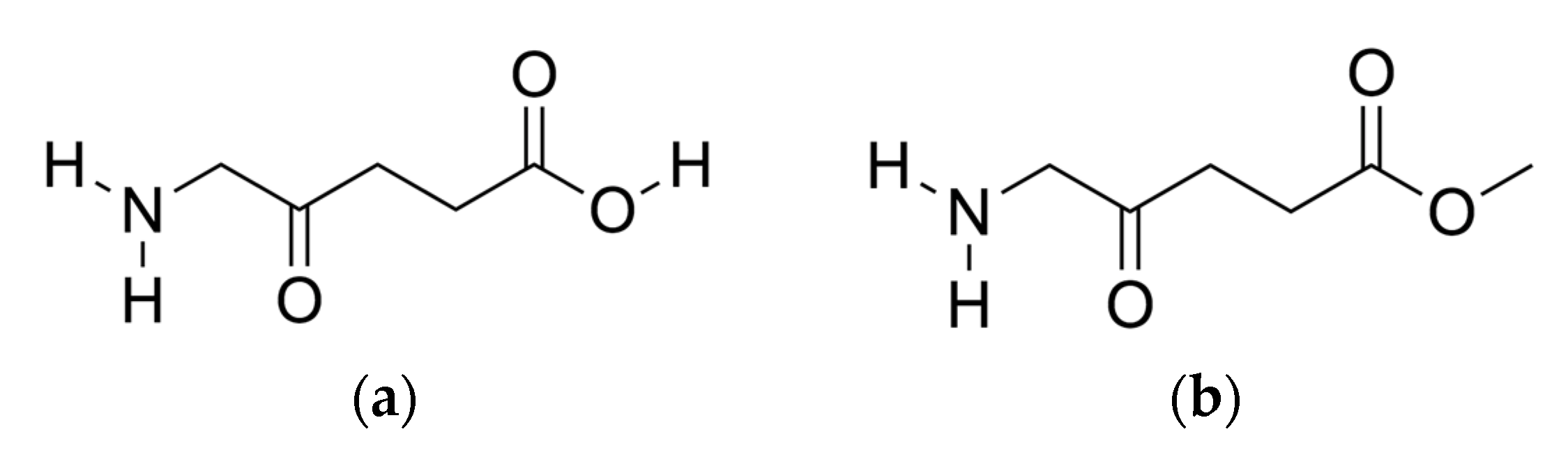
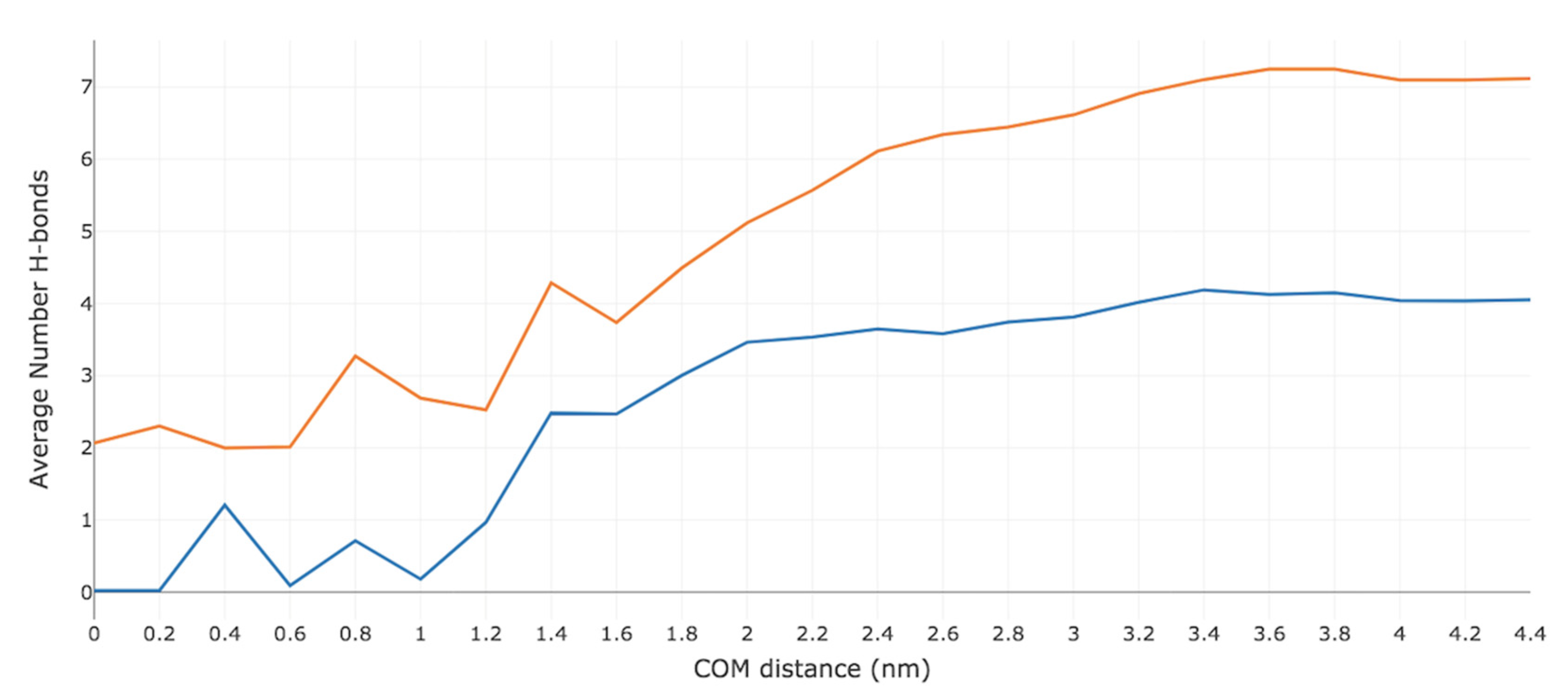
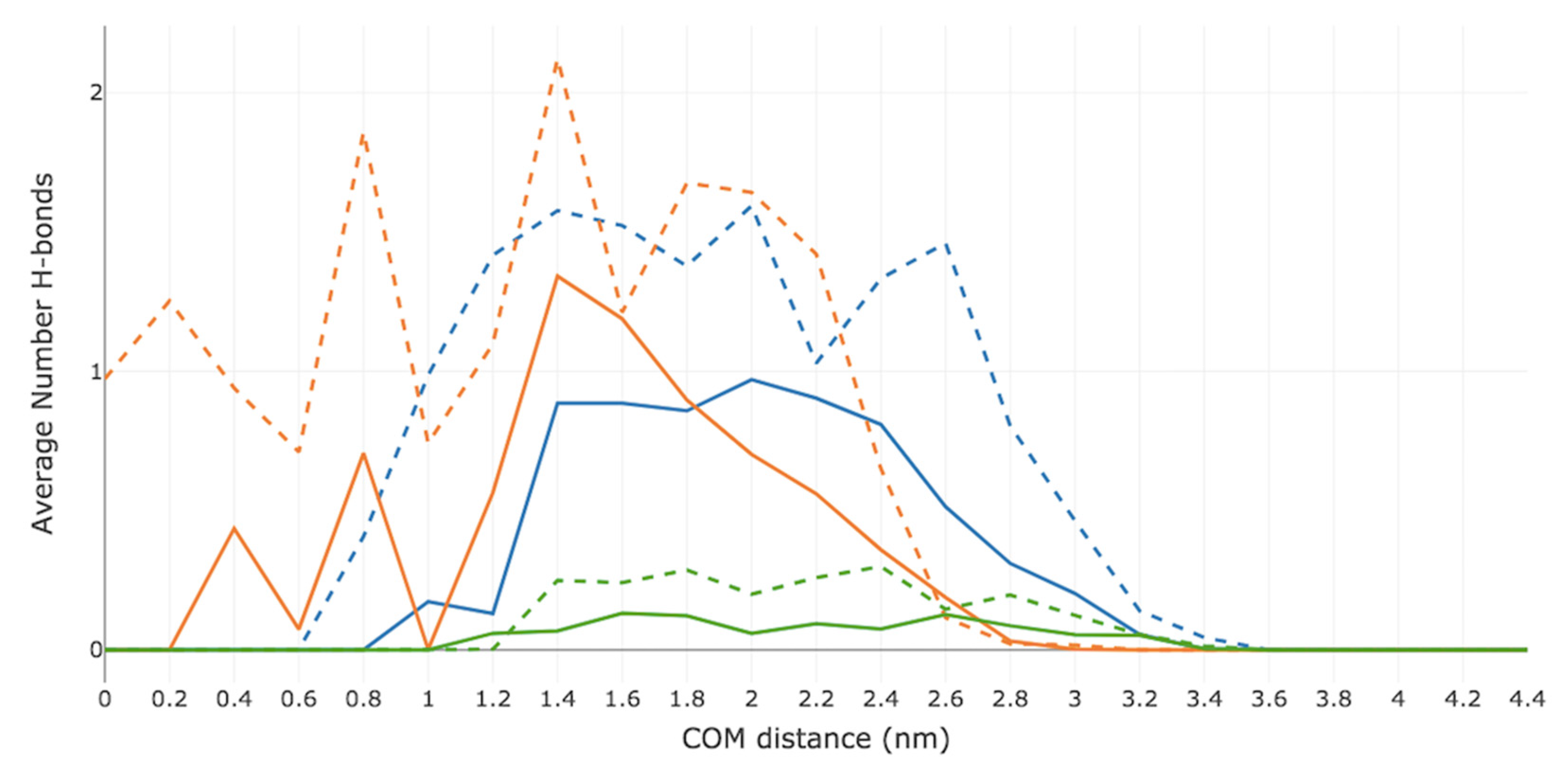
2.4. Hydrogen-Bonding Analysis
3. Discussion
4. Materials and Methods
4.1. Simulation System
4.2. Insertion of 5ALA and Me-5ALA in the Simulation System
4.3. Pull Simulation
4.4. Umbrella Sampling and Selective Replica Exchange with Solute Tempering
4.5. Convergence of the Free Energy of Permeation
4.6. Estimation of Diffusivity Profiles
4.7. Permeability Coefficient and Transdermal Permeability
4.8. Hydrogen-Bonding Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| US Window (COM Distance/nm) | Replica Scheme | Time (ns) | |
|---|---|---|---|
| No. of Replicas | Scaling Factors | ||
| 0.0 | 5 | 1.00, 0.88, 0.75, 0.63, 0.50 | 1250 |
| 0.2 | 5 | 1.00, 0.88, 0.75, 0.63, 0.50 | 1250 |
| 0.4 | 5 | 1.00, 0.88, 0.75, 0.63, 0.50 | 1250 |
| 0.6 | 5 | 1.00, 0.88, 0.75, 0.63, 0.50 | 1250 |
| 0.8 | 11 | 1.00, 0.91, 0.82, 0.73, 0.64, 0.55, 0.46, 0.37, 0.28, 0.19, 0.10 | 2750 |
| 1.0 | 11 | 1.00, 0.91, 0.82, 0.73, 0.64, 0.55, 0.46, 0.37, 0.28, 0.19, 0.10 | 2750 |
| 1.2 | 11 | 1.00, 0.91, 0.82, 0.73, 0.64, 0.55, 0.46, 0.37, 0.28, 0.19, 0.10 | 2750 |
| 1.4 | 11 | 1.00, 0.91, 0.82, 0.73, 0.64, 0.55, 0.46, 0.37, 0.28, 0.19, 0.10 | 2750 |
| 1.6 | 11 | 1.00, 0.91, 0.82, 0.73, 0.64, 0.55, 0.46, 0.37, 0.28, 0.19, 0.10 | 2750 |
| 1.8 | 11 | 1.00, 0.91, 0.82, 0.73, 0.64, 0.55, 0.46, 0.37, 0.28, 0.19, 0.10 | 2750 |
| 2.0 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 2.2 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 2.4 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 2.6 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 2.8 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 3.0 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 3.2 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 3.4 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 3.6 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 3.8 | 10 | 1.00, 0.90, 0.80, 0.70, 0.60, 0.50, 0.40, 0.30, 0.20, 0.10 | 2500 |
| 4.0 | 0 | NA | 250 |
| 4.2 | 0 | NA | 250 |
| 4.4 | 0 | NA | 250 |
References
- Roberts, M.S.; Cheruvu, H.S.; Mangion, S.E.; Alinaghi, A.; Benson, H.A.E.; Mohammed, Y.; Holmes, A.; van der Hoek, J.; Pastore, M.; Grice, J.E. Topical Drug Delivery: History, Percutaneous Absorption, and Product Development. Adv. Drug Deliv. Rev. 2021, 177, 113929. [Google Scholar] [CrossRef] [PubMed]
- Benson, H.A.E.; Grice, J.E.; Mohammed, Y.; Namjoshi, S.; Roberts, M.S. Topical and Transdermal Drug Delivery: From Simple Potions to Smart Technologies. Curr. Drug Deliv. 2019, 16, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Scheuplein, J.; Blank, H. Permeability of the Skin. Physiol. Rev. 1971, 51, 46. [Google Scholar] [CrossRef] [PubMed]
- van Smeden, J.; Janssens, M.; Gooris, G.S.; Bouwstra, J.A. The Important Role of Stratum Corneum Lipids for the Cutaneous Barrier Function. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2014, 1841, 295–313. [Google Scholar] [CrossRef]
- van Smeden, J.; Boiten, W.A.; Hankemeier, T.; Rissmann, R.; Bouwstra, J.A.; Vreeken, R.J. Combined LC/MS-Platform for Analysis of All Major Stratum Corneum Lipids, and the Profiling of Skin Substitutes. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2014, 1841, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Paudel, K.S.; Milewski, M.; Swadley, C.L.; Brogden, N.K.; Ghosh, P.; Stinchcomb, A.L. Challenges and Opportunities in Dermal/Transdermal Delivery. Ther. Deliv. 2010, 1, 109–131. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.C.; Barry, B.W. Penetration Enhancers. Adv. Drug Deliv. Rev. 2012, 64, 128–137. [Google Scholar] [CrossRef]
- El Maghraby, G.M.; Barry, B.W.; Williams, A.C. Liposomes and Skin: From Drug Delivery to Model Membranes. Eur. J. Pharm. Sci. 2008, 34, 203–222. [Google Scholar] [CrossRef]
- Zhang, L.; Falla, T.J. Cosmeceuticals and Peptides. Clin. Dermatol. 2009, 27, 485–494. [Google Scholar] [CrossRef]
- Caccetta, R.; Blanchfield, J.T.; Harrison, J.; Toth, I.; Benson, H.A.E. Epidermal Penetration of a Therapeutic Peptide by Lipid Conjugation; Stereo-Selective Peptide Availability of a Topical Diastereomeric Lipopeptide. Int. J. Pept. Res. Ther. 2006, 12, 327–333. [Google Scholar] [CrossRef]
- Benson, H.A.E.; Namjoshi, S. Proteins and Peptides: Strategies for Delivery to and Across the Skin. J. Pharm. Sci. 2008, 97, 3591–3610. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.A.; Szeimies, R.M.; Basset-Seguin, N.; Calzavara-Pinton, P.; Gilaberte, Y.; Hædersdal, M.; Hofbauer, G.F.L.; Hunger, R.E.; Karrer, S.; Piaserico, S.; et al. European Dermatology Forum Guidelines on Topical Photodynamic Therapy 2019 Part 1: Treatment Delivery and Established Indications—Actinic Keratoses, Bowen’s Disease and Basal Cell Carcinomas. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 2225–2238. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.A.; Szeimies, R.M.; Basset-Séguin, N.; Calzavara-Pinton, P.G.; Gilaberte, Y.; Hædersdal, M.; Hofbauer, G.F.L.; Hunger, R.E.; Karrer, S.; Piaserico, S.; et al. European Dermatology Forum Guidelines on Topical Photodynamic Therapy 2019 Part 2: Emerging Indications—Field Cancerization, Photorejuvenation and Inflammatory/Infective Dermatoses. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Morton, C. Methyl Aminolevulinate (Metvix®) Photodynamic Therapy—Practical Pearls. J. Dermatolog. Treat. 2003, 14, 23–26. [Google Scholar] [CrossRef]
- Erdtman, E.; dos Santos, D.J.V.A.; Löfgren, L.; Eriksson, L.A. Modelling the Behavior of 5-Aminolevulinic Acid and Its Alkyl Esters in a Lipid Bilayer. Chem. Phys. Lett. 2008, 463, 178–182. [Google Scholar] [CrossRef]
- Yazdanyar, S.; Zarchi, K.; Jemec, G.B.E. Pain during Topical Photodynamic Therapy—Comparing Methyl Aminolevulinate (Metvix®) to Aminolaevulinic Acid (Ameluz®); an Intra-Individual Clinical Study. Photodiagnosis Photodyn. Ther. 2017, 20, 6–9. [Google Scholar] [CrossRef]
- Abd, E.; Yousuf, S.; Pastore, M.; Telaprolu, K.; Mohammed, Y.; Namjoshi, S.; Grice, J.; Roberts, M. Skin Models for the Testing of Transdermal Drugs. Clin. Pharmacol. Adv. Appl. 2016, 8, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Van Gunsteren, W.F.; Bakowies, D.; Baron, R.; Chandrasekhar, I.; Christen, M.; Daura, X.; Gee, P.; Geerke, D.P.; Glättli, A.; Hünenberger, P.H.; et al. Biomolecular Modeling: Goals, Problems, Perspectives. Angew. Chem. Int. Ed. 2006, 45, 4064–4092. [Google Scholar] [CrossRef]
- Torin Huzil, J.; Sivaloganathan, S.; Kohandel, M.; Foldvari, M. Drug Delivery through the Skin: Molecular Simulations of Barrier Lipids to Design More Effective Noninvasive Dermal and Transdermal Delivery Systems for Small Molecules, Biologics, and Cosmetics. WIREs Nanomed. Nanobiotechnol. 2011, 3, 449–462. [Google Scholar] [CrossRef]
- Notman, R.; Anwar, J. Breaching the Skin Barrier—Insights from Molecular Simulation of Model Membranes. Adv. Drug Deliv. Rev. 2013, 65, 237–250. [Google Scholar] [CrossRef]
- Lopes, D.; Jakobtorweihen, S.; Nunes, C.; Sarmento, B.; Reis, S. Shedding Light on the Puzzle of Drug-Membrane Interactions: Experimental Techniques and Molecular Dynamics Simulations. Prog. Lipid Res. 2017, 65, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Bretschko, E.; Szeimies, R.-M.; Landthaler, M.; Lee, G. Topical 5-Aminolevulinic Acid for Photodynamic Therapy of Basal Cell Carcinoma. Evaluation of Stratum Corneum Permeability in Vitro. J. Control. Release 1996, 42, 203–208. [Google Scholar] [CrossRef]
- Hadgraft, J.; Walters, K. Skin Penetration Enhancement. J. Dermatol. Treat. 1994, 5, 43–47. [Google Scholar] [CrossRef]
- Jhanker, Y.M. Formulation Development and Optimization for Transdermal Delivery of Peptide and Peptide Like Drugs. Ph.D. Thesis, Curtin University, Perth, Australia, 2018. [Google Scholar]
- Donnelly, R.F.; Morrow, D.I.J.; McCarron, P.A.; Woolfson, A.D.; Morrissey, A.; Juzenas, P.; Juzeniene, A.; Iani, V.; McCarthy, H.O.; Moan, J. Microneedle-Mediated Intradermal Delivery of 5-Aminolevulinic Acid: Potential for Enhanced Topical Photodynamic Therapy. J. Control. Release 2008, 129, 154–162. [Google Scholar] [CrossRef]
- Jhanker, Y.; Mbano, M.N.; Ponto, T.; Espartero, L.J.L.; Yamada, M.; Prow, T.; Benson, H.A.E. Comparison of Physical Enhancement Technologies in the Skin Permeation of Methyl Amino Levulinic Acid (MALA). Int. J. Pharm. 2021, 610, 121258. [Google Scholar] [CrossRef]
- Merclin, N.; Bramer, T.; Edsman, K. Iontophoretic Delivery of 5-Aminolevulinic Acid and Its Methyl Ester Using a Carbopol Gel as Vehicle. J. Control. Release 2004, 98, 57–65. [Google Scholar] [CrossRef]
- Krishnan, G.; Roberts, M.S.; Grice, J.; Anissimov, Y.G.; Benson, H.A.E. Enhanced Transdermal Delivery of 5-Aminolevulinic Acid and a Dipeptide by Iontophoresis. Biopolymers 2011, 96, 166–171. [Google Scholar] [CrossRef]
- Krishnan, G.; Grice, J.E.; Roberts, M.S.; Benson, H.A.E.; Prow, T.W. Enhanced Sonophoretic Delivery of 5-Aminolevulinic Acid: Preliminary Human Ex Vivo Permeation Data. Skin Res. Technol. 2013, 19, e283–e289. [Google Scholar] [CrossRef]
- Schmitz, L.; Novak, B.; Hoeh, A.-K.; Luebbert, H.; Dirschka, T. Epidermal Penetration and Protoporphyrin IX Formation of Two Different 5-Aminolevulinic Acid Formulations in Ex Vivo Human Skin. Photodiagnosis Photodyn. Ther. 2016, 14, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Maisch, T.; Santarelli, F.; Schreml, S.; Babilas, P.; Szeimies, R.-M. Fluorescence Induction of Protoporphyrin IX by a New 5-Aminolevulinic Acid Nanoemulsion Used for Photodynamic Therapy in a Full-Thickness Ex Vivo Skin Model: Letter to the Editor. Exp. Dermatol. 2009, 19, e302–e305. [Google Scholar] [CrossRef]
- Shinoda, W. Permeability across Lipid Membranes. Biochim. Biophys. Acta BBA Biomembr. 2016, 1858, 2254–2265. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, C.; Ruiz-Perez, L.; Deplazes, E.; Mancera, R.L. Molecular Dynamics Simulation of Small Molecules Interacting with Biological Membranes. ChemPhysChem 2020, 21, 1486–1514. [Google Scholar] [CrossRef] [PubMed]
- van Gunsteren, W.F.; Daura, X.; Hansen, N.; Mark, A.E.; Oostenbrink, C.; Riniker, S.; Smith, L.J. Validation of Molecular Simulation: An Overview of Issues. Angew. Chem. Int. Ed. 2018, 57, 884–902. [Google Scholar] [CrossRef] [PubMed]
- Decherchi, S.; Cavalli, A. Thermodynamics and Kinetics of Drug-Target Binding by Molecular Simulation. Chem. Rev. 2020, 120, 12788–12833. [Google Scholar] [CrossRef] [PubMed]
- Zorec, B.; Préat, V.; Miklavčič, D.; Pavšelj, N. Active Enhancement Methods for Intra- and Transdermal Drug Delivery: A Review. Zdr. Vestn. 2013, 82, 339–356. [Google Scholar]
- Hoopes, M.I.; Noro, M.G.; Longo, M.L.; Faller, R. Bilayer Structure and Lipid Dynamics in a Model Stratum Corneum with Oleic Acid. J. Phys. Chem. B 2011, 115, 3164–3171. [Google Scholar] [CrossRef]
- Das, C.; Olmsted, P.D. The Physics of Stratum Corneum Lipid Membranes. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 2016, 374, 20150126. [Google Scholar] [CrossRef] [Green Version]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. PACKMOL: A Package for Building Initial Configurations for Molecular Dynamics Simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef]
- Poger, D.; Van Gunsteren, W.F.; Mark, A.E. A New Force Field for Simulating Phosphatidylcholine Bilayers. J. Comput. Chem. 2010, 31, 1117–1125. [Google Scholar] [CrossRef]
- Poger, D.; Mark, A.E. On the Validation of Molecular Dynamics Simulations of Saturated and Cis-Monounsaturated Phosphatidylcholine Lipid Bilayers: A Comparison with Experiment. J. Chem. Theory Comput. 2010, 6, 325–336. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Hermans, J. Interaction Models for Water in Relation to Protein Hydration. In Intermolecular Forces; Pullman, B., Ed.; The Jerusalem Symposia on Quantum Chemistry and Biochemistry; Springer: Dordrecht, The Netherlands, 1981; Volume 14, pp. 331–342. [Google Scholar] [CrossRef]
- Malde, A.K.; Zuo, L.; Breeze, M.; Stroet, M.; Poger, D.; Nair, P.C.; Oostenbrink, C.; Mark, A.E. An Automated Force Field Topology Builder (ATB) and Repository: Version 1.0. J. Chem. Theory Comput. 2011, 7, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical Dynamics: Equilibrium Phase-Space Distributions. Phys. Rev. A 1985, 31, 1695–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Kim, B.; Friesner, R.A.; Berne, B.J. Replica Exchange with Solute Tempering: A Method for Sampling Biological Systems in Explicit Water. Proc. Natl. Acad. Sci. USA 2005, 102, 13749–13754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Friesner, R.A.; Berne, B.J. Replica Exchange with Solute Scaling: A More Efficient Version of Replica Exchange with Solute Tempering (REST2). J. Phys. Chem. B 2011, 115, 9431–9438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hub, J.S.; de Groot, B.L.; van der Spoel, D. G_wham—A Free Weighted Histogram Analysis Implementation Including Robust Error and Autocorrelation Estimates. J. Chem. Theory Comput. 2010, 6, 3713–3720. [Google Scholar] [CrossRef] [Green Version]
- Hummer, G. Position-Dependent Diffusion Coefficients and Free Energies from Bayesian Analysis of Equilibrium and Replica Molecular Dynamics Simulations. New J. Phys. 2005, 7, 34. [Google Scholar] [CrossRef]
- Carpenter, T.S.; Kirshner, D.A.; Lau, E.Y.; Wong, S.E.; Nilmeier, J.P.; Lightstone, F.C. A Method to Predict Blood-Brain Barrier Permeability of Drug-Like Compounds Using Molecular Dynamics Simulations. Biophys. J. 2014, 107, 630–641. [Google Scholar] [CrossRef]







| Molecule | Sample Type | Method | P (nm/ps) |
|---|---|---|---|
| 5ALA | Human thigh skin | Horizontal-type diffusion cell | 2.25 × 10−13 |
| 5ALA | Human abdomen skin | Franz-type diffusion cell | 2.36 × 10−13 |
| Me-5ALA | Piglet skin | Franz-type diffusion cell | 4.17 × 10−13 (Solution) 7.50 × 10−13 (Gel) |
| 5ALA | DPPC bilayer | MD simulation | 1.89 × 10−4 |
| Me-5ALA | DPPC bilayer | MD simulation | 5.28 × 10−4 |
| 5ALA | SC lipid bilayer | MD simulation (this work) | 6.58 × 10−12 |
| Me-5ALA | SC lipid bilayer | MD simulation (this work) | 2.62 × 10−11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadyrov, J.; Ruiz-Perez, L.; Benson, H.A.E.; Mancera, R.L. Characterisation of the Molecular Mechanism of Permeation of the Prodrug Me-5ALA across the Human Stratum Corneum Using Molecular Dynamics Simulations. Int. J. Mol. Sci. 2022, 23, 16001. https://doi.org/10.3390/ijms232416001
Kadyrov J, Ruiz-Perez L, Benson HAE, Mancera RL. Characterisation of the Molecular Mechanism of Permeation of the Prodrug Me-5ALA across the Human Stratum Corneum Using Molecular Dynamics Simulations. International Journal of Molecular Sciences. 2022; 23(24):16001. https://doi.org/10.3390/ijms232416001
Chicago/Turabian StyleKadyrov, Janonna, Lanie Ruiz-Perez, Heather A. E. Benson, and Ricardo L. Mancera. 2022. "Characterisation of the Molecular Mechanism of Permeation of the Prodrug Me-5ALA across the Human Stratum Corneum Using Molecular Dynamics Simulations" International Journal of Molecular Sciences 23, no. 24: 16001. https://doi.org/10.3390/ijms232416001