
Deep Brain Stimulation in the Subthalamic Nucleus Can Improve Skilled Forelimb Movements and Retune Dynamics of Striatal Networks in a Rat Stroke Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
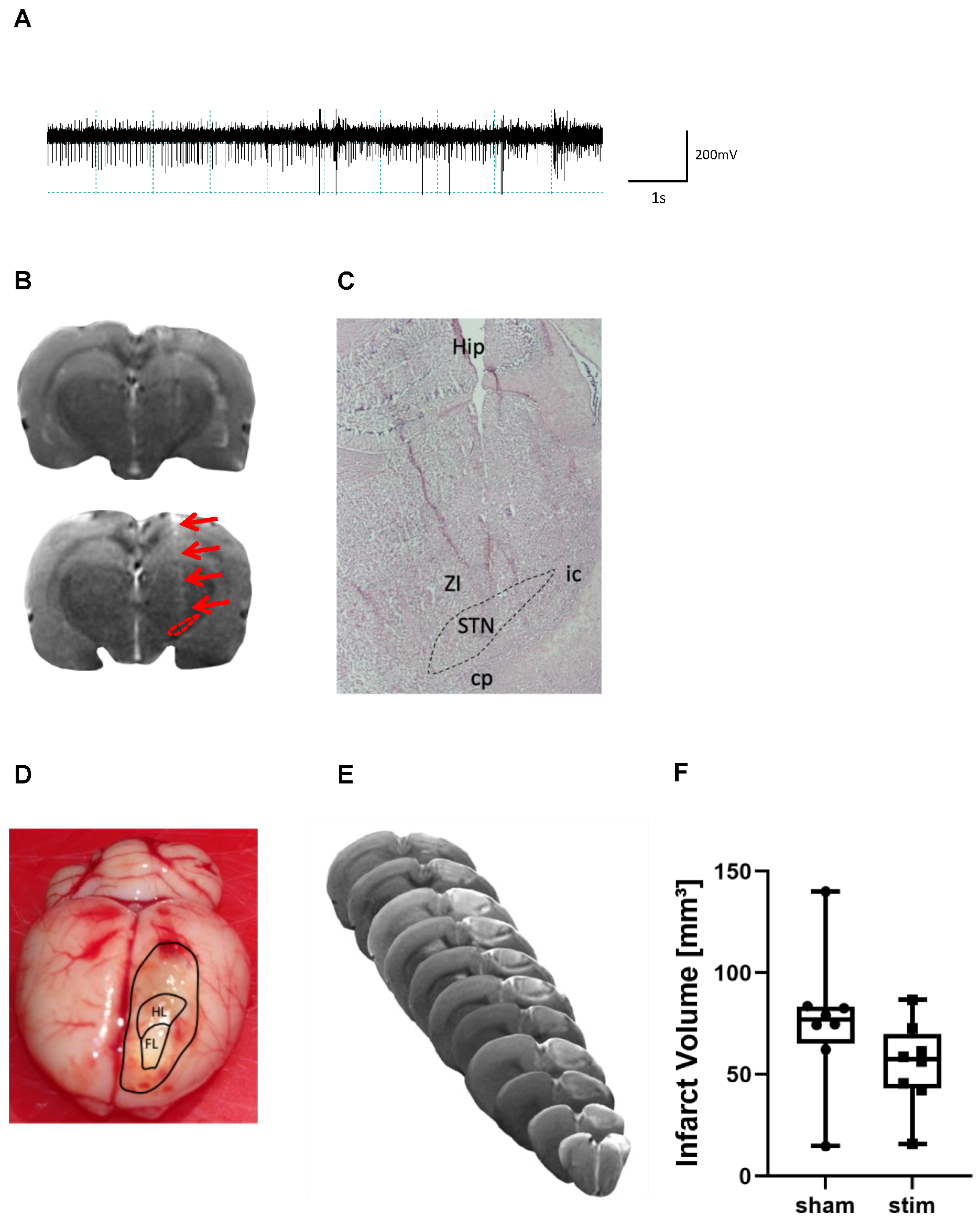
2.1. Determination of Electrode Placement and Lesion Volume
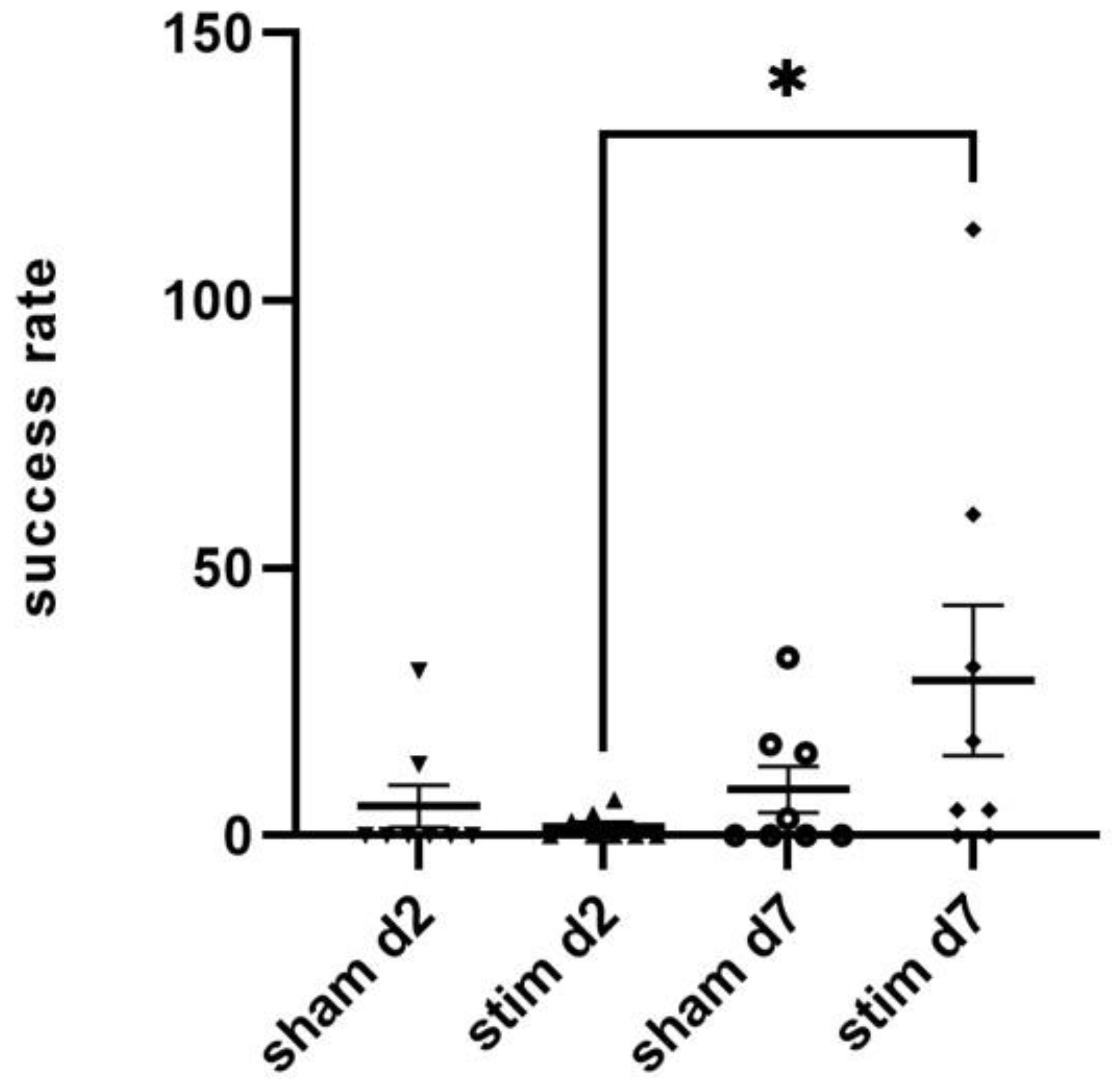
2.2. Single-Pellet Reaching Test (SPRT)
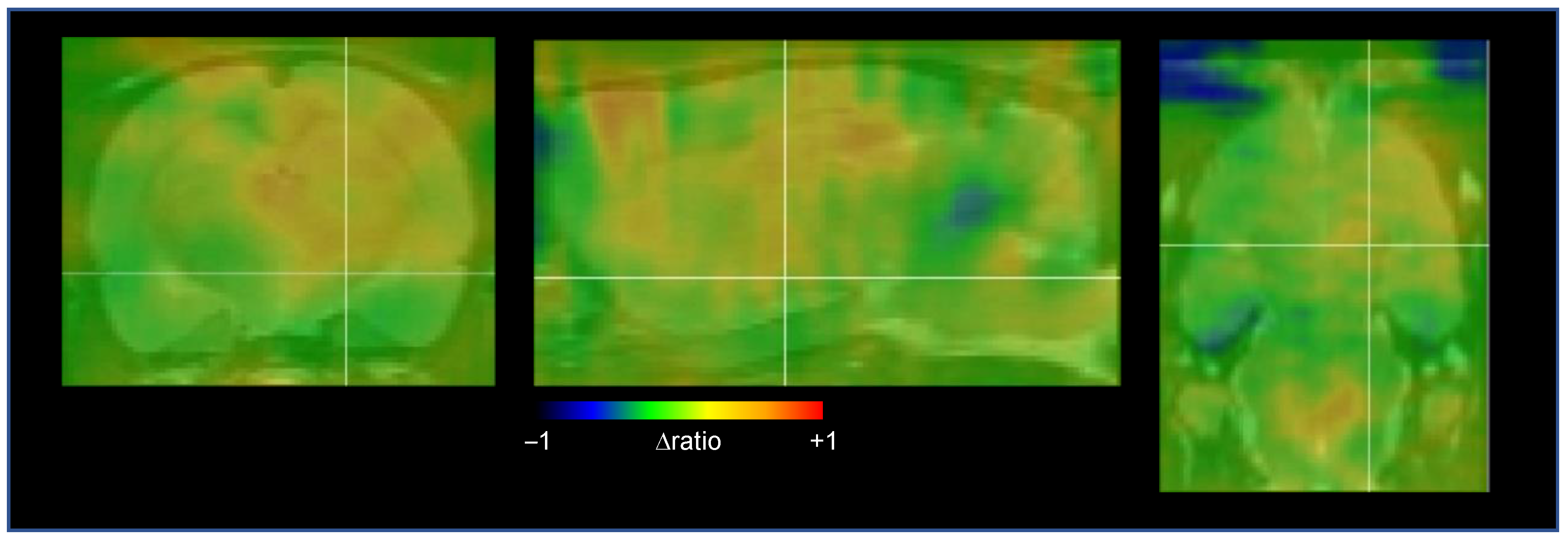
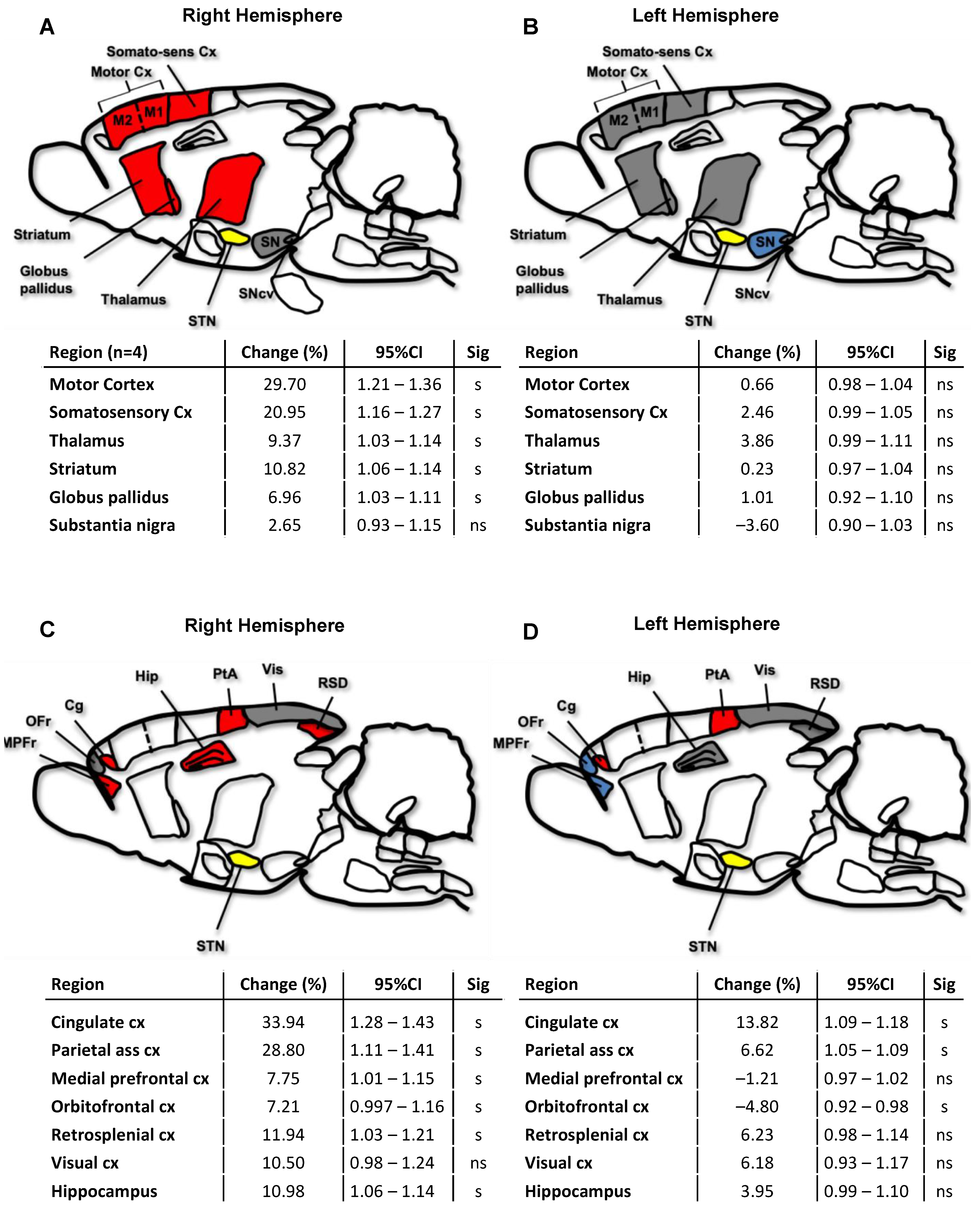
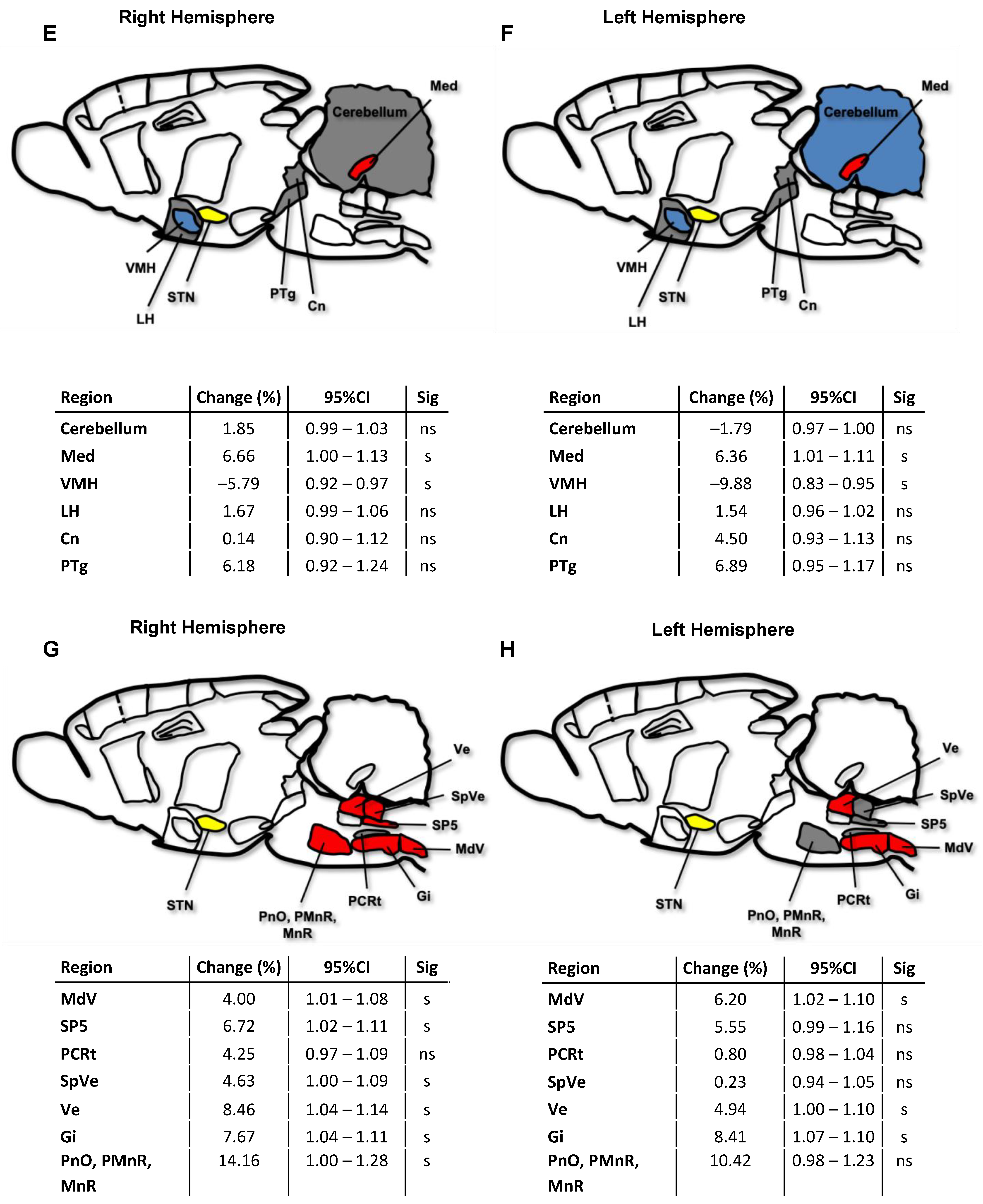
2.3. Cerebral Network Activation after STN-HFS Using PET Analyses
3. Discussion
4. Materials and Methods
4.1. Animals
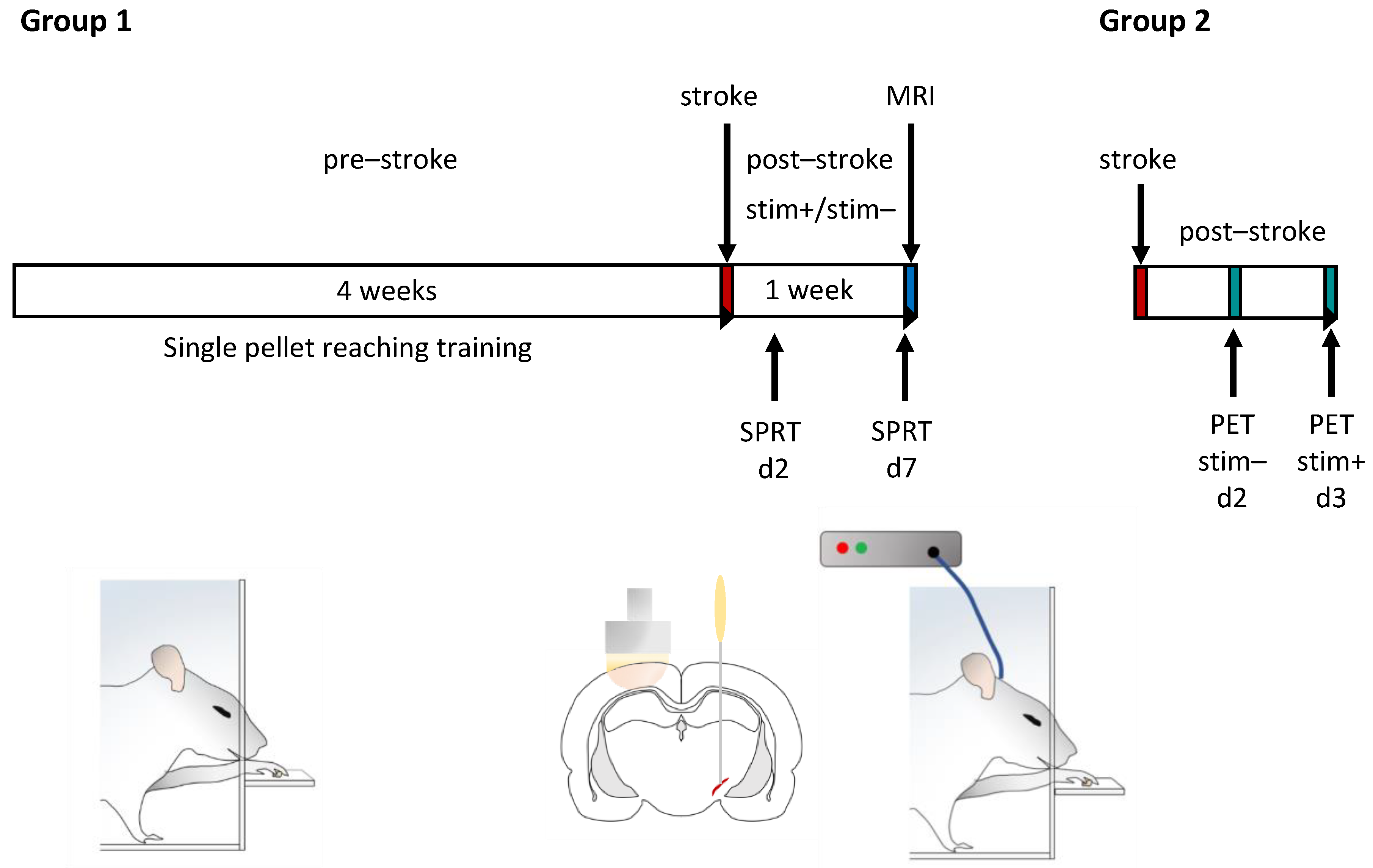
4.2. Experiment Design
4.3. Training and Testing of Skilled Forelimb Movements
4.4. Photothrombotic Stroke
4.5. Microelectrode Implantation
4.6. Stimulation of Subthalamic Nucleus
4.7. Radiochemistry
4.8. Small Animal Positron Emission Tomography (PET)
4.9. Evaluation of Glucose Metabolism in Different Brain Regions
4.10. Measurement of Photothrombotic Lesion Volume
4.11. Immunohistochemistry
4.12. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly-Hayes, M.; Beiser, A.; Kase, C.S.; Scaramucci, A.; D’Agostino, R.B.; Wolf, P.A. The Influence of Gender and Age on Disability Following Ischemic Stroke: The Framingham Study. J. Stroke Cerebrovasc. Dis. 2003, 12, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Wade, D.T.; Langton-Hewer, R.; Wood, V.A.; Skilbeck, C.E.; Ismail, H.M. The Hemiplegic Arm after Stroke: Measurement and Recovery. J. Neurol. Neurosurg. Psychiatry 1983, 46, 521–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Ejaz, N.; Hertler, B.; Branscheidt, M.; Widmer, M.; Faria, A.V.; Harran, M.D.; Cortes, J.C.; Kim, N.; Celnik, P.A.; et al. Separable Systems for Recovery of Finger Strength and Control after Stroke. J. Neurophysiol. 2017, 118, 1151–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Latash, M.L.; Yue, G.H.; Siemionow, V.; Sahgal, V. The Effects of Stroke and Age on Finger Interaction in Multi-Finger Force Production Tasks. Clin. Neurophysiol. 2003, 114, 1646–1655. [Google Scholar] [CrossRef] [PubMed]
- Posner, M.I.; Petersen, S.E.; Fox, P.T.; Raichle, M.E. Localization of Cognitive Operations in the Human Brain. Science 1988, 240, 1627–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wernicke, C. Der Aphasische Symptomenkomplex. Eine Psychologische Studie Auf Anatomischer Basis.; M. Cohn & Weigert: Breslau, 1874. [Google Scholar]
- Broca, P. Remarques Sur Le Siége de La Faculté Du Langage Articulé Suivies d’une Observation d’aphémie (Pert de La Parole). Bull. Soc. Anat. 1861, 6, 330–357. [Google Scholar]
- van den Heuvel, M.P.; Sporns, O. Rich-Club Organization of the Human Connectome. J. Neurosci. 2011, 31, 15775–15786. [Google Scholar] [CrossRef] [Green Version]
- Bowren, M., Jr.; Bruss, J.; Manzel, K.; Edwards, D.; Liu, C.; Corbetta, M.; Tranel, D.; Boes, A.D. Post-Stroke Outcomes Predicted from Multivariate Lesion-Behaviour and Lesion Network Mapping. Brain 2022, 145, 1338–1353. [Google Scholar] [CrossRef]
- Siegel, J.S.; Shulman, G.L.; Corbetta, M. Mapping Correlated Neurological Deficits after Stroke to Distributed Brain Networks. Brain Struct. Funct. 2022. [Google Scholar] [CrossRef]
- Salvalaggio, A.; De Filippo De Grazia, M.; Zorzi, M.; Thiebaut de Schotten, M.; Corbetta, M. Post-Stroke Deficit Prediction from Lesion and Indirect Structural and Functional Disconnection. Brain 2020, 143, 2173–2188. [Google Scholar] [CrossRef]
- Ward, N.S.; Newton, J.M.; Swayne, O.B.C.; Lee, L.; Thompson, A.J.; Greenwood, R.J.; Rothwell, J.C.; Frackowiak, R.S.J. Motor System Activation after Subcortical Stroke Depends on Corticospinal System Integrity. Brain 2006, 129, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Stam, C.J. Modern Network Science of Neurological Disorders. Nat. Rev. Neurosci. 2014, 15, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.S.; Ramsey, L.E.; Snyder, A.Z.; Metcalf, N.V.; Chacko, R.V.; Weinberger, K.; Baldassarre, A.; Hacker, C.D.; Shulman, G.L.; Corbetta, M. Disruptions of Network Connectivity Predict Impairment in Multiple Behavioral Domains after Stroke. Proc. Natl. Acad. Sci. USA 2016, 201521083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, P.; Evangelista, G.G.; Koch, P.J.; Park, C.-H.; Levin-Gleba, L.; Girard, G.; Beanato, E.; Lee, J.; Choirat, C.; Guggisberg, A.G.; et al. Disconnectomics of the Rich Club Impacts Motor Recovery After Stroke. Stroke 2021, 52, 2115–2124. [Google Scholar] [CrossRef]
- Hamani, C.; Florence, G.; Heinsen, H.; Plantinga, B.R.; Temel, Y.; Uludag, K.; Alho, E.; Teixeira, M.J.; Amaro, E.; Fonoff, E.T. Subthalamic Nucleus Deep Brain Stimulation: Basic Concepts and Novel Perspectives. eneuro 2017, 4, ENEURO.0140-17.2017. [Google Scholar] [CrossRef] [Green Version]
- Macerollo, A.; Zrinzo, L.; Akram, H.; Foltynie, T.; Limousin, P. Subthalamic Nucleus Deep Brain Stimulation for Parkinson’s Disease: Current Trends and Future Directions. Expert Rev. Med. Devices 2020, 17, 1063–1074. [Google Scholar] [CrossRef]
- Lozano, A.M.; Lipsman, N.; Bergman, H.; Brown, P.; Chabardes, S.; Chang, J.W.; Matthews, K.; McIntyre, C.C.; Schlaepfer, T.E.; Schulder, M.; et al. Deep Brain Stimulation: Current Challenges and Future Directions. Nat. Rev. Neurol. 2019, 15, 148–160. [Google Scholar] [CrossRef]
- Volkmann, J.; Allert, N.; Voges, J.; Weiss, P.H.; Freund, H.-J.; Sturm, V. Safety and Efficacy of Pallidal or Subthalamic Nucleus Stimulation in Advanced PD. Neurology 2001, 56, 548–551. [Google Scholar] [CrossRef]
- Limousin, P.; Foltynie, T. Long-Term Outcomes of Deep Brain Stimulation in Parkinson Disease. Nat. Rev. Neurol. 2019, 15, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Guimarães Rocha, M.S.; Letícia de Freitas, J.; Torres, I.A.; Alves Matos, P.C.; Terzian, P.R.; Santos Tatsch, J.F.; Godinho, F. Subthalamic Nucleus Deep Brain Stimulation Lessens Acquired Dystonia: Report of Two Patients and Systematic Review of Published Cases. Stereotact. Funct. Neurosurg. 2021, 99, 412–424. [Google Scholar] [CrossRef]
- Ostrem, J.L.; Racine, C.A.; Glass, G.A.; Grace, J.K.; Volz, M.M.; Heath, S.L.; Starr, P.A. Subthalamic Nucleus Deep Brain Stimulation in Primary Cervical Dystonia. Neurology 2011, 76, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Handforth, A.; DeSalles, A.A.F.; Krahl, S.E. Deep Brain Stimulation of the Subthalamic Nucleus as Adjunct Treatment for Refractory Epilepsy. Epilepsia 2006, 47, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Yu, T.; Wang, D.; Wang, X.; Ni, D.; Zhang, G.; Bartolomei, F.; Wang, Y.; Li, Y. Subthalamic Nucleus Stimulation Modulates Motor Epileptic Activity in Humans. Ann. Neurol. 2020, 88, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Iorio-Morin, C.; Fomenko, A.; Kalia, S.K. Deep-Brain Stimulation for Essential Tremor and Other Tremor Syndromes: A Narrative Review of Current Targets and Clinical Outcomes. Brain Sci. 2020, 10, 925. [Google Scholar] [CrossRef] [PubMed]
- Mallet, L.; Polosan, M.; Jaafari, N.; Baup, N.; Welter, M.-L.; Fontaine, D.; du Montcel, S.T.; Yelnik, J.; Chéreau, I.; Arbus, C.; et al. Subthalamic Nucleus Stimulation in Severe Obsessive-Compulsive Disorder. N. Engl. J. Med. 2008, 359, 2121–2134. [Google Scholar] [CrossRef] [Green Version]
- Pelloux, Y.; Baunez, C. Deep Brain Stimulation for Addiction: Why the Subthalamic Nucleus Should Be Favored. Curr. Opin. Neurobiol. 2013, 23, 713–720. [Google Scholar] [CrossRef]
- Hamani, C. The Subthalamic Nucleus in the Context of Movement Disorders. Brain 2004, 127, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Rojas, R.; Pineda-Pardo, J.A.; Mañez-Miro, J.; Sanchez-Turel, A.; Martinez-Fernandez, R.; del Alamo, M.; DeLong, M.; Obeso, J.A. Functional Topography of the Human Subthalamic Nucleus: Relevance for Subthalamotomy in Parkinson’s Disease. Mov. Disord. 2022, 37, 279–290. [Google Scholar] [CrossRef]
- Nambu, A.; Takada, M.; Inase, M.; Tokuno, H. Dual Somatotopical Representations in the Primate Subthalamic Nucleus: Evidence for Ordered but Reversed Body-Map Transformations from the Primary Motor Cortex and the Supplementary Motor Area. J. Neurosci. 1996, 16, 2671–2683. [Google Scholar] [CrossRef]
- Obeso, J.A.; Rodríguez-Oroz, M.C.; Benitez-Temino, B.; Blesa, F.J.; Guridi, J.; Marin, C.; Rodriguez, M. Functional Organization of the Basal Ganglia: Therapeutic Implications for Parkinson’s Disease. Mov. Disord. 2008, 23, S548–S559. [Google Scholar] [CrossRef]
- Smith, Y.; Bolam, J.P.; Von Krosigk, M. Topographical and Synaptic Organization of the GABA-Containing Pallidosubthalamic Projection in the Rat. Eur. J. Neurosci. 1990, 2, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Marani, E. (Ed.) The Subthalamic Nucleus: Development, Cytology, Topography and Connections; Advances in Anatomy, Embryology, and Cell Biology; Springer: Berlin, Germany, 2008; ISBN 978-3-540-79459-2. [Google Scholar]
- Anderson, R.W.; Farokhniaee, A.; Gunalan, K.; Howell, B.; McIntyre, C.C. Action Potential Initiation, Propagation, and Cortical Invasion in the Hyperdirect Pathway during Subthalamic Deep Brain Stimulation. Brain Stimulat. 2018, 11, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Kita, T.; Kita, H. The Subthalamic Nucleus Is One of Multiple Innervation Sites for Long-Range Corticofugal Axons: A Single-Axon Tracing Study in the Rat. J. Neurosci. 2012, 32, 5990–5999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, A.; Crossman, A.R. Subthalamic Nucleus Efferent Projection to the Cerebral Cortex. Neuroscience 1981, 6, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Kita, H.; Kitai, S.T. Efferent Projections of the Subthalamic Nucleus in the Rat: Light and Electron Microscopic Analysis with the PHA-L Method. J. Comp. Neurol. 1987, 260, 435–452. [Google Scholar] [CrossRef]
- Crossman, A.R. Primate Models of Dyskinesia: The Experimental Approach to the Study of Basal Ganglia-Related Involuntary Movement Disorders. Neuroscience 1987, 21, 1–40. [Google Scholar] [CrossRef]
- DeLong, M.R. Primate Models of Movement Disorders of Basal Ganglia Origin. Trends Neurosci. 1990, 13, 281–285. [Google Scholar] [CrossRef]
- Guridi, J.; Obeso, J.A. The Subthalamic Nucleus, Hemiballismus and Parkinson’s Disease: Reappraisal of a Neurosurgical Dogma. Brain 2001, 124, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Krack, P.; Volkmann, J.; Tinkhauser, G.; Deuschl, G. Deep Brain Stimulation in Movement Disorders: From Experimental Surgery to Evidence-Based Therapy. Mov. Disord. 2019, 34, 1795–1810. [Google Scholar] [CrossRef]
- Klein, J.; Soto-Montenegro, M.L.; Pascau, J.; Günther, L.; Kupsch, A.; Desco, M.; Winter, C. A Novel Approach to Investigate Neuronal Network Activity Patterns Affected by Deep Brain Stimulation in Rats. J. Psychiatr. Res. 2011, 45, 927–930. [Google Scholar] [CrossRef]
- Liang, S.; Jiang, X.; Zhang, Q.; Duan, S.; Zhang, T.; Huang, Q.; Sun, X.; Liu, H.; Dong, J.; Liu, W.; et al. Abnormal Metabolic Connectivity in Rats at the Acute Stage of Ischemic Stroke. Neurosci. Bull. 2018, 34, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Krämer, S.D.; Schuhmann, M.K.; Schadt, F.; Israel, I.; Samnick, S.; Volkmann, J.; Fluri, F. Changes of Cerebral Network Activity after Invasive Stimulation of the Mesencephalic Locomotor Region in a Rat Stroke Model. Exp. Neurol. 2021, 113884. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Zhang, H.; Li, D.; Zhan, S.; Zhang, J.; Zhang, X.; Zuo, C.; Sun, B. Modified Fluorodeoxyglucose Metabolism in Motor Circuitry by Subthalamic Deep Brain Stimulation. Stereotact. Funct. Neurosurg. 2017, 95, 93–101. [Google Scholar] [CrossRef]
- Fukuda, M.; Mentis, M.J.; Ma, Y.; Dhawan, V.; Antonini, A.; Lang, A.E.; Lozano, A.M.; Hammerstad, J.; Lyons, K.; Koller, W.C.; et al. Networks Mediating the Clinical Effects of Pallidal Brain Stimulation for Parkinson’s DiseaseA PET Study of Resting-State Glucose Metabolism. Brain 2001, 124, 1601–1609. [Google Scholar] [CrossRef]
- Gao, F.; Guo, Y.; Zhang, H.; Wang, S.; Wang, J.; Wu, J.-M.; Chen, Z.; Ding, M.-P. Anterior Thalamic Nucleus Stimulation Modulates Regional Cerebral Metabolism: An FDG-MicroPET Study in Rats. Neurobiol. Dis. 2009, 34, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Mattis, P.; Tang, C.; Perrine, K.; Carbon, M.; Eidelberg, D. Metabolic Brain Networks Associated with Cognitive Function in Parkinson’s Disease. NeuroImage 2007, 34, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Whishaw, I.Q.; Pellis, S.M. The Structure of Skilled Forelimb Reaching in the Rat: A Proximally Driven Movement with a Single Distal Rotatory Component. Behav. Brain Res. 1990, 41, 49–59. [Google Scholar] [CrossRef]
- Hammond, C.; Rouzaire-Dubois, B.; Féger, J.; Jackson, A.; Crossman, A.R. Anatomical and Electrophysiological Studies on the Reciprocal Projections between the Subthalamic Nucleus and Nucleus Tegmenti Pedunculopontinus in the Rat. Neuroscience 1983, 9, 41–52. [Google Scholar] [CrossRef]
- Degos, B.; Deniau, J.-M.; Le Cam, J.; Mailly, P.; Maurice, N. Evidence for a Direct Subthalamo-Cortical Loop Circuit in the Rat. Eur. J. Neurosci. 2008, 27, 2599–2610. [Google Scholar] [CrossRef]
- Takada, M.; Li, Z.K.; Hattori, T. Long Descending Direct Projection from the Basal Ganglia to the Spinal Cord: A Revival of the Extrapyramidal Concept. Brain Res. 1987, 436, 129–135. [Google Scholar] [CrossRef]
- Kitai, S.T.; Deniau, J.M. Cortical Inputs to the Subthalamus: Intracellular Analysis. Brain Res. 1981, 214, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Canteras, N.S.; Shammah-Lagnado, S.J.; Silva, B.A.; Ricardo, J.A. Somatosensory Inputs to the Subthalamic Nucleus: A Combined Retrograde and Anterograde Horseradish Peroxidase Study in the Rat. Brain Res. 1988, 458, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, T.; Hattori, T. Confirmation of Thalamosubthalamic Projections by Electron Microscopic Autoradiography. Brain Res. 1983, 267, 335–339. [Google Scholar] [CrossRef]
- Saper, C.B.; Swanson, L.W.; Cowan, W.M. The Efferent Connections of the Ventromedial Nucleus of the Hypothalamus of the Rat. J. Comp. Neurol. 1976, 169, 409–442. [Google Scholar] [CrossRef] [PubMed]
- Canteras, N.S.; Shammah-Lagnado, S.J.; Silva, B.A.; Ricardo, J.A. Afferent Connections of the Subthalamic Nucleus: A Combined Retrograde and Anterograde Horseradish Peroxidase Study in the Rat. Brain Res. 1990, 513, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Cavdar, S.; Özgür, M.; Çakmak, Y.Ö.; Kuvvet, Y.; Kunt, S.K.; Sağlam, G. Afferent Projections of the Subthalamic Nucleus in the Rat: Emphasis on Bilateral and Interhemispheric Connections. Acta Neurobiol. Exp. 2018, 78, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Dromerick, A.W.; Lang, C.E.; Birkenmeier, R.; Hahn, M.G.; Sahrmann, S.A.; Edwards, D.F. Relationships between Upper-Limb Functional Limitation and Self-Reported Disability 3 Months after Stroke. J. Rehabil. Res. Dev. 2006, 43, 401–408. [Google Scholar] [CrossRef]
- Fluri, F.; Malzahn, U.; Homola, G.A.; Schuhmann, M.K.; Kleinschnitz, C.; Volkmann, J. Stimulation of the Mesencephalic Locomotor Region for Gait Recovery after Stroke: DBS for Gait Recovery. Ann. Neurol. 2017, 82, 828–840. [Google Scholar] [CrossRef]
- Machado, A.G.; Cooperrider, J.; Furmaga, H.T.; Baker, K.B.; Park, H.-J.; Chen, Z.; Gale, J.T. Chronic 30-Hz Deep Cerebellar Stimulation Coupled with Training Enhances Post-Ischemia Motor Recovery and Peri-Infarct Synaptophysin Expression in Rodents. Neurosurgery 2013, 73, 344–353, discussion 353. [Google Scholar] [CrossRef]
- Khanna, P.; Totten, D.; Novik, L.; Roberts, J.; Morecraft, R.J.; Ganguly, K. Low-Frequency Stimulation Enhances Ensemble Co-Firing and Dexterity after Stroke. Cell 2021, 184, 912–930.e20. [Google Scholar] [CrossRef]
- Gharbawie, O.A.; Karl, J.M.; Whishaw, I.Q. Recovery of Skilled Reaching Following Motor Cortex Stroke: Do Residual Corticofugal Fibers Mediate Compensatory Recovery? Eur. J. Neurosci. 2007, 26, 3309–3327. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.-K.; Alaverdashvili, M.; Cross, A.R.; Whishaw, I.Q. Both Compensation and Recovery of Skilled Reaching Following Small Photothrombotic Stroke to Motor Cortex in the Rat. Exp. Neurol. 2009, 218, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Jeffers, M.S.; Touvykine, B.; Ripley, A.; Lahey, G.; Carter, A.; Dancause, N.; Corbett, D. Poststroke Impairment and Recovery Are Predicted by Task-Specific Regionalization of Injury. J. Neurosci. 2020, 40, 6082–6097. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Sugiyama, K.; Akamine, S.; Sun, W.; Namba, H. The Different Performance among Motor Tasks during the Increasing Current Intensity of Deep Brain Stimulation of the Subthalamic Nucleus in Rats with Different Degrees of the Unilateral Striatal Lesion. Neurosci. Lett. 2010, 480, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Chassain, C.; Melon, C.; Salin, P.; Vitale, F.; Couraud, S.; Durif, F.; Kerkerian-Le Goff, L.; Gubellini, P. Metabolic, Synaptic and Behavioral Impact of 5-Week Chronic Deep Brain Stimulation in Hemiparkinsonian Rats. J. Neurochem. 2016, 136, 1004–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musacchio, T.; Rebenstorff, M.; Fluri, F.; Brotchie, J.M.; Volkmann, J.; Koprich, J.B.; Ip, C.W. Subthalamic Nucleus Deep Brain Stimulation Is Neuroprotective in the A53T α-Synuclein Parkinson’s Disease Rat Model. Ann. Neurol. 2017, 81, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Spieles-Engemann, A.L.; Steece-Collier, K.; Behbehani, M.M.; Collier, T.J.; Wohlgenant, S.L.; Kemp, C.J.; Cole-Strauss, A.; Levine, N.D.; Gombash, S.E.; Thompson, V.B.; et al. Subthalamic Nucleus Stimulation Increases Brain Derived Neurotrophic Factor in the Nigrostriatal System and Primary Motor Cortex. J. Park. Dis. 2011, 1, 123–136. [Google Scholar] [CrossRef]
- Faust, K.; Vajkoczy, P.; Xi, B.; Harnack, D. The Effects of Deep Brain Stimulation of the Subthalamic Nucleus on Vascular Endothelial Growth Factor, Brain-Derived Neurotrophic Factor, and Glial Cell Line-Derived Neurotrophic Factor in a Rat Model of Parkinson’s Disease. Stereotact. Funct. Neurosurg. 2021, 99, 256–266. [Google Scholar] [CrossRef]
- Cassar, I.R.; Grill, W.M. The Cortical Evoked Potential Corresponds with Deep Brain Stimulation Efficacy in Rats. J. Neurophysiol. 2022, 127, 1253–1268. [Google Scholar] [CrossRef]
- Li, Q.; Ke, Y.; Chan, D.C.W.; Qian, Z.-M.; Yung, K.K.L.; Ko, H.; Arbuthnott, G.W.; Yung, W.-H. Therapeutic Deep Brain Stimulation in Parkinsonian Rats Directly Influences Motor Cortex. Neuron 2012, 76, 1030–1041. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Qian, Z.-M.; Arbuthnott, G.W.; Ke, Y.; Yung, W.-H. Cortical Effects of Deep Brain Stimulation: Implications for Pathogenesis and Treatment of Parkinson Disease. JAMA Neurol. 2014, 71, 100. [Google Scholar] [CrossRef]
- Kumaravelu, K.; Oza, C.S.; Behrend, C.E.; Grill, W.M. Model-Based Deconstruction of Cortical Evoked Potentials Generated by Subthalamic Nucleus Deep Brain Stimulation. J. Neurophysiol. 2018, 120, 662–680. [Google Scholar] [CrossRef] [Green Version]
- Nudo, R.J.; Milliken, G.W. Reorganization of Movement Representations in Primary Motor Cortex Following Focal Ischemic Infarcts in Adult Squirrel Monkeys. J. Neurophysiol. 1996, 75, 2144–2149. [Google Scholar] [CrossRef]
- Gharbawie, O.A.; Gonzalez, C.L.R.; Whishaw, I.Q. Skilled Reaching Impairments from the Lateral Frontal Cortex Component of Middle Cerebral Artery Stroke: A Qualitative and Quantitative Comparison to Focal Motor Cortex Lesions in Rats. Behav. Brain Res. 2005, 156, 125–137. [Google Scholar] [CrossRef]
- Latifi, S.; Mitchell, S.; Habibey, R.; Hosseini, F.; Donzis, E.; Estrada-Sánchez, A.M.; Nejad, H.R.; Levine, M.; Golshani, P.; Carmichael, S.T. Neuronal Network Topology Indicates Distinct Recovery Processes after Stroke. Cereb. Cortex 2020, 30, 6363–6375. [Google Scholar] [CrossRef]
- Balbi, M.; Xiao, D.; Jativa Vega, M.; Hu, H.; Vanni, M.P.; Bernier, L.-P.; LeDue, J.; MacVicar, B.; Murphy, T.H. Gamma Frequency Activation of Inhibitory Neurons in the Acute Phase after Stroke Attenuates Vascular and Behavioral Dysfunction. Cell Rep. 2021, 34, 108696. [Google Scholar] [CrossRef]
- Guo, L.; Kondapavulur, S.; Lemke, S.M.; Won, S.J.; Ganguly, K. Coordinated Increase of Reliable Cortical and Striatal Ensemble Activations during Recovery after Stroke. Cell Rep. 2021, 36, 109370. [Google Scholar] [CrossRef]
- Ruder, L.; Schina, R.; Kanodia, H.; Valencia-Garcia, S.; Pivetta, C.; Arber, S. A Functional Map for Diverse Forelimb Actions within Brainstem Circuitry. Nature 2021, 590, 445–450. [Google Scholar] [CrossRef]
- Wolff, S.B.E.; Ko, R.; Ölveczky, B.P. Distinct Roles for Motor Cortical and Thalamic Inputs to Striatum during Motor Skill Learning and Execution. Sci. Adv. 2022, 8, eabk0231. [Google Scholar] [CrossRef]
- Cramer, S.C. Repairing the Human Brain after Stroke: I. Mechanisms of Spontaneous Recovery. Ann. Neurol. 2008, 63, 272–287. [Google Scholar] [CrossRef]
- Grefkes, C.; Ward, N.S. Cortical Reorganization After Stroke How Much and How Functional? Neuroscientist 2014, 20, 56–70. [Google Scholar] [CrossRef]
- Rüber, T.; Schlaug, G.; Lindenberg, R. Compensatory Role of the Cortico-Rubro-Spinal Tract in Motor Recovery after Stroke. Neurology 2012, 79, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Dudman, J.T.; Krakauer, J.W. The Basal Ganglia: From Motor Commands to the Control of Vigor. Curr. Opin. Neurobiol. 2016, 37, 158–166. [Google Scholar] [CrossRef]
- Fobbs, W.C.; Bariselli, S.; Licholai, J.A.; Miyazaki, N.L.; Matikainen-Ankney, B.A.; Creed, M.C.; Kravitz, A.V. Continuous Representations of Speed by Striatal Medium Spiny Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2020, 40, 1679–1688. [Google Scholar] [CrossRef]
- Yin, H.H.; Mulcare, S.P.; Hilário, M.R.F.; Clouse, E.; Holloway, T.; Davis, M.I.; Hansson, A.C.; Lovinger, D.M.; Costa, R.M. Dynamic Reorganization of Striatal Circuits during the Acquisition and Consolidation of a Skill. Nat. Neurosci. 2009, 12, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Lemke, S.M.; Ramanathan, D.S.; Guo, L.; Won, S.J.; Ganguly, K. Emergent Modular Neural Control Drives Coordinated Motor Actions. Nat. Neurosci. 2019, 22, 1122–1131. [Google Scholar] [CrossRef]
- Florio, T.; Scarnati, E.; Confalone, G.; Minchella, D.; Galati, S.; Stanzione, P.; Stefani, A.; Mazzone, P. High-Frequency Stimulation of the Subthalamic Nucleus Modulates the Activity of Pedunculopontine Neurons through Direct Activation of Excitatory Fibres as Well as through Indirect Activation of Inhibitory Pallidal Fibres in the Rat. Eur. J. Neurosci. 2007, 25, 1174–1186. [Google Scholar] [CrossRef]
- Svoboda, K.; Li, N. Neural Mechanisms of Movement Planning: Motor Cortex and Beyond. Curr. Opin. Neurobiol. 2018, 49, 33–41. [Google Scholar] [CrossRef]
- Arber, S.; Costa, R.M. Networking Brainstem and Basal Ganglia Circuits for Movement. Nat. Rev. Neurosci. 2022, 23, 342–360. [Google Scholar] [CrossRef]
- Esposito, M.S.; Capelli, P.; Arber, S. Brainstem Nucleus MdV Mediates Skilled Forelimb Motor Tasks. Nature 2014, 508, 351–356. [Google Scholar] [CrossRef]
- Donegan, D.; Kanzler, C.M.; Büscher, J.; Viskaitis, P.; Bracey, E.F.; Lambercy, O.; Burdakov, D. Hypothalamic Control of Forelimb Motor Adaptation. J. Neurosci. 2022, 42, 6243–6257. [Google Scholar] [CrossRef]
- Lindau, N.T.; Bänninger, B.J.; Gullo, M.; Good, N.A.; Bachmann, L.C.; Starkey, M.L.; Schwab, M.E. Rewiring of the Corticospinal Tract in the Adult Rat after Unilateral Stroke and Anti-Nogo-A Therapy. Brain 2013, awt336. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Fluri, F.; Bieber, M.; Volkmann, J.; Kleinschnitz, C. Microelectrode Guided Implantation of Electrodes into the Subthalamic Nucleus of Rats for Long-Term Deep Brain Stimulation. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [Green Version]
- Maesawa, S.; Kaneoke, Y.; Kajita, Y.; Usui, N.; Misawa, N.; Nakayama, A.; Yoshida, J. Long-Term Stimulation of the Subthalamic Nucleus in Hemiparkinsonian Rats: Neuroprotection of Dopaminergic Neurons. J. Neurosurg. 2004, 100, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Lapa, C.; Lückerath, K.; Malzahn, U.; Samnick, S.; Einsele, H.; Buck, A.K.; Herrmann, K.; Knop, S. 18 FDG-PET/CT for Prognostic Stratification of Patients with Multiple Myeloma Relapse after Stem Cell Transplantation. Oncotarget 2014, 5, 7381–7391. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krämer, S.D.; Schuhmann, M.K.; Volkmann, J.; Fluri, F. Deep Brain Stimulation in the Subthalamic Nucleus Can Improve Skilled Forelimb Movements and Retune Dynamics of Striatal Networks in a Rat Stroke Model. Int. J. Mol. Sci. 2022, 23, 15862. https://doi.org/10.3390/ijms232415862
Krämer SD, Schuhmann MK, Volkmann J, Fluri F. Deep Brain Stimulation in the Subthalamic Nucleus Can Improve Skilled Forelimb Movements and Retune Dynamics of Striatal Networks in a Rat Stroke Model. International Journal of Molecular Sciences. 2022; 23(24):15862. https://doi.org/10.3390/ijms232415862
Chicago/Turabian StyleKrämer, Stefanie D., Michael K. Schuhmann, Jens Volkmann, and Felix Fluri. 2022. "Deep Brain Stimulation in the Subthalamic Nucleus Can Improve Skilled Forelimb Movements and Retune Dynamics of Striatal Networks in a Rat Stroke Model" International Journal of Molecular Sciences 23, no. 24: 15862. https://doi.org/10.3390/ijms232415862