
Anti-Inflammatory Mechanisms of Dietary Flavones: Tapping into Nature to Control Chronic Inflammation in Obesity and Cancer

Abstract
:1. Introduction
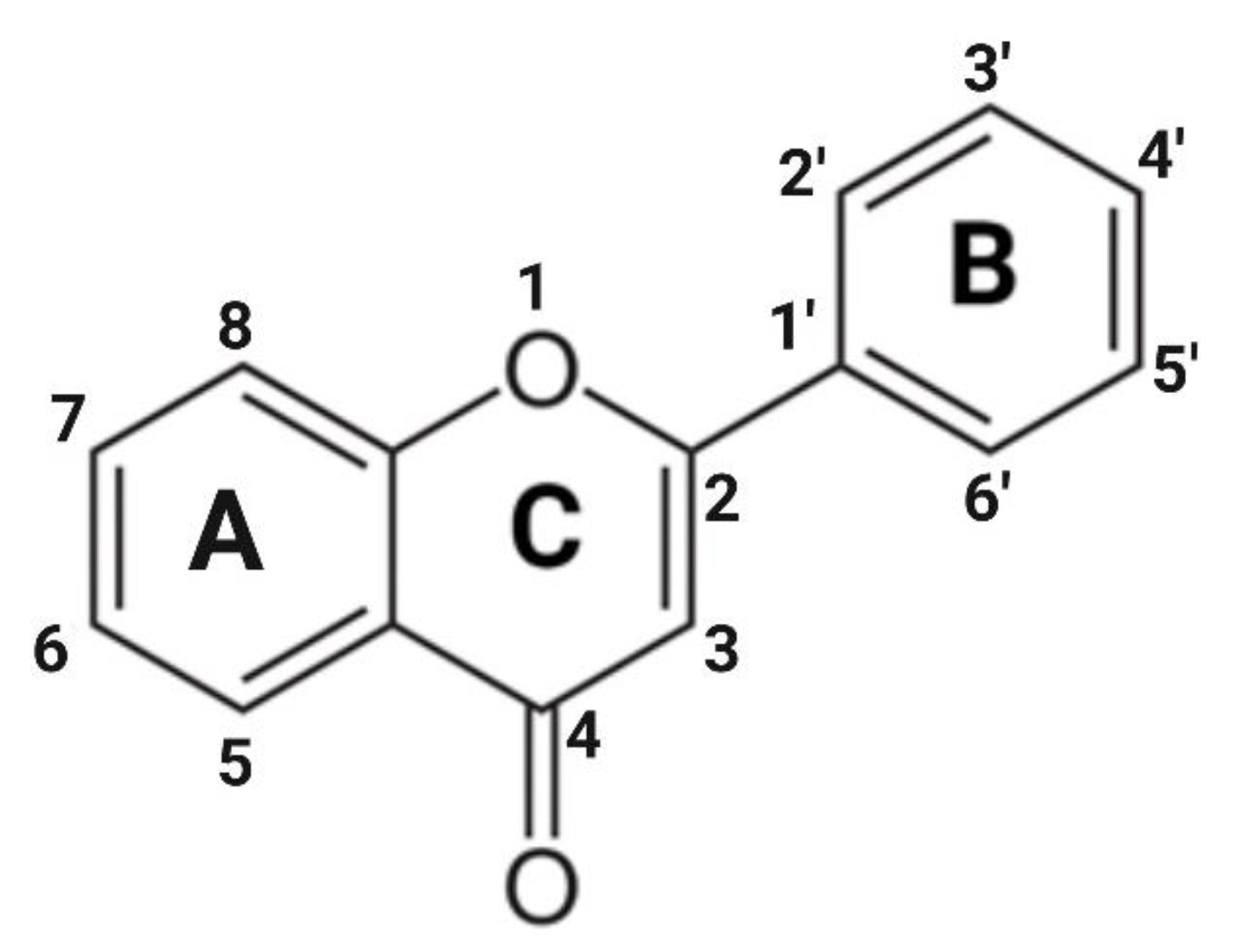
2. Flavones: Structural Characteristics and Distribution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavone | Hydroxyl Position | Modifications | Source | Ref. |
|---|---|---|---|---|
| Chrysin | 5, 7 | - | Bitter melon, wild Himalayan pear, honey, propolis, passion flowers, oyster mushroom | [25] |
| Apigenin | 5, 7, 4′ | - | Parsley, celery, onions, chamomile, thyme, basil, oregano, artichoke | [22,29] |
| Apigetrin | 5, 7, 4′ | 7-O-glycoside | Dandelion | [45] |
| Acacetin | 5, 7, 4′ | 4′- OCH3 | Black locust, silver birch | [46] |
| Vitexin | 5, 7, 4′ | 8-C-glycoside | Passion flower, chasteberry, bamboo, Hawthorn, fenugreek | [19,47] |
| Baicalein | 5, 6, 7 | - | Thyme, Scutellaria baicalensis | [26] |
| Baicalin | 5, 6, 7 | 7-O-glycoside | Scutellaria baicalensis, Oroxylum indicum | [26] |
| Wogonin | 5, 7, 8 | 8-OCH3 | Scutellaria baicalensis | [26] |
| Luteolin | 5, 7, 3′, 4′ | - | Celery, broccoli, bell pepper, parsley, thyme, carrots, rosemary, chamomile, oregano, green onions, fennel, sage | [22,48] |
| Diosmetin | 5, 7, 3′, 4′ | 4′-OCH3 | Caucasian vetch, citrus fruit | [48] |
| Diosmin | 5, 7, 3′, 4′ | 7-O-rutinoside, 4′-OCH3 | Citrus fruit | [48] |
| Cynaroside | 5, 7, 3′, 4′ | 7-O-glycoside | Cumin | [49] |
| Orientin | 5, 7, 3′, 4′ | 8-C-glycoside | Passion flower, buckwheat sprouts, millets | [50] |
| Isoorientin | 5, 7, 3′, 4′ | 6-C-glycoside | Passion flower, Acai palm | [50,51] |
| Scutellarein | 5, 6, 7, 4′ | - | Scutellaria baicalensis | [26] |
| Scutellarin | 5, 6, 7, 4′ | 7-O-glucurinide | Scutellaria baicalensis | [26] |
| Tangeretin | 5, 6, 7, 8, 4′ | 4′-OCH3 | Citrus fruit | [48] |
| Nobiletin | 5, 6, 7, 8, 3′, 4′ | 4′-OCH3 | Citrus fruit | [48] |
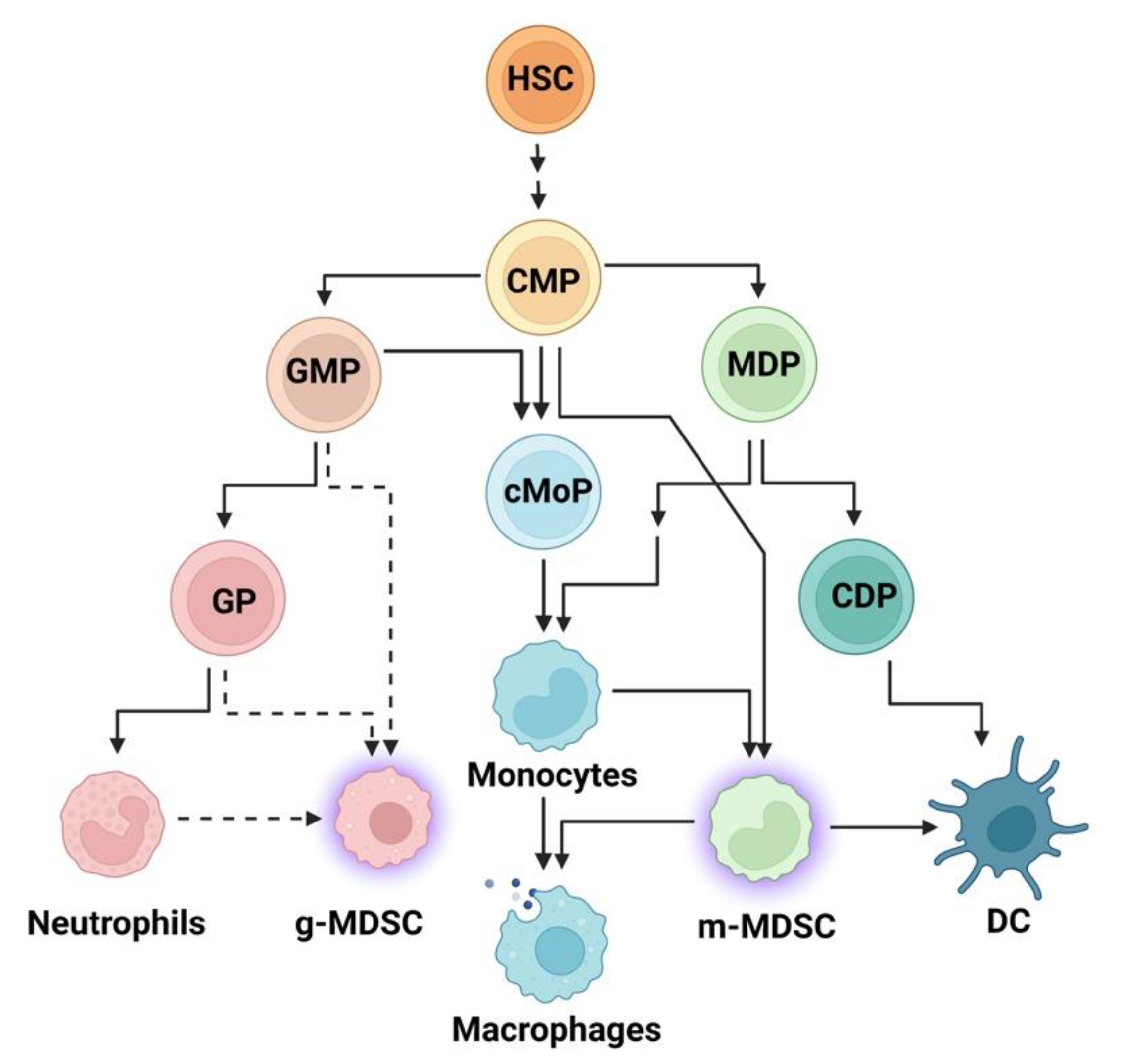
3. The Innate Immune System and Inflammation
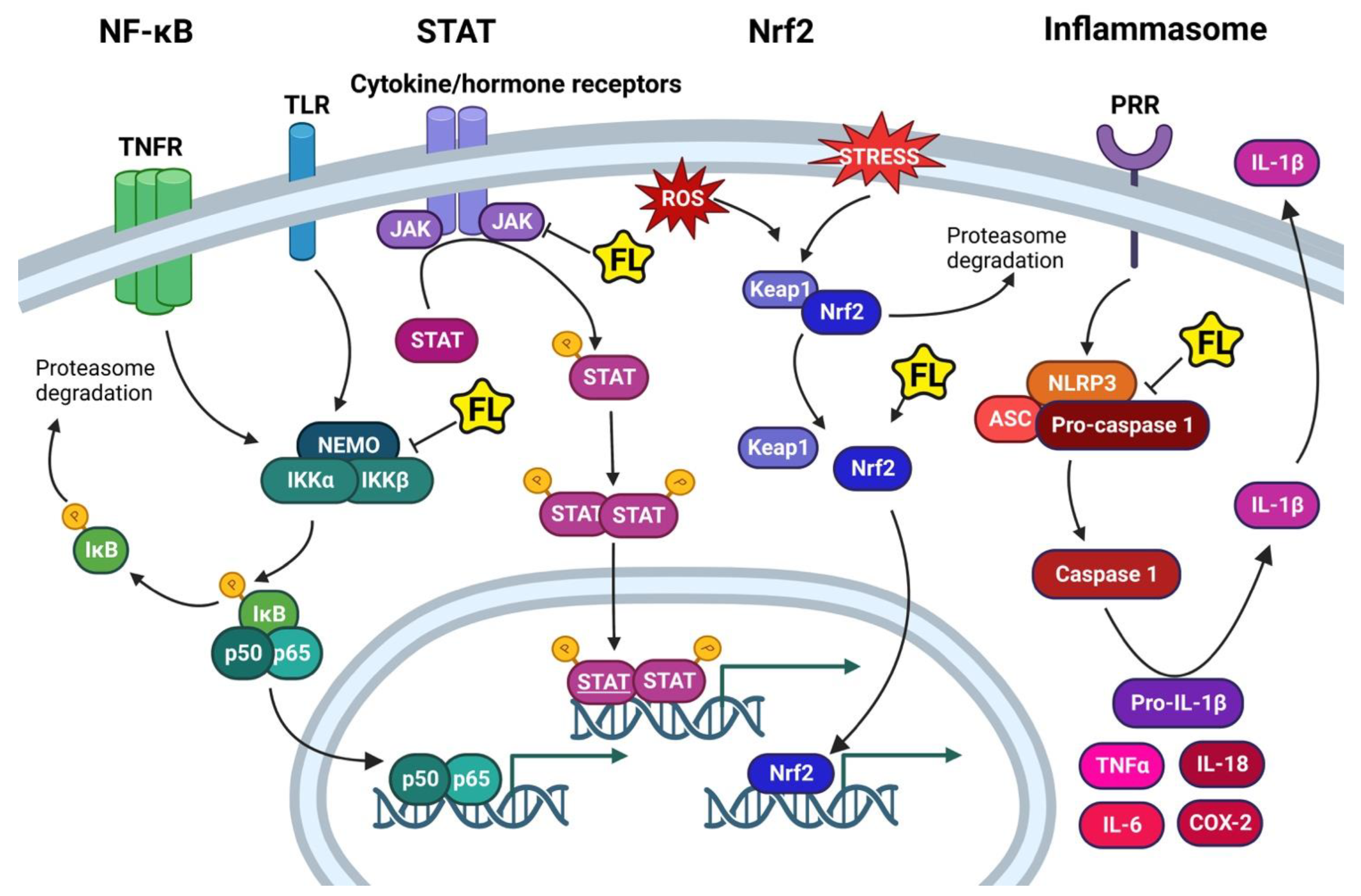
4. Inflammatory Pathways
4.1. NF-κB
4.2. STAT Pathway
4.3. Nrf2 Pathway
4.4. The Inflammasome Pathway
5. Chronic Inflammatory Diseases: Obesity and Cancer
5.1. Obesity-Induced Inflammation
5.2. Cancer-Induced Dysregulation of Innate Immunity
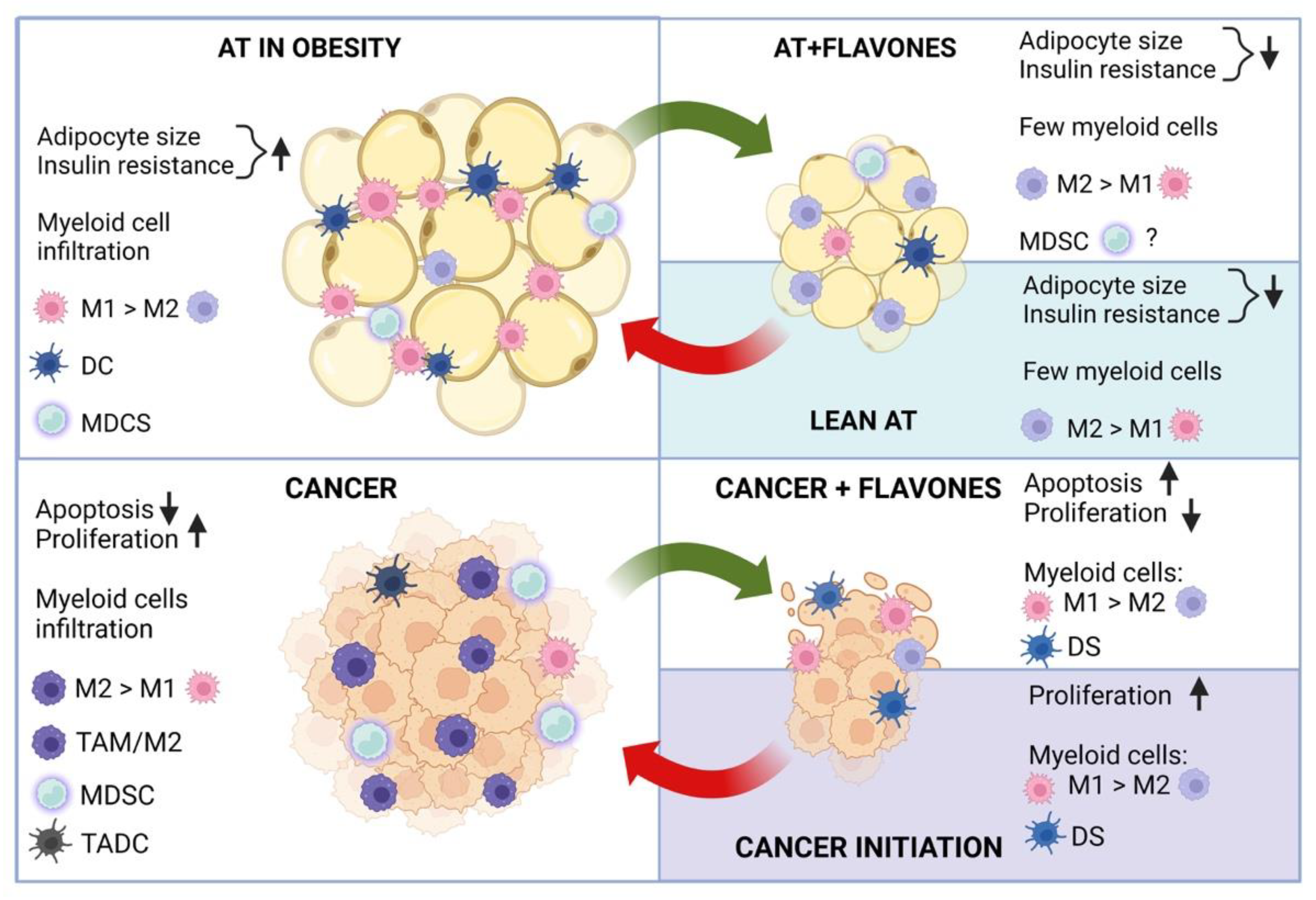
6. Anti-Inflammatory Mechanisms of Flavones
6.1. Flavones and Innate Immune Cell Repertoires in Obesity
6.2. Flavones Control Innate Immune Cell Repertoires and Inflammation in Cancer
7. Conclusions and Future Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pahwa, R.; Goyal, A.; Jialal, I. Chronic Inflammation; StatPearl Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Raghupathi, W.; Raghupathi, V. An Empirical Study of Chronic Diseases in the United States: A Visual Analytics Approach. Int. J. Environ. Res. Public Health 2018, 15, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, D.P.; Vona-Davis, L. Influence of Obesity on Breast Cancer Receptor Status and Prognosis. Expert Rev. Anticancer Ther. 2009, 9, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Bandera, E.V.; Maskarinec, G.; Romieu, I.; John, E.M. Racial and Ethnic Disparities in the Impact of Obesity on Breast Cancer Risk and Survival: A global perspective. Adv. Nutr. 2015, 6, 803–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yedjou, C.G.; Sims, J.N.; Miele, L.; Noubissi, F.; Lowe, L.; Fonseca, D.D.; Alo, R.A.; Payton, M.; Tchounwou, P.B. Health and Racial Disparity in Breast Cancer. Adv. Exp. Med. Biol. 2019, 1152, 31–49. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Jemal, A.; Wender, R.C.; Gansler, T.; Ma, J.; Brawley, O.W. An Assessment of Progress in Cancer Control. CA Cancer J. Clin. 2018, 68, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.C.; Kunnumakkara, A.B.; Aggarwal, S.; Aggarwal, B.B. Inflammation, a Double-Edge Sword for Cancer and Other Age-Related Diseases. Front. Immunol. 2018, 9, 2160. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose Tissue Inflammation and Metabolic Dysfunction in Obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef]
- Hou, J.; Karin, M.; Sun, B. Targeting Cancer-Promoting Inflammation—Have Anti-Inflammatory Therapies Come of Age? Nat. Rev. Clin. Oncol. 2021, 18, 261–279. [Google Scholar] [CrossRef]
- Veglia, F.; Gabrilovich, D.I. Dendritic Cells in Cancer: The Role Revisited. Curr. Opin. Immunol. 2017, 45, 43–51. [Google Scholar] [CrossRef]
- Sanchez-Pino, M.D.; Gilmore, L.A.; Ochoa, A.C.; Brown, J.C. Obesity-Associated Myeloid Immunosuppressive Cells, Key Players in Cancer Risk and Response to Immunotherapy. Obesity 2021, 29, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Soedono, S.; Cho, K.W. Adipose Tissue Dendritic Cells: Critical Regulators of Obesity-Induced Inflammation and Insulin Resistance. Int. J. Mol. Sci. 2021, 22, 8666. [Google Scholar] [CrossRef] [PubMed]
- Zappavigna, S.; Cossu, A.M.; Grimaldi, A.; Bocchetti, M.; Ferraro, G.A.; Nicoletti, G.F.; Filosa, R.; Caraglia, M. Anti-Inflammatory Drugs as Anticancer Agents. Int. J. Mol. Sci. 2020, 21, 2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) and Organ Damage: A Current Perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-Inflammatory Effects of Flavonoids. Food. Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Butt, G.; El-Zahaby, S.A.; Attar, R.; Sabitaliyevich, U.Y.; Jovic, J.J.; Tang, K.F.; Naureen, H.; Xu, B. Luteolin Mediated Targeting of Protein Network and Micrornas in Different Cancers: Focus on JAK-STAT, NOTCH, mTOR and TRAIL-Mediated Signaling Pathways. Pharmacol. Res. 2020, 160, 105188. [Google Scholar] [CrossRef]
- Gendrisch, F.; Esser, P.R.; Schempp, C.M.; Wölfle, U. Luteolin as a Modulator of Skin Aging and Inflammation. Biofactors 2021, 47, 170–180. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Parama, D.; Daimari, E.; Girisa, S.; Banik, K.; Harsha, C.; Dutta, U.; Kunnumakkara, A.B. Rationalizing the Therapeutic Potential of Apigenin against Cancer. Life Sci. 2021, 267, 118814. [Google Scholar] [CrossRef]
- Abid, R.; Ghazanfar, S.; Farid, A.; Sulaman, S.M.; Idrees, M.; Amen, R.A.; Muzammal, M.; Shahzad, M.K.; Mohamed, M.O.; Khaled, A.A.; et al. Pharmacological Properties of 4′, 5, 7-Trihydroxyflavone (Apigenin) and Its Impact on Cell Signaling Pathways. Molecules 2022, 27, 4304. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponte, L.G.S.; Pavan, I.C.B.; Mancini, M.C.S.; da Silva, L.G.S.; Morelli, A.P.; Severino, M.B.; Bezerra, R.M.N.; Simabuco, F.M. The Hallmarks of Flavonoids in Cancer. Molecules 2021, 26, 2029. [Google Scholar] [CrossRef] [PubMed]
- Woo, E.R.; Pokharel, Y.R.; Yang, J.W.; Lee, S.Y.; Kang, K.W. Inhibition of Nuclear Factor-KappaB Activation By 2′,8″-Biapigenin. Biol. Pharm. Bull. 2006, 29, 976–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stompor-Gorący, M.; Bajek-Bil, A.; Machaczka, M. Chrysin: Perspectives on Contemporary Status and Future Possibilities as Pro-Health Agent. Nutrients 2021, 13, 2038. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A Review of Its Traditional Uses, Botany, Phytochemistry, Pharmacology and Toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, G.; Riedl, K.; Cardenas, H.; Diosa-Toro, M.; Arango, D.; Schwartz, S.; Doseff, A.I. Flavone Deglycosylation Increases Their Anti-Inflammatory Activity and Absorption. Mol. Nutr. Food Res. 2012, 56, 558–569. [Google Scholar] [CrossRef]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy Isoflavone Aglycones Are Absorbed Faster and in Higher Amounts Than Their Glucosides in Humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Borges, G.; Fong, R.Y.; Ensunsa, J.L.; Kimball, J.; Medici, V.; Ottaviani, J.I.; Crozier, A. Absorption, Distribution, Metabolism and Excretion of Apigenin and Its Glycosides in Healthy Male Adults. Free Radic. Biol. Med. 2022, 185, 90–96. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Luján-Barroso, L.; Slimani, N.; Romieu, I.; Fedirko, V.; de Magistris, M.S.; Ericson, U.; Amiano, P.; Trichopoulou, A.; et al. Estimated Dietary Intakes Of Flavonols, Flavanones and Flavones in The European Prospective Investigation into Cancer and Nutrition (EPIC) 24 Hour Dietary Recall Cohort. Br. J. Nutr. 2011, 106, 1915–1925. [Google Scholar] [CrossRef] [Green Version]
- Jennings, A.; Welch, A.A.; Spector, T.; Macgregor, A.; Cassidy, A. Intakes of Anthocyanins and Flavones Are Associated with Biomarkers of Insulin Resistance and Inflammation in Women. J. Nutr. 2014, 144, 202–208. [Google Scholar] [CrossRef]
- Sun, C.; Wang, H.; Wang, D.; Chen, Y.; Zhao, Y.; Xia, W. Using an FFQ to Assess Intakes of Dietary Flavonols and Flavones Among Female Adolescents in The Suihua Area of Northern China. Public Health Nutr. 2015, 18, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nounou, M.I.; Ko, Y.; Helal, N.A.; Boltz, J.F. Adulteration and Counterfeiting of Online Nutraceutical Formulations in the United States: Time for Intervention? J. Diet. Suppl. 2018, 15, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Parihar; Arti; Duarte, S.; Doseff, A.I. The Saga of Food Supplements: Safety and Missing Knowledge—A Commentary. J. Med. Act. Plants 2015, 4, 16–19. [Google Scholar]
- Ronis, M.J.J.; Pedersen, K.B.; Watt, J. Adverse Effects of Nutraceuticals and Dietary Supplements. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 583–601. [Google Scholar] [CrossRef] [PubMed]
- Li, L.P.; Jiang, H.D. Determination and Assay Validation of Luteolin and Apigenin in Human Urine After Oral Administration of Tablet of Chrysanthemum Morifolium Extract by HPLC. J. Pharm. Biomed. Anal. 2006, 41, 261–265. [Google Scholar] [CrossRef]
- Aiello, P.; Consalvi, S.; Poce, G.; Raguzzini, A.; Toti, E.; Palmery, M.; Biava, M.; Bernardi, M.; Kamal, M.A.; Perry, G.; et al. Dietary flavonoids: Nano Delivery and Nanoparticles for Cancer Therapy. Semin. Cancer Biol. 2021, 69, 150–165. [Google Scholar] [CrossRef]
- Wu, C.; Xu, Q.; Chen, X.; Liu, J. Delivery Luteolin with Folacin-Modified Nanoparticle for Glioma Therapy. Int. J. Nanomed. 2019, 14, 7515–7531. [Google Scholar] [CrossRef] [Green Version]
- Ahn-Jarvis, J.H.; Parihar, A.; Doseff, A.I. Dietary Flavonoids for Immunoregulation and Cancer: Food Design for Targeting Disease. Antioxidants 2019, 8, 202. [Google Scholar] [CrossRef] [Green Version]
- Mallery, S.R.; Stoner, G.D.; Larsen, P.E.; Fields, H.W.; Rodrigo, K.A.; Schwartz, S.J.; Tian, Q.; Dai, J.; Mumper, R.J. Formulation and In-Vitro and In-Vivo Evaluation of a Mucoadhesive Gel Containing Freeze Dried Black Raspberries: Implications for Oral Cancer Chemoprevention. Pharm. Res. 2007, 24, 728–737. [Google Scholar] [CrossRef]
- Bosetti, C.; Spertini, L.; Parpinel, M.; Gnagnarella, P.; Lagiou, P.; Negri, E.; Franceschi, S.; Montella, M.; Peterson, J.; Dwyer, J.; et al. Flavonoids and Breast Cancer Risk in Italy. Cancer Epidemiol. Biomark. Prev. 2005, 14, 805–808. [Google Scholar] [CrossRef] [Green Version]
- Grosso, G.; Godos, J.; Lamuela-Raventos, R.; Ray, S.; Micek, A.; Pajak, A.; Sciacca, S.; D’Orazio, N.; Del Rio, D.; Galvano, F. A Comprehensive Meta-Analysis on Dietary Flavonoid and Lignan Intake and Cancer Risk: Level of Evidence and Limitations. Mol. Nutr. Food Res. 2017, 61, 1600930. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rodríguez, O.P.; Thompson-Bonilla, M.D.R.; Jaramillo-Flores, M.E. Association Between Obesity and Breast Cancer: Molecular Bases and the Effect of Flavonoids in Signaling Pathways. Crit. Rev. Food Sci. Nutr. 2020, 60, 3770–3792. [Google Scholar] [CrossRef] [PubMed]
- Rufino, A.T.; Costa, V.M.; Carvalho, F.; Fernandes, E. Flavonoids as Antiobesity Agents: A Review. Med. Res. Rev. 2021, 41, 556–585. [Google Scholar] [CrossRef]
- Bhosale, P.B.; Abusaliya, A.; Kim, H.H.; Ha, S.E.; Park, M.Y.; Jeong, S.H.; Vetrivel, P.; Heo, J.D.; Kim, J.A.; Won, C.K.; et al. Apigetrin Promotes TNFα-Induced Apoptosis, Necroptosis, G2/M Phase Cell Cycle Arrest, and ROS Generation through Inhibition of NF-κB Pathway in Hep3B Liver Cancer Cells. Cells 2022, 11, 2734. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Gupta, P.; Meena, A.; Luqman, S. Acacetin, a Flavone with Diverse Therapeutic Potential in Cancer, Inflammation, Infections and Other Metabolic Disorders. Food Chem. Toxicol. 2020, 145, 111708. [Google Scholar] [CrossRef]
- Zhou, G.; Cui, J.; Xie, S.; Wan, H.; Luo, Y.; Guo, G. Vitexin, a Fenugreek Glycoside, Ameliorated Obesity-Induced Diabetic Nephropathy Via Modulation of NF-Κb/Ikbα and AMPK/ACC Pathways in Mice. Biosci. Biotechnol. Biochem. 2021, 85, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Mandalari, G.; Calderaro, A.; Smeriglio, A.; Trombetta, D.; Felice, M.R.; Gattuso, G. Flavones: An Update on Sources, Biological Functions, and Health Promoting Properties. Plants 2020, 9, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodarzi, S.; Tabatabaei, M.J.; Mohammad Jafari, R.; Shemirani, F.; Tavakoli, S.; Mofasseri, M.; Tofighi, Z. Fruits as Source of Luteolin- 7-O-Glucoside, Potent Cytotoxic Flavonoid against Breast Cancer Cell Lines. Nat. Prod. Res. 2020, 34, 1602–1606. [Google Scholar] [CrossRef]
- Ożarowski, M.; Karpiński, T.M. Extracts and Flavonoids of Passiflora Species as Promising Anti-inflammatory and Antioxidant Substances. Curr. Pharm. Des. 2021, 27, 2582–2604. [Google Scholar] [CrossRef]
- Anilkumar, K.; Reddy, G.V.; Azad, R.; Yarla, N.S.; Dharmapuri, G.; Srivastava, A.; Kamal, M.A.; Pallu, R. Evaluation of Anti-Inflammatory Properties of Isoorientin Isolated from Tubers of Pueraria tuberosa. Oxid. Med. Cell Longev. 2017, 2017, 5498054. [Google Scholar] [CrossRef] [Green Version]
- Yáñez, A.; Coetzee, S.G.; Olsson, A.; Muench, D.E.; Berman, B.P.; Hazelett, D.J.; Salomonis, N.; Grimes, H.L.; Goodridge, H.S. Granulocyte-Monocyte Progenitors and Monocyte-Dendritic Cell Progenitors Independently Produce Functionally Distinct Monocytes. Immunity 2017, 47, 890–902.e4. [Google Scholar] [CrossRef] [PubMed]
- Canè, S.; Ugel, S.; Trovato, R.; Marigo, I.; De Sanctis, F.; Sartoris, S.; Bronte, V. The Endless Saga of Monocyte Diversity. Front. Immunol. 2019, 10, 1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dress, R.J.; Liu, Z.; Ginhoux, F. Towards the Better Understanding of Myelopoiesis Using Single-Cell Technologies. Mol. Immunol. 2020, 122, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Ostrand-Rosenberg, S.; Sinha, P. Myeloid-Derived Suppressor Cells: Linking Inflammation and Cancer. J. Immunol. 2009, 182, 4499–4506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; French, D.L.; Ma, G.; Eisenstein, S.; Chen, Y.; Divino, C.M.; Keller, G.; Chen, S.H.; Pan, P.Y. Development and Function of Myeloid-Derived Suppressor Cells Generated from Mouse Embryonic and Hematopoietic Stem Cells. Stem Cells 2010, 28, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Veglia, F.; Perego, M.; Gabrilovich, D. Myeloid-Derived Suppressor Cells Coming of Age. Nat. Immunol. 2018, 19, 108–119. [Google Scholar] [CrossRef]
- Bergenfelz, C.; Larsson, A.M.; von Stedingk, K.; Gruvberger-Saal, S.; Aaltonen, K.; Jansson, S.; Jernström, H.; Janols, H.; Wullt, M.; Bredberg, A.; et al. Systemic Monocytic-MDSCs Are Generated from Monocytes and Correlate with Disease Progression in Breast Cancer Patients. PLoS ONE 2015, 10, e0127028. [Google Scholar] [CrossRef] [Green Version]
- Bertola, A.; Ciucci, T.; Rousseau, D.; Bourlier, V.; Duffaut, C.; Bonnafous, S.; Blin-Wakkach, C.; Anty, R.; Iannelli, A.; Gugenheim, J.; et al. Identification of Adipose Tissue Dendritic Cells Correlated with Obesity-Associated Insulin-Resistance and Inducing Th17 Responses in Mice and Patients. Diabetes 2012, 61, 2238–2247. [Google Scholar] [CrossRef] [Green Version]
- Stefanovic-Racic, M.; Yang, X.; Turner, M.S.; Mantell, B.S.; Stolz, D.B.; Sumpter, T.L.; Sipula, I.J.; Dedousis, N.; Scott, D.K.; Morel, P.A.; et al. Dendritic Cells Promote Macrophage Infiltration and Comprise a Substantial Proportion of Obesity-Associated Increases in CD11c+ Cells in Adipose Tissue And Liver. Diabetes 2012, 61, 2330–2339. [Google Scholar] [CrossRef] [Green Version]
- Hannibal, T.D.; Schmidt-Christensen, A.; Nilsson, J.; Fransén-Pettersson, N.; Hansen, L.; Holmberg, D. Deficiency in Plasmacytoid Dendritic Cells and Type I Interferon Signalling Prevents Diet-Induced Obesity and Insulin Resistance in Mice. Diabetologia 2017, 60, 2033–2041. [Google Scholar] [CrossRef] [Green Version]
- Hernández-García, E.; Cueto, F.J.; Cook, E.C.L.; Redondo-Urzainqui, A.; Charro-Zanca, S.; Robles-Vera, I.; Conde-Garrosa, R.; Nikolić, I.; Sabio, G.; Sancho, D.; et al. Conventional Type 1 Dendritic Cells Protect against Age-Related Adipose Tissue Dysfunction and Obesity. Cell. Mol. Immunol. 2022, 19, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Duong, E.; Fessenden, T.B.; Lutz, E.; Dinter, T.; Yim, L.; Blatt, S.; Bhutkar, A.; Wittrup, K.D.; Spranger, S. Type I Interferon Activates MHC Class I-Dressed CD11b. Immunity 2022, 55, 308–323.e9. [Google Scholar] [CrossRef] [PubMed]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Miyazaki, M.; Ikeda, S.; Tsumura, H. Dendritic Cells Combined with Doxorubicin Induces Immunogenic Cell Death and Exhibits Antitumor Effects for Osteosarcoma. Oncol. Lett. 2016, 11, 2169–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, H.; Yin, X.; Bian, Y. Oxaliplatin Induces Immunogenic Cell Death in Human and Murine Laryngeal Cancer. J. Oncol. 2022, 2022, 3760766. [Google Scholar] [CrossRef] [PubMed]
- Boudewijns, S.; Westdorp, H.; Koornstra, R.H.; Aarntzen, E.H.; Schreibelt, G.; Creemers, J.H.; Punt, C.J.; Figdor, C.G.; de Vries, I.J.; Gerritsen, W.R.; et al. Immune-related Adverse Events of Dendritic Cell Vaccination Correlate with Immunologic and Clinical Outcome in Stage III and IV Melanoma Patients. J. Immunother. 2016, 39, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A.; de Mingo Pulido, Á.; Ruffell, B. Dendritic Cells and Their Role in Immunotherapy. Front. Immunol. 2020, 11, 924. [Google Scholar] [CrossRef]
- Gonzalez-Mejia, M.E.; Doseff, A.I. Regulation of Monocytes and Macrophages Cell Fate. Front. Biosci. 2009, 14, 2413–2431. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.A.; Zhang, Y.; Fullerton, J.N.; Boelen, L.; Rongvaux, A.; Maini, A.A.; Bigley, V.; Flavell, R.A.; Gilroy, D.W.; Asquith, B.; et al. The Fate and Lifespan of Human Monocyte Subsets in Steady State and Systemic Inflammation. J. Exp. Med. 2017, 214, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Jakubzick, C.V.; Randolph, G.J.; Henson, P.M. Monocyte Differentiation and Antigen-Presenting Functions. Nat. Rev. Immunol. 2017, 17, 349–362. [Google Scholar] [CrossRef]
- Coillard, A.; Segura, E. Differentiation of Human Monocytes. Front. Immunol. 2019, 10, 1907. [Google Scholar] [CrossRef] [Green Version]
- Villani, A.C.; Satija, R.; Reynolds, G.; Sarkizova, S.; Shekhar, K.; Fletcher, J.; Griesbeck, M.; Butler, A.; Zheng, S.; Lazo, S.; et al. Single-Cell RNA-Seq Reveals New Types of Human Blood Dendritic Cells, Monocytes, and Progenitors. Science 2017, 356, eaah4573. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Vadgama, J.V.; Wang, P. CCL2/CCR2 Signaling in Cancer Pathogenesis. Cell. Commun. Signal. 2020, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Simińska, D.; Kojder, K.; Grochans, S.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Fractalkine/CX3CL1 in Neoplastic Processes. Int. J. Mol. Sci. 2020, 21, 3723. [Google Scholar] [CrossRef] [PubMed]
- Hirose, S.; Lin, Q.; Ohtsuji, M.; Nishimura, H.; Verbeek, J.S. Monocyte Subsets Involved in the Development of Systemic Lupus Erythematosus and Rheumatoid Arthritis. Int. Immunol. 2019, 31, 687–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breznik, J.A.; Naidoo, A.; Foley, K.P.; Schulz, C.; Lau, T.C.; Loukov, D.; Sloboda, D.M.; Bowdish, D.M.E.; Schertzer, J.D. TNF, but not Hyperinsulinemia or Hyperglycemia, Is a Key Driver of Obesity-Induced Monocytosis Revealing That Inflammatory Monocytes Correlate With Insulin in Obese Male Mice. Physiol. Rep. 2018, 6, e13937. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, K.; Sommer, M.; Strobel, S.; Thrum, S.; Blüher, M.; Wagner, U.; Rossol, M. Perturbation of the Monocyte Compartment in Human Obesity. Front. Immunol. 2019, 10, 1874. [Google Scholar] [CrossRef] [Green Version]
- Cassetta, L.; Fragkogianni, S.; Sims, A.H.; Swierczak, A.; Forrester, L.M.; Zhang, H.; Soong, D.Y.H.; Cotechini, T.; Anur, P.; Lin, E.Y.; et al. Human Tumor-Associated Macrophage and Monocyte Transcriptional Landscapes Reveal Cancer-Specific Reprogramming, Biomarkers, and Therapeutic Targets. Cancer Cell. 2019, 35, 588–602.e10. [Google Scholar] [CrossRef] [Green Version]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-Resident Macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Papalexi, E.; Satija, R. Single-Cell RNA Sequencing to Explore Immune Cell Heterogeneity. Nat. Rev. Immunol. 2018, 18, 35–45. [Google Scholar] [CrossRef]
- Mills, C.D. Anatomy of a Discovery: M1 and M2 Macrophages. Front. Immunol. 2015, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Sun, Q. Macrophage recruitment in obese adipose tissue. Obes. Rev. 2015, 16, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouin, M.; Saenz, J.; Chiffoleau, E. C-Type Lectin-Like Receptors: Head or Tail in Cell Death Immunity. Front. Immunol. 2020, 11, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheedy, F.J.; Grebe, A.; Rayner, K.J.; Kalantari, P.; Ramkhelawon, B.; Carpenter, S.B.; Becker, C.E.; Ediriweera, H.N.; Mullick, A.E.; Golenbock, D.T.; et al. CD36 Coordinates NLRP3 Inflammasome Activation by Facilitating Intracellular Nucleation of Soluble Ligands into Particulate Ligands in Sterile Inflammation. Nat. Immunol. 2013, 14, 812–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, B.Z.; Zhang, H.; Li, J.; He, T.; Yeo, E.J.; Soong, D.Y.; Carragher, N.O.; Munro, A.; Chang, A.; Bresnick, A.R.; et al. FLT1 Signaling in Metastasis-Associated Macrophages Activates an Inflammatory Signature That Promotes Breast Cancer Metastasis. J. Exp. Med. 2015, 212, 1433–1448. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; DelProposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic Switching of Adipose Tissue Macrophages with Obesity Is Generated by Spatiotemporal Differences in Macrophage Subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, K.; Davis, A.T.; Peters, D.E.; LeGrand, A.C.; Bharadwaj, M.S.; Molina, A.J. Regulators of Mitochondrial Quality Control Differ in Subcutaneous Fat of Metabolically Healthy and Unhealthy Obese Monkeys. Obesity 2017, 25, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- Wagner, J.; Rapsomaniki, M.A.; Chevrier, S.; Anzeneder, T.; Langwieder, C.; Dykgers, A.; Rees, M.; Ramaswamy, A.; Muenst, S.; Soysal, S.D.; et al. A Single-Cell Atlas of the Tumor and Immune Ecosystem of Human Breast Cancer. Cell 2019, 177, 1330–1345.e18. [Google Scholar] [CrossRef] [Green Version]
- Santoni, M.; Romagnoli, E.; Saladino, T.; Foghini, L.; Guarino, S.; Capponi, M.; Giannini, M.; Cognigni, P.D.; Ferrara, G.; Battelli, N. Triple Negative Breast Cancer: Key Role of Tumor-Associated Macrophages in Regulating the Activity of Anti-PD-1/PD-L1 Agents. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 78–84. [Google Scholar] [CrossRef]
- Feng, Q.; Chang, W.; Mao, Y.; He, G.; Zheng, P.; Tang, W.; Wei, Y.; Ren, L.; Zhu, D.; Ji, M.; et al. Tumor-associated Macrophages as Prognostic and Predictive Biomarkers for Postoperative Adjuvant Chemotherapy in Patients with Stage II Colon Cancer. Clin. Cancer Res. 2019, 25, 3896–3907. [Google Scholar] [CrossRef]
- Xu, F.; Wei, Y.; Tang, Z.; Liu, B.; Dong, J. Tumor-Associated Macrophages in Lung Cancer: Friend or Foe? Mol. Med. Rep. 2020, 22, 4107–4115. [Google Scholar] [CrossRef] [PubMed]
- Donadon, M.; Torzilli, G.; Cortese, N.; Soldani, C.; Di Tommaso, L.; Franceschini, B.; Carriero, R.; Barbagallo, M.; Rigamonti, A.; Anselmo, A.; et al. Macrophage Morphology Correlates with Single-Cell Diversity and Prognosis in Colorectal Liver Metastasis. J. Exp. Med. 2020, 217, e20191847. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Song, K.; Zhou, J.; Shi, Y.H.; Liu, W.R.; Shi, G.M.; Gao, Q.; Wang, X.Y.; Ding, Z.B.; Fan, J. Tumor-Associated Macrophages Modulate Resistance to Oxaliplatin Via Inducing Autophagy in Hepatocellular Carcinoma. Cancer Cell Int. 2019, 19, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shree, T.; Olson, O.C.; Elie, B.T.; Kester, J.C.; Garfall, A.L.; Simpson, K.; Bell-McGuinn, K.M.; Zabor, E.C.; Brogi, E.; Joyce, J.A. Macrophages and Cathepsin Proteases Blunt Chemotherapeutic Response in Breast Cancer. Genes Dev. 2011, 25, 2465–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwada, K.; Kagawa, S.; Yoshida, R.; Sakamoto, S.; Ito, A.; Watanabe, M.; Ieda, T.; Kuroda, S.; Kikuchi, S.; Tazawa, H.; et al. The Epithelial-To-Mesenchymal Transition Induced by Tumor-Associated Macrophages Confers Chemoresistance in Peritoneally Disseminated Pancreatic Cancer. J. Exp. Clin. Cancer Res. 2018, 37, 307. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wei, C.; Liu, Y.; Qiu, Y.; Liu, C.; Guo, F. Selective Ablation of Tumor-Associated Macrophages Suppresses Metastasis and Angiogenesis. Cancer Sci. 2013, 104, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Marchesi, F.; Garlanda, C. Macrophages as Tools and Targets in Cancer Therapy. Nat. Rev. Drug Discov. 2022, 21, 799–820. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-Associated Macrophages in Tumor Metastasis: Biological Roles and Clinical Therapeutic Applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Xia, S.; Sha, H.; Yang, L.; Ji, Y.; Ostrand-Rosenberg, S.; Qi, L. Gr-1+ CD11b+ Myeloid-Derived Suppressor Cells Suppress Inflammation and Promote Insulin Sensitivity in Obesity. J. Biol. Chem. 2011, 286, 23591–23599. [Google Scholar] [CrossRef] [Green Version]
- Brcervic, I.; Doder, R.; Perisic, N.; Petrovic, S.; Vojvodic, D. Myeloid-Derived Suppressor Like Cells—whether Their Frequency Changes in Patients with Different Stages of CRC. Vojnosanit. Pregl. 2022, 91. [Google Scholar] [CrossRef]
- Kusmartsev, S.; Gabrilovich, D.I. Inhibition of Myeloid Cell Differentiation in Cancer: The Role of Reactive Oxygen Species. J. Leukoc. Biol. 2003, 74, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Cheng, P.; Condamine, T.; Mony, S.; Languino, L.R.; McCaffrey, J.C.; Hockstein, N.; Guarino, M.; Masters, G.; Penman, E.; et al. CD45 Phosphatase Inhibits STAT3 Transcription Factor Activity in Myeloid Cells and Promotes Tumor-Associated Macrophage Differentiation. Immunity 2016, 44, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.L.; Zhou, X.; Xue, Y.F.; Wang, K.; Shen, Y.F.; Mao, J.J.; Guo, H.F.; Miao, Z.N. Increased Frequency and Clinical Significance of Myeloid-Derived Suppressor Cells in Human Colorectal Carcinoma. World J. Gastroenterol. 2012, 18, 3303–3309. [Google Scholar] [CrossRef]
- Huang, A.; Zhang, B.; Wang, B.; Zhang, F.; Fan, K.X.; Guo, Y.J. Increased CD14(+) HLA-DR (-/low) Myeloid-Derived Suppressor Cells Correlate with Extrathoracic Metastasis and Poor Response to Chemotherapy in Non-Small Cell Lung Cancer Patients. Cancer Immunol. Immunother. 2013, 62, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Anestakis, D.; Petanidis, S.; Domvri, K.; Tsavlis, D.; Zarogoulidis, P.; Katopodi, T. Carboplatin Chemoresistance is Associated with CD11b. Mol. Immunol. 2020, 118, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB Signaling in Inflammation. Signal Transduct. Target Ther. 2017, 2, e17023. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-Κb Pathway for the Therapy of Diseases: Mechanism and Clinical Study. Signal Transduct. Target Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T.B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B.L.; et al. Inflammation, NF-κB, and Chronic Diseases: How are They Linked? Crit. Rev. Immunol. 2020, 40, 1–39. [Google Scholar] [CrossRef]
- He, F.; Huang, Y.; Song, Z.; Zhou, H.J.; Zhang, H.; Perry, R.J.; Shulman, G.I.; Min, W. Mitophagy-Mediated Adipose Inflammation Contributes to Type 2 Diabetes with Hepatic Insulin Resistance. J. Exp. Med. 2021, 218, e20201416. [Google Scholar] [CrossRef]
- Heida, A.; Gruben, N.; Catrysse, L.; Koehorst, M.; Koster, M.; Kloosterhuis, N.J.; Gerding, A.; Havinga, R.; Bloks, V.W.; Bongiovanni, L.; et al. The Hepatocyte IKK:NF-Κb Axis Promotes Liver Steatosis by Stimulating De Novo Lipogenesis and Cholesterol Synthesis. Mol. Metab. 2021, 54, 101349. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, M.H.; Nam, D.H.; Song, H.K.; Kang, Y.S.; Lee, J.E.; Kim, H.W.; Cha, J.J.; Hyun, Y.Y.; Han, S.Y.; et al. Celastrol, an NF-κB Inhibitor, Improves Insulin Resistance and Attenuates Renal Injury in Db/Db Mice. PLoS ONE 2013, 8, e62068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Guo, Y.; Cheng, Y.; Zhao, J.; Wang, Y.; Rong, J. Natural Product Celastrol Suppressed Macrophage M1 Polarization against Inflammation in Diet-Induced Obese Mice Via Regulating Nrf2/HO-1, MAP Kinase and NF-κB Pathways. Aging (Albany NY) 2017, 9, 2069–2082. [Google Scholar] [CrossRef] [Green Version]
- La Rosa, F.A.; Pierce, J.W.; Sonenshein, G.E. Differential Regulation of the C-Myc Oncogene Promoter by the NF-Kappa B Rel Family of Transcription Factors. Mol. Cell. Biol. 1994, 14, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Takebayashi, T.; Higashi, H.; Sudo, H.; Ozawa, H.; Suzuki, E.; Shirado, O.; Katoh, H.; Hatakeyama, M. NF-kappa B-Dependent Induction of Cyclin D1 by Retinoblastoma Protein (Prb) Family Proteins and Tumor-Derived Prb Mutants. J. Biol. Chem. 2003, 278, 14897–14905. [Google Scholar] [CrossRef] [Green Version]
- Pires, B.R.; Mencalha, A.L.; Ferreira, G.M.; de Souza, W.F.; Morgado-Díaz, J.A.; Maia, A.M.; Corrêa, S.; Abdelhay, E.S. NF-kappaB Is Involved in the Regulation of EMT Genes in Breast Cancer Cells. PLoS ONE 2017, 12, e0169622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, C.F.; Chao, T.T.; Su, Y.F.; Hsu, C.C.; Chien, C.Y.; Chiu, K.C.; Shiah, S.G.; Lee, C.H.; Liu, S.Y.; Shieh, Y.S. Metformin-Treated Cancer Cells Modulate Macrophage Polarization through AMPK-NF-Κb Signaling. Oncotarget 2017, 8, 20706–20718. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, K.; Karin, M. NF-κB, Inflammation, Immunity and Cancer: Coming of Age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Eluard, B.; Thieblemont, C.; Baud, V. NF-κB in the New Era of Cancer Therapy. Trends Cancer 2020, 6, 677–687. [Google Scholar] [CrossRef]
- Suhail, M.; Tarique, M.; Muhammad, N.; Naz, H.; Hafeez, A.; Zughaibi, T.A.; Kamal, M.A.; Rehan, M. A Critical Transcription Factor NF-κB as a Cancer Therapeutic Target and its Inhibitors as Cancer Treatment Options. Curr. Med. Chem. 2021, 28, 4117–4132. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zeng, H.; Jin, K.; Liu, Z.; Hu, B.; Liu, C.; Yan, S.; Yu, Y.; You, R.; Zhang, H.; et al. Infiltration and Polarization of Tumor-associated Macrophages Predict Prognosis and Therapeutic Benefit in Muscle-Invasive Bladder Cancer. Cancer Immunol. Immunother. 2022, 71, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Verma, R.K.; Alpini, G.; Meng, F.; Kumar, N. Role of Janus Kinase 3 in Predisposition to Obesity-associated Metabolic Syndrome. J. Biol. Chem. 2015, 290, 29301–29312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrian, A.D.; Galkina, E.V.; Ma, Q.; Hatcher, M.; Aye, S.M.; Butcher, M.J.; Ma, K.; Haynes, B.A.; Kaplan, M.H.; Nadler, J.L. STAT4 Deficiency Reduces Obesity-Induced Insulin Resistance and Adipose Tissue Inflammation. Diabetes 2013, 62, 4109–4121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, H.; Xu, D.; Zhao, J.; Li, J.; Wang, Q.; Tian, X.; Li, M.; Zeng, X. JAK Inhibitors: Prospects in Connective Tissue Diseases. Clin. Rev. Allergy Immunol. 2020, 59, 334–351. [Google Scholar] [CrossRef] [PubMed]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Antonucci, L.; Karin, M. NRF2 as a Regulator of Cell Metabolism and Inflammation in Cancer. Carcinogenesis 2020, 41, 405–416. [Google Scholar] [CrossRef]
- Ganesh Yerra, V.; Negi, G.; Sharma, S.S.; Kumar, A. Potential Therapeutic Effects of the Simultaneous Targeting of the Nrf2 and NF-Κb Pathways in Diabetic Neuropathy. Redox. Biol. 2013, 1, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 Suppresses Macrophage Inflammatory Response by Blocking Proinflammatory Cytokine Transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Wakabayashi, J.; Yates, M.S.; Wakabayashi, N.; Dolan, P.M.; Aja, S.; Liby, K.T.; Sporn, M.B.; Yamamoto, M.; Kensler, T.W. Role of Nrf2 in Prevention of High-Fat Diet-Induced Obesity by Synthetic Triterpenoid CDDO-Imidazolide. Eur. J. Pharmacol. 2009, 620, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Slocum, S.L.; Skoko, J.J.; Wakabayashi, N.; Aja, S.; Yamamoto, M.; Kensler, T.W.; Chartoumpekis, D.V. Keap1/Nrf2 Pathway Activation Leads to a Repressed Hepatic Gluconeogenic and Lipogenic Program in Mice on a High-Fat Diet. Arch. Biochem. Biophys. 2016, 591, 57–65. [Google Scholar] [CrossRef]
- Chartoumpekis, D.V.; Palliyaguru, D.L.; Wakabayashi, N.; Fazzari, M.; Khoo, N.K.H.; Schopfer, F.J.; Sipula, I.; Yagishita, Y.; Michalopoulos, G.K.; O’Doherty, R.M.; et al. Nrf2 Deletion from Adipocytes, but not Hepatocytes, Potentiates Systemic Metabolic Dysfunction After Long-Term High-Fat Diet-Induced Obesity in Mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E180–E195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Gomez, M.; Kwak, M.K.; Dolan, P.M.; Itoh, K.; Yamamoto, M.; Talalay, P.; Kensler, T.W. Sensitivity to Carcinogenesis Is Increased and Chemoprotective Efficacy of Enzyme Inducers Is Lost in Nrf2 Transcription Factor-Deficient Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 3410–3415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, T.O.; Huang, M.T.; Prawan, A.; Liu, Y.; Hao, X.; Yu, S.; Cheung, W.K.; Chan, J.Y.; Reddy, B.S.; Yang, C.S.; et al. Increased Susceptibility of Nrf2 Knockout Mice to Colitis-Associated Colorectal Cancer. Cancer Prev. Res. 2008, 1, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Ma, D.; Wang, P.; Pan, C.; Fang, Q.; Wang, J. Nrf2 Overexpression Increases Risk of High Tumor Mutation Burden in Acute Myeloid Leukemia by Inhibiting MSH2. Cell Death Dis. 2021, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Knatko, E.V.; Higgins, M.; Dayalan Naidu, S.; Smith, G.; Honda, T.; de la Vega, L.; Dinkova-Kostova, A.T. Pirin, an Nrf2-Regulated Protein, Is Overexpressed in Human Colorectal Tumors. Antioxidants 2022, 11, 262. [Google Scholar] [CrossRef]
- Ji, X.J.; Chen, S.H.; Zhu, L.; Pan, H.; Zhou, Y.; Li, W.; You, W.C.; Gao, C.C.; Zhu, J.H.; Jiang, K.; et al. Knockdown of NF-E2-Related Factor 2 Inhibits the Proliferation and Growth of U251MG Human Glioma Cells in a Mouse Xenograft Model. Oncol. Rep. 2013, 30, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, J.L.; Kim, E.H.; Jang, H.; Shin, D. Nrf2 Inhibition Reverses the Resistance of Cisplatin-Resistant Head and Neck Cancer Cells to Artesunate-Induced Ferroptosis. Redox. Biol. 2017, 11, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hou, Z.; Aldrich, K.E.; Lockwood, L.; Odom, A.L.; Liby, K.T. A Novel Nrf2 Pathway Inhibitor Sensitizes Keap1-Mutant Lung Cancer Cells to Chemotherapy. Mol. Cancer Ther. 2021, 20, 1692–1701. [Google Scholar] [CrossRef]
- Ohl, K.; Tenbrock, K. Reactive Oxygen Species as Regulators of MDSC-Mediated Immune Suppression. Front. Immunol. 2018, 9, 2499. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.R.; Kanneganti, T.D. NLRP3 Inflammasome in Cancer and Metabolic Diseases. Nat. Immunol. 2021, 22, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhong, H.; Wei, J.; Lin, S.; Zong, Z.; Gong, F.; Huang, X.; Sun, J.; Li, P.; Lin, H.; et al. Inhibition of Nrf2/HO-1 Signaling Leads to Increased Activation of the NLRP3 Inflammasome in Osteoarthritis. Arthritis Res. Ther. 2019, 21, 300. [Google Scholar] [CrossRef] [Green Version]
- Stienstra, R.; van Diepen, J.A.; Tack, C.J.; Zaki, M.H.; van de Veerdonk, F.L.; Perera, D.; Neale, G.A.; Hooiveld, G.J.; Hijmans, A.; Vroegrijk, I.; et al. Inflammasome is a Central Player in the Induction of Obesity and Insulin Resistance. Proc. Natl. Acad. Sci. USA 2011, 108, 15324–15329. [Google Scholar] [CrossRef] [Green Version]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 Inflammasome Instigates Obesity-Induced Inflammation and Insulin Resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Rheinheimer, J.; de Souza, B.M.; Cardoso, N.S.; Bauer, A.C.; Crispim, D. Current Role of the NLRP3 Inflammasome on Obesity and Insulin Resistance: A Systematic Review. Metabolism 2017, 74, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zaki, M.H.; Vogel, P.; Body-Malapel, M.; Lamkanfi, M.; Kanneganti, T.D. IL-18 Production Downstream of the Nlrp3 Inflammasome Confers Protection against Colorectal Tumor Formation. J. Immunol. 2010, 185, 4912–4920. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Fu, S.; Zhang, J.; Liu, B.; Li, Z. Targeting Inflammasome/IL-1 Pathways for Cancer Immunotherapy. Sci. Rep. 2016, 6, 36107. [Google Scholar] [CrossRef] [Green Version]
- Bruchard, M.; Mignot, G.; Derangère, V.; Chalmin, F.; Chevriaux, A.; Végran, F.; Boireau, W.; Simon, B.; Ryffel, B.; Connat, J.L.; et al. Chemotherapy-Triggered Cathepsin B Release in Myeloid-Derived Suppressor Cells Activates the Nlrp3 Inflammasome and Promotes Tumor Growth. Nat. Med. 2013, 19, 57–64. [Google Scholar] [CrossRef]
- Cho, K.W.; Zamarron, B.F.; Muir, L.A.; Singer, K.; Porsche, C.E.; DelProposto, J.B.; Geletka, L.; Meyer, K.A.; O’Rourke, R.W.; Lumeng, C.N. Adipose Tissue Dendritic Cells Are Independent Contributors to Obesity-Induced Inflammation and Insulin Resistance. J. Immunol. 2016, 197, 3650–3661. [Google Scholar] [CrossRef] [Green Version]
- Aouadi, M.; Tencerova, M.; Vangala, P.; Yawe, J.C.; Nicoloro, S.M.; Amano, S.U.; Cohen, J.L.; Czech, M.P. Gene Silencing in Adipose Tissue Macrophages Regulates Whole-Body Metabolism in Obese Mice. Proc. Natl. Acad. Sci. USA 2013, 110, 8278–8283. [Google Scholar] [CrossRef] [Green Version]
- Muir, L.A.; Kiridena, S.; Griffin, C.; DelProposto, J.B.; Geletka, L.; Martinez-Santibañez, G.; Zamarron, B.F.; Lucas, H.; Singer, K.; O’ Rourke, R.W.; et al. Frontline Science: Rapid Adipose Tissue Expansion Triggers Unique Proliferation and Lipid Accumulation Profiles in Adipose Tissue Macrophages. J. Leukoc. Biol. 2018, 103, 615–628. [Google Scholar] [CrossRef]
- Aarts, S.; Reiche, M.; den Toom, M.; Gijbels, M.; Beckers, L.; Gerdes, N.; Lutgens, E. Depletion of CD40 on CD11c. Sci. Rep. 2019, 9, 14702. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Mo, J.; Ruan, L.; Li, G. Increased Monocytic CD14⁺Hladrlow/- Myeloid-Derived Suppressor Cells in Obesity. Mol. Med. Rep. 2015, 11, 2322–2328. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Ruiz, J.C.; Galindo-De Ávila, J.C.; Martínez-Fierro, M.L.; Garza-Veloz, I.; Cervantes-Villagrana, A.R.; Valtierra-Alvarado, M.A.; Serrano, C.J.; García-Hernández, M.H.; Enciso-Moreno, J.A.; Castañeda-Delgado, J.E. Myeloid-Derived Suppressor Cells Show Different Frequencies in Diabetics and Subjects with Arterial Hypertension. J. Diabetes Res. 2019, 2019, 1568457. [Google Scholar] [CrossRef] [Green Version]
- Del Fabbro, E.; Parsons, H.; Warneke, C.L.; Pulivarthi, K.; Litton, J.K.; Dev, R.; Palla, S.L.; Brewster, A.; Bruera, E. The Relationship Between Body Composition and Response to Neoadjuvant Chemotherapy in Women with Operable Breast Cancer. Oncologist 2012, 17, 1240–1245. [Google Scholar] [CrossRef] [Green Version]
- Hidayat, K.; Du, X.; Chen, G.; Shi, M.; Shi, B. Abdominal Obesity and Lung Cancer Risk: Systematic Review and Meta-Analysis of Prospective Studies. Nutrients 2016, 8, 810. [Google Scholar] [CrossRef]
- Genkinger, J.M.; Kitahara, C.M.; Bernstein, L.; Berrington de Gonzalez, A.; Brotzman, M.; Elena, J.W.; Giles, G.G.; Hartge, P.; Singh, P.N.; Stolzenberg-Solomon, R.Z.; et al. Central Adiposity, Obesity During Early Adulthood, and Pancreatic Cancer Mortality in a Pooled Analysis of Cohort Studies. Ann. Oncol. 2015, 26, 2257–2266. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.S.M.; Vieira, A.R.; Aune, D.; Bandera, E.V.; Greenwood, D.C.; McTiernan, A.; Navarro Rosenblatt, D.; Thune, I.; Vieira, R.; Norat, T. Body Mass Index and Survival in Women With Breast Cancer-Systematic Literature Review and Meta-Analysis of 82 Follow-Up Studies. Ann. Oncol. 2014, 25, 1901–1914. [Google Scholar] [CrossRef]
- Heetun, A.; Cutress, R.I.; Copson, E.R. Early Breast Cancer: Why Does Obesity Affect Prognosis? Proc. Nutr. Soc. 2018, 77, 369–381. [Google Scholar] [CrossRef]
- Playdon, M.C.; Bracken, M.B.; Sanft, T.B.; Ligibel, J.A.; Harrigan, M.; Irwin, M.L. Weight Gain After Breast Cancer Diagnosis and All-Cause Mortality: Systematic Review and Meta-Analysis. J. Natl. Cancer Inst. 2015, 107, djv275. [Google Scholar] [CrossRef] [Green Version]
- Playdon, M.; Thomas, G.; Sanft, T.; Harrigan, M.; Ligibel, J.; Irwin, M. Weight Loss Intervention for Breast Cancer Survivors: A Systematic Review. Curr. Breast Cancer Rep. 2013, 5, 222–246. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.Q.; Du, W.L.; Cai, M.H.; Yao, J.Y.; Zhao, Y.Y.; Mou, X.Z. The Roles of Tumor-Associated Macrophages in Tumor Angiogenesis And Metastasis. Cell Immunol. 2020, 353, 104119. [Google Scholar] [CrossRef]
- Hale, M.; Itani, F.; Buchta, C.M.; Wald, G.; Bing, M.; Norian, L.A. Obesity Triggers Enhanced MDSC Accumulation in Murine Renal Tumors Via Elevated Local Production of CCL2. PLoS ONE 2015, 10, e0118784. [Google Scholar] [CrossRef] [Green Version]
- Olingy, C.E.; Dinh, H.Q.; Hedrick, C.C. Monocyte Heterogeneity and Functions in Cancer. J. Leukoc. Biol. 2019, 106, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Gouveia-Fernandes, S. Monocytes and Macrophages in Cancer: Unsuspected Roles. Adv. Exp. Med. Biol. 2020, 1219, 161–185. [Google Scholar] [CrossRef]
- Mehta, A.K.; Kadel, S.; Townsend, M.G.; Oliwa, M.; Guerriero, J.L. Macrophage Biology and Mechanisms of Immune Suppression in Breast Cancer. Front. Immunol. 2021, 12, 643771. [Google Scholar] [CrossRef]
- Zeisberger, S.M.; Odermatt, B.; Marty, C.; Zehnder-Fjällman, A.H.; Ballmer-Hofer, K.; Schwendener, R.A. Clodronate-Liposome-Mediated Depletion of Tumour-Associated Macrophages: A New and Highly Effective Antiangiogenic Therapy Approach. Br. J. Cancer 2006, 95, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Mitchem, J.B.; Brennan, D.J.; Knolhoff, B.L.; Belt, B.A.; Zhu, Y.; Sanford, D.E.; Belaygorod, L.; Carpenter, D.; Collins, L.; Piwnica-Worms, D.; et al. Targeting Tumor-Infiltrating Macrophages Decreases Tumor-Initiating Cells, Relieves Immunosuppression, and Improves Chemotherapeutic Responses. Cancer Res. 2013, 73, 1128–1141. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Pang, Y.; Moses, H.L. TGF-beta and Immune Cells: An Important Regulatory Axis in The Tumor Microenvironment and Progression. Trends Immunol. 2010, 31, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Saccani, A.; Schioppa, T.; Porta, C.; Biswas, S.K.; Nebuloni, M.; Vago, L.; Bottazzi, B.; Colombo, M.P.; Mantovani, A.; Sica, A. p50 Nuclear Factor-Kappab Overexpression in Tumor-Associated Macrophages Inhibits M1 Inflammatory Responses and Antitumor Resistance. Cancer Res. 2006, 66, 11432–11440. [Google Scholar] [CrossRef] [PubMed]
- Roland, C.L.; Lynn, K.D.; Toombs, J.E.; Dineen, S.P.; Udugamasooriya, D.G.; Brekken, R.A. Cytokine Levels Correlate with Immune Cell Infiltration After Anti-VEGF Therapy in Preclinical Mouse Models of Breast Cancer. PLoS ONE 2009, 4, e7669. [Google Scholar] [CrossRef] [PubMed]
- Vicari, A.P.; Chiodoni, C.; Vaure, C.; Aït-Yahia, S.; Dercamp, C.; Matsos, F.; Reynard, O.; Taverne, C.; Merle, P.; Colombo, M.P.; et al. Reversal of Tumor-Induced Dendritic Cell Paralysis by Cpg Immunostimulatory Oligonucleotide and Anti-Interleukin 10 Receptor Antibody. J. Exp. Med. 2002, 196, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.; Karin, E.; Terzic, J.; Mucida, D.; Yu, G.Y.; Vallabhapurapu, S.; Scheller, J.; Rose-John, S.; Cheroutre, H.; Eckmann, L.; et al. IL-6 and Stat3 Are Required for Survival of Intestinal Epithelial Cells and Development of Colitis-Associated Cancer. Cancer Cell 2009, 15, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Mohammadpour, H.; MacDonald, C.R.; Qiao, G.; Chen, M.; Dong, B.; Hylander, B.L.; McCarthy, P.L.; Abrams, S.I.; Repasky, E.A. β2 Adrenergic Receptor-Mediated Signaling Regulates the Immunosuppressive Potential of Myeloid-Derived Suppressor Cells. J. Clin. Investig. 2019, 129, 5537–5552. [Google Scholar] [CrossRef] [Green Version]
- Clements, V.K.; Long, T.; Long, R.; Figley, C.; Smith, D.M.C.; Ostrand-Rosenberg, S. Frontline Science: High Fat Diet and Leptin Promote Tumor Progression by Inducing Myeloid-Derived Suppressor Cells. J. Leukoc. Biol. 2018, 103, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Guo, T.; Deng, R.; Liu, L.; Yu, Y. Apigenin Ameliorates Insulin Resistance and Lipid Accumulation by Endoplasmic Reticulum Stress and SREBP-1c/SREBP-2 Pathway in Palmitate-Induced HepG2 Cells and High-Fat Diet-Fed Mice. J. Pharmacol. Exp. Ther. 2021, 377, 146–156. [Google Scholar] [CrossRef]
- Baek, Y.; Lee, M.N.; Wu, D.; Pae, M. Luteolin Reduces Adipose Tissue Macrophage Inflammation and Insulin Resistance in Postmenopausal Obese Mice. J. Nutr. Biochem. 2019, 71, 72–81. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, X. Luteolin Alleviates Polycystic Ovary Syndrome in Rats by Resolving Insulin Resistance and Oxidative Stress. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E1085–E1092. [Google Scholar] [CrossRef]
- Jannasch, F.; Kröger, J.; Schulze, M.B. Dietary Patterns and Type 2 Diabetes: A Systematic Literature Review and Meta-Analysis of Prospective Studies. J. Nutr. 2017, 147, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Steffen, L.M.; Van Horn, L.; Daviglus, M.L.; Zhou, X.; Reis, J.P.; Loria, C.M.; Jacobs, D.R.; Duffey, K.J. A Modified Mediterranean Diet Score Is Associated with a Lower Risk of Incident Metabolic Syndrome Over 25 Years Among Young Adults: The CARDIA (Coronary Artery Risk Development in Young Adults) Study. Br. J. Nutr. 2014, 112, 1654–1661. [Google Scholar] [CrossRef] [Green Version]
- Mancini, J.G.; Filion, K.B.; Atallah, R.; Eisenberg, M.J. Systematic Review of the Mediterranean Diet for Long-Term Weight Loss. Am. J. Med. 2016, 129, 407–415.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, J.K.; Shankar, E.; Gupta, S. Chamomile: A Herbal Medicine of the Past with Bright Future. Mol. Med. Rep. 2010, 3, 895–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miguel, F.G.; Cavalheiro, A.H.; Spinola, N.F.; Ribeiro, D.L.; Barcelos, G.R.; Antunes, L.M.; Hori, J.I.; Marquele-Oliveira, F.; Rocha, B.A.; Berretta, A.A. Validation of a RP-HPLC-DAD Method for Chamomile (Matricaria recutita) Preparations and Assessment of the Marker, Apigenin-7-glucoside, Safety and Anti-Inflammatory Effect. Evid. Based Complement. Alternat. Med. 2015, 2015, 828437. [Google Scholar] [CrossRef] [Green Version]
- Rafraf, M.; Zemestani, M.; Asghari-Jafarabadi, M. Effectiveness of Chamomile Tea on Glycemic Control and Serum Lipid Profile in Patients with Type 2 Diabetes. J. Endocrinol. Investig. 2015, 38, 163–170. [Google Scholar] [CrossRef]
- Kato, A.; Minoshima, Y.; Yamamoto, J.; Adachi, I.; Watson, A.A.; Nash, R.J. Protective Effects of Dietary Chamomile Tea on Diabetic Complications. J. Agric. Food Chem. 2008, 56, 8206–8211. [Google Scholar] [CrossRef]
- Morze, J.; Danielewicz, A.; Przybyłowicz, K.; Zeng, H.; Hoffmann, G.; Schwingshackl, L. An Updated Systematic Review and Meta-Analysis on Adherence to Mediterranean Diet and Risk of Cancer. Eur. J. Nutr. 2021, 60, 1561–1586. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Schwedhelm, C.; Galbete, C.; Hoffmann, G. Adherence to Mediterranean Diet and Risk of Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2017, 9, 1063. [Google Scholar] [CrossRef]
- Gates, M.A.; Vitonis, A.F.; Tworoger, S.S.; Rosner, B.; Titus-Ernstoff, L.; Hankinson, S.E.; Cramer, D.W. Flavonoid Intake and Ovarian Cancer Risk in a Population-Based Case-Control Study. Int. J. Cancer 2009, 124, 1918–1925. [Google Scholar] [CrossRef] [Green Version]
- Van Blarigan, E.L.; Fuchs, C.S.; Niedzwiecki, D.; Zhang, S.; Saltz, L.B.; Mayer, R.J.; Mowat, R.B.; Whittom, R.; Hantel, A.; Benson, A.; et al. Association of Survival with Adherence to the American Cancer Society Nutrition and Physical Activity Guidelines for Cancer Survivors After Colon Cancer Diagnosis: The CALGB 89803/Alliance Trial. JAMA Oncol. 2018, 4, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Arango, D.; Diosa-Toro, M.; Rojas-Hernandez, L.S.; Cooperstone, J.L.; Schwartz, S.J.; Mo, X.; Jiang, J.; Schmittgen, T.D.; Doseff, A.I. Dietary Apigenin Reduces LPS-Induced Expression of Mir-155 Restoring Immune Balance During Inflammation. Mol. Nutr. Food Res. 2015, 59, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Weng, D.; Zhou, F.; Owen, Y.D.; Qin, H.; Zhao, J.; Wen, Y.; Huang, Y.; Chen, J.; Fu, H.; et al. Activation of PPARγ by a Natural Flavonoid Modulator, Apigenin Ameliorates Obesity-Related Inflammation Via Regulation of Macrophage Polarization. EBioMedicine 2016, 9, 61–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalobos-Ayala, K.; Ortiz Rivera, I.; Alvarez, C.; Husain, K.; DeLoach, D.; Krystal, G.; Hibbs, M.L.; Jiang, K.; Ghansah, T. Apigenin Increases SHIP-1 Expression, Promotes Tumoricidal Macrophages and Anti-Tumor Immune Responses in Murine Pancreatic Cancer. Cancers 2020, 12, 3631. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhao, D.; Zhou, H.G.; Wang, X.H.; Zhong, W.L.; Chen, S.; Gu, W.G.; Wang, W.; Zhang, C.H.; Liu, Y.R.; et al. Apigenin Inhibits NF-Κb and Snail Signaling, EMT and Metastasis in Human Hepatocellular Carcinoma. Oncotarget 2016, 7, 41421–41431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.; Shankar, E.; Fu, P.; MacLennan, G.T.; Gupta, S. Suppression of NF-κB and NF-κB-Regulated Gene Expression by Apigenin through IκBα and IKK Pathway in TRAMP Mice. PLoS ONE 2015, 10, e0138710. [Google Scholar] [CrossRef] [Green Version]
- Bauer, D.; Redmon, N.; Mazzio, E.; Soliman, K.F. Apigenin Inhibits Tnfα/IL-1α-Induced CCL2 Release through IKBK-Epsilon Signaling in MDA-MB-231 Human Breast Cancer Cells. PLoS ONE 2017, 12, e0175558. [Google Scholar] [CrossRef] [Green Version]
- Liou, C.J.; Wu, S.J.; Chen, L.C.; Yeh, K.W.; Chen, C.Y.; Huang, W.C. Acacetin from Traditionally Used Saussurea involucrata Kar. Et. Kir. Suppressed Adipogenesis in 3T3-L1 Adipocytes and Attenuated Lipid Accumulation in Obese Mice. Front. Pharmacol. 2017, 8, 589. [Google Scholar] [CrossRef] [Green Version]
- Noh, J.W.; Kwon, O.J.; Lee, B.C. The Immunomodulating Effect of Baicalin on Inflammation and Insulin Resistance in High-Fat-Diet-Induced Obese Mice. Evid. Based Complement. Alternat. Med. 2021, 2021, 5531367. [Google Scholar] [CrossRef]
- He, S.; Wang, S.; Liu, S.; Li, Z.; Liu, X.; Wu, J. Baicalein Potentiated M1 Macrophage Polarization in Cancer through Targeting PI3Kγ/ NF-κB Signaling. Front. Pharmacol. 2021, 12, 743837. [Google Scholar] [CrossRef]
- Feng, X.; Qin, H.; Shi, Q.; Zhang, Y.; Zhou, F.; Wu, H.; Ding, S.; Niu, Z.; Lu, Y.; Shen, P. Chrysin Attenuates Inflammation by Regulating M1/M2 Status Via Activating PPARγ. Biochem. Pharmacol. 2014, 89, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.Y.; Choi, M.S. Luteolin Targets the Toll-Like Receptor Signaling Pathway in Prevention of Hepatic and Adipocyte Fibrosis and Insulin Resistance in Diet-Induced Obese Mice. Nutrients 2018, 10, 1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkoong, S.; Sung, J.; Yang, J.; Choi, Y.; Jeong, H.S.; Lee, J. Nobiletin Attenuates the Inflammatory Response through Heme Oxygenase-1 Induction in the Crosstalk Between Adipocytes and Macrophages. J. Med. Food 2017, 20, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, L.; Qiu, C. Tangeretin Protects Mice from Diet-Induced Metabolic Inflammation Via Activating Adipose Lactate Accumulation and Macrophage M2 Polarization. Biochem. Biophys. Res. Commun. 2022, 630, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, B.; Yuan, X.; Lu, Y.; Hu, J.; Gao, J.; Lin, J.; Liang, J.; Hou, S.; Chen, S. Vitexin Prevents Colitis-Associated Carcinogenesis in Mice through Regulating Macrophage Polarization. Phytomedicine 2021, 83, 153489. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, Y.; Tian, K.; Chen, X.; Zhang, R.; Mu, X.; Wu, Y.; Wang, D.; Wang, S.; Liu, F.; et al. Apigenin Suppresses PD-L1 Expression in Melanoma and Host Dendritic Cells to Elicit Synergistic Therapeutic Effects. J. Exp. Clin. Cancer Res. 2018, 37, 261. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.B.; Wang, W.J.; Xu, C.; Xie, Y.J.; Wang, X.R.; Zhang, Y.Z.; Huang, J.M.; Huang, M.; Xie, C.; Liu, P.; et al. Luteolin and Its Derivative Apigenin Suppress the Inducible PD-L1 Expression to Improve Anti-Tumor Immunity in KRAS-Mutant Lung Cancer. Cancer Lett. 2021, 515, 36–48. [Google Scholar] [CrossRef]
- Nunes, C.; Almeida, L.; Barbosa, R.M.; Laranjinha, J. Luteolin Suppresses the JAK/STAT Pathway in a Cellular Model of Intestinal Inflammation. Food Funct. 2017, 8, 387–396. [Google Scholar] [CrossRef]
- Aneknan, P.; Kukongviriyapan, V.; Prawan, A.; Kongpetch, S.; Sripa, B.; Senggunprai, L. Luteolin Arrests Cell Cycling, Induces Apoptosis and Inhibits the JAK/STAT3 Pathway in Human Cholangiocarcinoma Cells. Asian Pac. J. Cancer Prev. 2014, 15, 5071–5076. [Google Scholar] [CrossRef] [Green Version]
- Paredes-Gonzalez, X.; Fuentes, F.; Jeffery, S.; Saw, C.L.; Shu, L.; Su, Z.Y.; Kong, A.N. Induction of NRF2-Mediated Gene Expression by Dietary Phytochemical Flavones Apigenin and Luteolin. Biopharm. Drug Dispos. 2015, 36, 440–451. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Zhang, X.; Wang, L.; Hao, T.; Cheng, Y.; Wang, D. Scutellarin Exerts Hypoglycemic and Renal Protective Effects in db/db Mice Via the Nrf2/HO-1 Signaling Pathway. Oxid. Med. Cell Longev. 2019, 2019, 1354345. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Feng, S.L.; He, F.; Yan, P.Y.; Yao, X.J.; Fan, X.X.; Leung, E.L.; Zhou, H. Down-Regulating Nrf2 by Tangeretin Reverses Multiple Drug Resistance to Both Chemotherapy and EGFR Tyrosine Kinase Inhibitors in Lung Cancer. Pharmacol. Res. 2022, 186, 106514. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Zhang, F.; Sun, Z.; Zhou, W.; Li, Z.Y.; You, Q.D.; Guo, Q.L.; Hu, R. Drug Resistance Associates with Activation of Nrf2 in MCF-7/DOX Cells, and Wogonin Reverses It by Down-Regulating Nrf2-Mediated Cellular Defense Response. Mol. Carcinog. 2013, 52, 824–834. [Google Scholar] [CrossRef]
- Lee, M.N.; Lee, Y.; Wu, D.; Pae, M. Luteolin Inhibits NLRP3 Inflammasome Activation Via Blocking ASC Oligomerization. J. Nutr. Biochem. 2021, 92, 108614. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ma, S.; Pi, D.; Wu, Y.; Zuo, Q.; Li, C.; Ouyang, M. Luteolin Induces Pyroptosis in HT-29 Cells by Activating the Caspase1/Gasdermin D Signalling Pathway. Front. Pharmacol. 2022, 13, 952587. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Moore, L.B.; Smith-Oliver, T.A.; Wilkison, W.O.; Willson, T.M.; Kliewer, S.A. An Antidiabetic Thiazolidinedione Is a High Affinity Ligand for Peroxisome Proliferator-Activated Receptor Gamma (PPAR Gamma). J. Biol. Chem. 1995, 270, 12953–12956. [Google Scholar] [CrossRef] [Green Version]
- Woo, K.J.; Jeong, Y.J.; Inoue, H.; Park, J.W.; Kwon, T.K. Chrysin Suppresses Lipopolysaccharide-Induced Cyclooxygenase-2 Expression through the Inhibition of Nuclear Factor for IL-6 (NF-IL6) DNA-Binding Activity. FEBS Lett. 2005, 579, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, C.; Batra, S.; Vargo, M.A.; Voss, O.H.; Gavrilin, M.A.; Wewers, M.D.; Guttridge, D.C.; Grotewold, E.; Doseff, A.I. Apigenin Blocks Lipopolysaccharide-Induced Lethality In Vivo And Proinflammatory Cytokines Expression by Inactivating NF-KappaB through the Suppression of p65 Phosphorylation. J. Immunol. 2007, 179, 7121–7127. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, H.; Arango, D.; Nicholas, C.; Duarte, S.; Nuovo, G.J.; He, W.; Voss, O.H.; Gonzalez-Mejia, M.E.; Guttridge, D.C.; Grotewold, E.; et al. Dietary Apigenin Exerts Immune-Regulatory Activity in Vivo by Reducing NF-κB Activity, Halting Leukocyte Infiltration and Restoring Normal Metabolic Function. Int. J. Mol. Sci. 2016, 17, 323. [Google Scholar] [CrossRef] [Green Version]
- Coll, R.C.; Robertson, A.A.; Chae, J.J.; Higgins, S.C.; Muñoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A Small-Molecule Inhibitor of the NLRP3 Inflammasome for the Treatment of Inflammatory Diseases. Nat. Med. 2015, 21, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Østergaard, J.A.; Jha, J.C.; Sharma, A.; Dai, A.; Choi, J.S.Y.; de Haan, J.B.; Cooper, M.E.; Jandeleit-Dahm, K. Adverse Renal Effects of NLRP3 Inflammasome Inhibition by MCC950 in an Interventional Model of Diabetic Kidney Disease. Clin. Sci. 2022, 136, 167–180. [Google Scholar] [CrossRef]
- He, B.; Nohara, K.; Park, N.; Park, Y.S.; Guillory, B.; Zhao, Z.; Garcia, J.M.; Koike, N.; Lee, C.C.; Takahashi, J.S.; et al. The Small Molecule Nobiletin Targets the Molecular Oscillator to Enhance Circadian Rhythms and Protect against Metabolic Syndrome. Cell Metab. 2016, 23, 610–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.W.; Lan, T.; Sheng, H.; Zheng, F.; Lei, M.K.; Wang, L.X.; Chen, H.F.; Xu, C.Y.; Zhang, F. Nobiletin Alleviates Non-alcoholic Steatohepatitis in MCD-Induced Mice by Regulating Macrophage Polarization. Front. Physiol. 2021, 12, 687744. [Google Scholar] [CrossRef] [PubMed]
- Sudhakaran, M.; Doseff, A.I. The Targeted Impact of Flavones on Obesity-Induced Inflammation and the Potential Synergistic Role in Cancer and the Gut Microbiota. Molecules 2020, 25, 2477. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, R.; Huang, X.; Chen, C.; Dou, D.; Wang, Q.; Wu, X.; Liu, H.; Sun, T. Chrysin Targets Myeloid-Derived Suppressor Cells and Enhances Tumour Response to Anti-PD-1 Immunotherapy. Clin. Transl. Med. 2022, 12, e1019. [Google Scholar] [CrossRef]
- Singh, S.; Meena, A.; Luqman, S. Acacetin and Pinostrobin as a Promising Inhibitor of Cancer-Associated Protein Kinases. Food Chem. Toxicol. 2021, 151, 112091. [Google Scholar] [CrossRef]
- Arango, D.; Morohashi, K.; Yilmaz, A.; Kuramochi, K.; Parihar, A.; Brahimaj, B.; Grotewold, E.; Doseff, A.I. Molecular Basis for the Action of a Dietary Flavonoid Revealed by the Comprehensive Identification of Apigenin Human Targets. Proc. Natl. Acad. Sci. USA 2013, 110, E2153–E2162. [Google Scholar] [CrossRef] [Green Version]
- Sudhakaran, M.; Parra, M.R.; Stoub, H.; Gallo, K.A.; Doseff, A.I. Apigenin by Targeting Hnrnpa2 Sensitizes Triple-Negative Breast Cancer Spheroids to Doxorubicin-Induced Apoptosis and Regulates Expression of ABCC4 and ABCG2 Drug Efflux Transporters. Biochem. Pharmacol. 2020, 182, 114259. [Google Scholar] [CrossRef]
- Voss, O.H.; Arango, D.; Tossey, J.C.; Villalona Calero, M.A.; Doseff, A.I. Splicing Reprogramming of TRAIL/DISC-Components Sensitizes Lung Cancer Cells to TRAIL-Mediated Apoptosis. Cell Death Dis. 2021, 12, 287. [Google Scholar] [CrossRef]
- Yahaya, M.A.F.; Bakar, A.R.A.; Stanslas, J.; Nordin, N.; Zainol, M.; Mehat, M.Z. Insights from Molecular Docking and Molecular Dynamics on the Potential of Vitexin as an Antagonist Candidate against Lipopolysaccharide (LPS) for Microglial Activation in Neuroinflammation. BMC Biotechnol. 2021, 21, 38. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Ali, M.C.; Ruma, R.A.; Mahmud, S.; Paul, G.K.; Saleh, M.A.; Alshahrani, M.M.; Obaidullah, A.J.; Biswas, S.K.; Rahman, M.M.; et al. Molecular Docking and Dynamics Simulation of Natural Compounds from Betel Leaves (Piper betle L.) for Investigating the Potential Inhibition of Alpha-Amylase and Alpha-Glucosidase of Type 2 Diabetes. Molecules 2022, 27, 4526. [Google Scholar] [CrossRef] [PubMed]
- Baruah, I.; Kashyap, C.; Guha, A.K.; Borgohain, G. Insights into the Interaction between Polyphenols and β-Lactoglobulin through Molecular Docking, MD Simulation, and QM/MM Approaches. ACS Omega 2022, 7, 23083–23095. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.S.; Lima, D.R.; Bezerra, L.L.; Monteiro, N.K.V.; Loiola, O.D.; Silva, M.G.V. Virtual Screening of Flavonoids from Chamaecrista Genus: ADME and Pharmacokinetic Properties, Interactions of Flavonoid-DNA complex by Molecular Docking and Molecular Dynamics. J. Biomol. Struct. Dyn. 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]




| Target | Flavone | Dose | Activity | Model System | Ref. |
|---|---|---|---|---|---|
| 10, 30, 50 mg/kg/day i.p. | Systemic TNFα, IL-6, IL-1β, IL-12 ↓ * | C57Bl/6j male mice fed with HFD | [194] | ||
| 25 mg/kg/day i.p. | TNFα, IL-6, IFNγ, MCP-1 ↓ | Pancreatic cancer in C57Bl/6j mice | [195] | ||
| Apigenin | 300 mg/kg/day i.p. | Nuclear NF-κB ↓ | Human hepatocellular carcinoma xenografts in Balb/c nu/nu mice | [196] | |
| 20 and 50 μg/mouse/day p.o. | Phospho-IκBα, IKKα, NF-κB p65 and p50, COX-2 ↓ | Prostate cancer in C57Bl/6j mice | [197] | ||
| 40 μM | IKBKε, IL-1α, IL-6, MCP-1, GM-CSF ↓ | Human MDA-MB-231 breast cancer cells | [198] | ||
| Acacetin | 3–100 μM | Phospho-IκB, nuclear p65, COX-2, TNFα, IL-6, MCP-1 ↓ | Murine RAW264.7 macrophages cultured with 3T3-L1 adipocytes | [199] | |
| Baicalin | 50 mg/kg/day i.p. | TNFα, MCP-1 mRNA ↓ | WAT from HFD-fed C57Bl/6j male mice | [200] | |
| Baicalein | 200, 400 mg/kg/day p.o. | NF-κB ↓ | Murine melanoma B16-F10 xenografts in C57Bl/6j mice | [201] | |
| NF-κB | Chrysin | 25, 30 mg/kg/day i.p. | Systemic TNFα, IL-1β ↓ | C57Bl/6j male mice fed with HFD | [202] |
| Luteolin | 0.005% w/w in HFD chow | Systemic TNFα, IL-1β, IL-6, MIP-1β ↓ | C57Bl/6j male mice fed with HFD | [203] | |
| 0.005% w/w in HFD chow | TNFα, IL-6, MCP-1 ↓ | WAT from OVXed C57Bl/6j female mice fed with HFD * | [180] | ||
| Nobiletin | 10–100 μM | TNFα, MCP-1 ↓ | Murine RAW264.7 macrophages cultured with 3T3-L1 adipocytes | [204] | |
| Tangeretin | 20 mg/kg/day p.o. | TNFα, IL-6, IL-1β, MCP-1 ↓ | WAT from HFD-fed C57Bl/6j male mice | [205] | |
| 20, 40 μM | TNFα, IL-6, IL-1β, MCP-1 ↓ | Murine bone marrow-derived macrophages cultured with adipocytes | [205] | ||
| Vitexin | 30, 60 mg/kg/day p.o. | Phospho-IκB, NF-κB p65, TNFα, IL-6, IL-1β ↓ | C57Bl/6j male mice fed with HFD | [47] | |
| 40, 160 mg/kg/day p.o. | Phospho-IκBα, nuclear p65, TNFα ↑ | M1 macrophages from colon cancers in Balb/c mice | [206] | ||
| STAT | Apigenin | 5–60 μM | Phospho-STAT1, IFNγ- induced PD-L1 expression ↓ | Human melanoma A375, A2058, RPMI-7951 cell lines | [207] |
| 10–50 μM | Phospho-STAT1 and STAT3, PD-L1 expression ↓ | Human NSCLC H460, H538, A549 cell lines | [208] | ||
| Luteolin | 50–100 μM | Phospho-JAK1 and STAT1, IL-8 ↓ | Human colon cancer HT-29 cells | [209] | |
| 0.3–10 μM | IL-6-induced STAT3 phosphorylation ↓ | Human cholangiosarcoma KKU-M156 cells | [210] | ||
| Nrf2 | Apigenin | 1.56–6.25 μM | ARE-luciferase reporter, Nrf2-dependent gene HO-1 ↑ | Human hepatocellular carcinoma HepG2 cells | [211] |
| Nobiletin | 10–100 μM | Nrf2-dependent gene HO-1 ↑ | Murine RAW264.7 macrophages cultured with 3T3-L1 adipocytes | [204] | |
| Scutellarin | 25, 50, 100 mg/kg/day p.o. | Nrf2 ↑ GSK, IL-1β, IL-2 ↓ | Db/db diabetic mice | [212] | |
| Tangeretin | 20–60 μM | Nrf2 ↓ Overcomes drug resistance | Human lung cancer A549/T cells | [213] | |
| Wogonin | 20–60 μM | Nrf2 ↓ Overcomes drug resistance | Human breast cancer MCF7 cells | [214] | |
| NLRP3 | 0.005% w/w in HFD chow | NLRP3, caspase-1, IL-1β ↓ | WAT from OVXed C57Bl/6j female mice fed with HFD * | [215] | |
| Luteolin | 50–150 μM | NLRP3, caspase-1, IL-1β ↑ | Human colon cancer HT-29 cells | [216] | |
| 50 mg/kg/day i.p. | NLRP3, caspase-1, IL-1β ↑ | Human colon cancer cells xenografts in Balb/c nude mice | [216] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kariagina, A.; Doseff, A.I. Anti-Inflammatory Mechanisms of Dietary Flavones: Tapping into Nature to Control Chronic Inflammation in Obesity and Cancer. Int. J. Mol. Sci. 2022, 23, 15753. https://doi.org/10.3390/ijms232415753
Kariagina A, Doseff AI. Anti-Inflammatory Mechanisms of Dietary Flavones: Tapping into Nature to Control Chronic Inflammation in Obesity and Cancer. International Journal of Molecular Sciences. 2022; 23(24):15753. https://doi.org/10.3390/ijms232415753
Chicago/Turabian StyleKariagina, Anastasia, and Andrea I. Doseff. 2022. "Anti-Inflammatory Mechanisms of Dietary Flavones: Tapping into Nature to Control Chronic Inflammation in Obesity and Cancer" International Journal of Molecular Sciences 23, no. 24: 15753. https://doi.org/10.3390/ijms232415753