
The Sperm Small RNA Transcriptome: Implications beyond Reproductive Disorder

 ,
,
Abstract
:
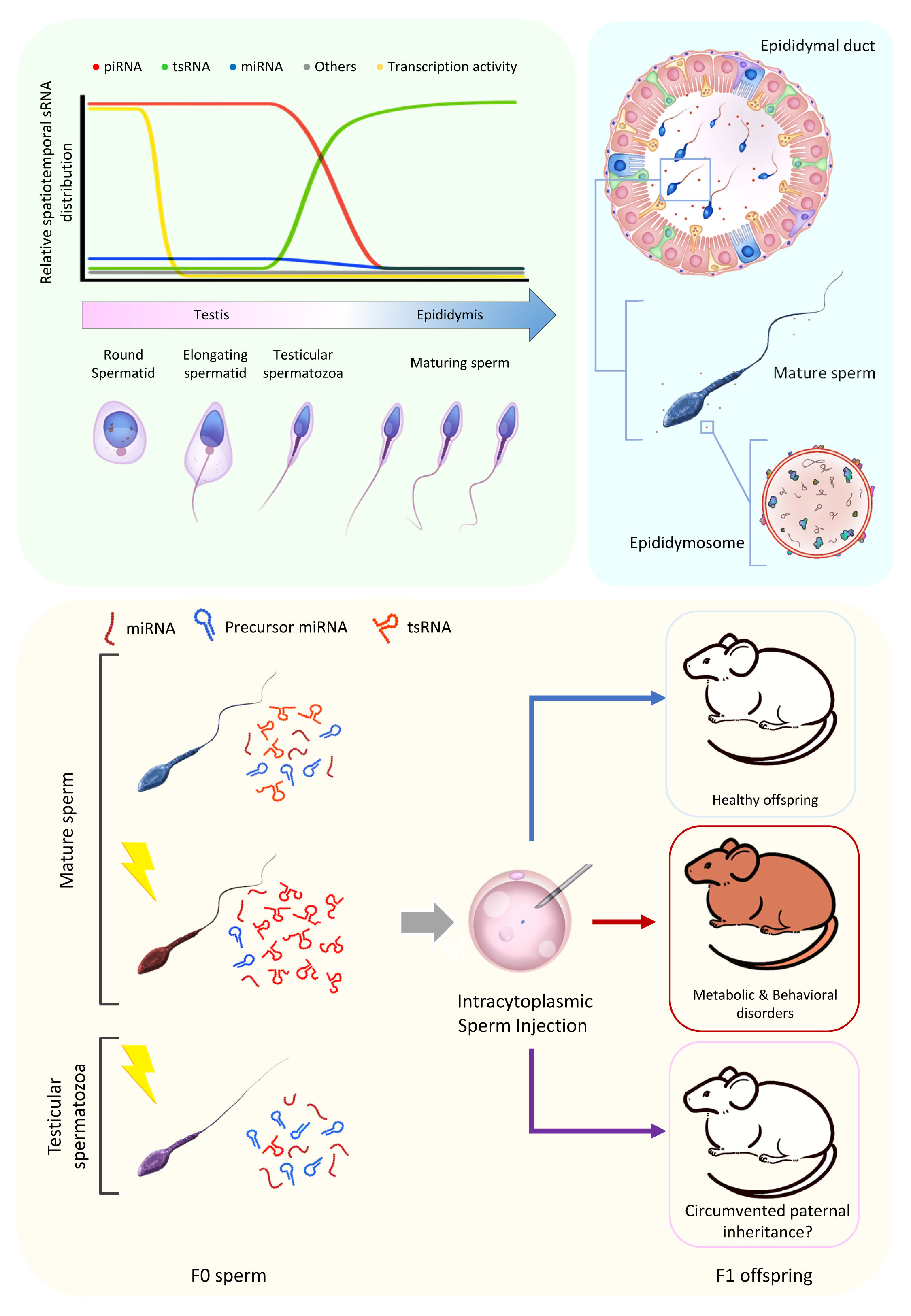{kind=link}
1. Introduction
2. Establishment of the Sperm Epigenome
3. Sperm sncRNA Transcriptome and Fecundity/Fertility
4. Alteration of the Sperm sncRNA Transcriptome in Response to Environmental Insults
4.1. Alteration of Sperm sncRNA in Chronic Stress
4.2. Alteration of Sperm sncRNA in Obesity
4.3. Other Inheritable Traits Involving Sperm sncRNAs
5. Origin of Altered Sperm sncRNA Payload
6. Implications for Assisted Reproductive Technology
7. Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Russell, L.D.; Ettlin, R.A.; Hikim, A.P.S.; Clegg, E.D. Histological and Histopathological Evaluation of the Testis. Int. J. Androl. 1993, 16, 83. [Google Scholar] [CrossRef]
- Yoshida, S.; Sukeno, M.; Nabeshima, Y.-I. A Vasculature-Associated Niche for Undifferentiated Spermatogonia in the Mouse Testis. Science 2007, 317, 1722–1726. [Google Scholar] [CrossRef] [PubMed]
- Handel, M.A.; Schimenti, J.C. Genetics of mammalian meiosis: Regulation, dynamics and impact on fertility. Nat. Rev. Genet. 2010, 11, 124–136. [Google Scholar] [CrossRef]
- Sassone-Corsi, P. Unique Chromatin Remodeling and Transcriptional Regulation in Spermatogenesis. Science 2002, 296, 2176–2178. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, L. Mechanisms of spermiogenesis and spermiation and how they are disturbed. Spermatogenesis 2014, 4, e979623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breucker, H.; Holstein, A.-F. Morphogenesis and fate of the residual body in human spermiogenesis. Cell Tissue Res. 1985, 240, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.S.; Coffey, D.S. DNA Packaging and Organization in Mammalian Spermatozoa: Comparison with Somatic Cell. Biol. Reprod. 1991, 44, 569–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siklenka, K.; Erkek, S.; Godmann, M.; Lambrot, R.; McGraw, S.; Lafleur, C.; Cohen, T.; Xia, J.; Suderman, M.; Hallett, M.; et al. Disruption of histone methylation in developing sperm impairs offspring health transgenerationally. Science 2015, 350, aab2006. [Google Scholar] [CrossRef]
- Wang, T.; Gao, H.; Li, W.; Liu, C. Essential Role of Histone Replacement and Modifications in Male Fertility. Front. Genet. 2019, 10, 962. [Google Scholar] [CrossRef]
- Miller, D.; Brinkworth, M.; Iles, D. Paternal DNA packaging in spermatozoa: More than the sum of its parts? DNA, histones, protamines and epigenetics. Reproduction 2010, 139, 287–301. [Google Scholar] [CrossRef]
- Hammoud, S.S.; Nix, D.A.; Zhang, H.; Purwar, J.; Carrell, D.T.; Cairns, B.R. Distinctive chromatin in human sperm packages genes for embryo development. Nature 2009, 460, 473–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Muratani, M.; Araki, H.; Miura, F.; Suzuki, T.; Dohmae, N.; Katou, Y.; Shirahige, K.; Ito, T.; Ishii, S. Mapping of histone-binding sites in histone replacement-completed spermatozoa. Nat. Commun. 2018, 9, 3885. [Google Scholar] [CrossRef] [PubMed]
- Ajduk, A.; Yamauchi, Y.; Ward, M.A. Sperm Chromatin Remodeling after Intracytoplasmic Sperm Injection Differs from That of In Vitro Fertilization1. Biol. Reprod. 2006, 75, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heijden, G.; Derijck, A.; Ramos, L.; Giele, M.; van der Vlag, J.; de Boer, P. Transmission of modified nucleosomes from the mouse male germline to the zygote and subsequent remodeling of paternal chromatin. Dev. Biol. 2006, 298, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heijden, G.W.; Ramos, L.; Baart, E.B.; Berg, I.M.V.D.; Derijck, A.A.; van der Vlag, J.; Martini, E.; de Boer, P. Sperm-derived histones contribute to zygotic chromatin in humans. BMC Dev. Biol. 2008, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Bao, J.; Bedford, M.T. Epigenetic regulation of the histone-to-protamine transition during spermiogenesis. Reproduction 2016, 151, R55–R70. [Google Scholar] [CrossRef] [Green Version]
- Ben Maamar, M.; Sadler-Riggleman, I.; Beck, D.; Skinner, M.K. Epigenetic Transgenerational Inheritance of Altered Sperm Histone Retention Sites. Sci. Rep. 2018, 8, 5308. [Google Scholar] [CrossRef] [Green Version]
- Galan, C.; Serra, R.W.; Sun, F.; Rinaldi, V.D.; Conine, C.C.; Rando, O.J. Stability of the cytosine methylome during post-testicular sperm maturation in mouse. PLoS Genet. 2021, 17, e1009416. [Google Scholar] [CrossRef]
- Bedi, Y.S.; Roach, A.N.; Thomas, K.N.; Mehta, N.A.; Golding, M.C. Chromatin alterations during the epididymal maturation of mouse sperm refine the paternally inherited epigenome. Epigenet. Chromatin 2022, 15, 2. [Google Scholar] [CrossRef]
- Cappallo-Obermann, H.; Schulze, W.; Jastrow, H.; Baukloh, V.; Spiess, A.-N. Highly purified spermatozoal RNA obtained by a novel method indicates an unusual 28S/18S rRNA ratio and suggests impaired ribosome assembly. Mol. Hum. Reprod. 2011, 17, 669–678. [Google Scholar] [CrossRef]
- Pessot, C.A.; Brito, M.; Figueroa, J.; Concha, I.I.; Yañez, A.; Burzio, L.O. Presence of RNA in the sperm nucleus. Biochem. Biophys. Res. Commun. 1989, 158, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Briggs, D.; Snowden, H.; Hamlington, J.; Rollinson, S.; Lilford, R.; Krawetz, S.A. A complex population of RNAs exists in human ejaculate spermatozoa: Implications for understanding molecular aspects of spermiogenesis. Gene 1999, 237, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Miller, D. Analysis and significance of messenger RNA in human ejaculated spermatozoa. Mol. Reprod. Dev. 2000, 56, 259–264. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Dix, D.J.; Miller, D.; Khatri, P.; Krawetz, S.A. Spermatozoal RNA profiles of normal fertile men. Lancet 2002, 360, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Ostermeier, G.C.; Goodrich, R.J.; Moldenhauer, J.S.; Diamond, M.; Krawetz, S.A. A Suite of Novel Human Spermatozoal RNAs. J. Androl. 2005, 26, 70–74. [Google Scholar] [PubMed]
- Krawetz, S.A.; Kruger, A.; Lalancette, C.; Tagett, R.; Anton, E.; Draghici, S.; Diamond, M.P. A survey of small RNAs in human sperm. Hum. Reprod. 2011, 26, 3401–3412. [Google Scholar] [CrossRef] [Green Version]
- Ostermeier, G.C.; Miller, D.; Huntriss, J.D.; Diamond, M.P.; Krawetz, S.A. Delivering spermatozoan RNA to the oocyte. Nature 2004, 429, 154. [Google Scholar] [CrossRef]
- Liu, W.-M.; Pang, R.T.K.; Chiu, P.C.N.; Wong, B.P.C.; Lao, K.; Lee, K.-F.; Yeung, W.S.B. Sperm-borne microRNA-34c is required for the first cleavage division in mouse. Proc. Natl. Acad. Sci. USA 2011, 109, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Ntostis, P.; Carter, D.; Iles, D.; Huntriss, J.; Tzetis, M.; Miller, D. Potential sperm contributions to the murine zygote predicted by in silico analysis. Reproduction 2017, 154, 777–788. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Chao, S.-B.; Xiao, L.; Wang, Z.-B.; Meng, T.-G.; Li, Y.-Y.; Han, Z.-M.; Ouyang, Y.-C.; Hou, Y.; Sun, Q.-Y.; et al. Sperm-carried RNAs play critical roles in mouse embryonic development. Oncotarget 2017, 8, 67394–67405. [Google Scholar] [CrossRef]
- Conine, C.; Sun, F.; Song, L.; Rivera-Pérez, J.A.; Rando, O.J. Small RNAs Gained during Epididymal Transit of Sperm Are Essential for Embryonic Development in Mice. Dev. Cell 2018, 46, 470–480.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, N.; Strillacci, M.G.; Peñagaricano, F.; Khatib, H. Characterization and functional roles of paternal RNAs in 2–4 cell bovine embryos. Sci. Rep. 2019, 9, 20347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral-Vazquez, C.; Blanco, J.; Cigliano, R.A.; Sarrate, Z.; Rivera-Egea, R.; Vidal, F.; Garrido, N.; Daub, C.; Anton, E. The RNA content of human sperm reflects prior events in spermatogenesis and potential post-fertilization effects. Mol. Hum. Reprod. 2021, 27, gaab035. [Google Scholar] [CrossRef]
- Sharma, U.; Sun, F.; Conine, C.C.; Reichholf, B.; Kukreja, S.; Herzog, V.A.; Ameres, S.L.; Rando, O.J. Small RNAs Are Trafficked from the Epididymis to Developing Mammalian Sperm. Dev. Cell 2018, 46, 481–494.e6. [Google Scholar] [CrossRef] [PubMed]
- Nixon, B.; Stanger, S.J.; Mihalas, B.P.; Reilly, J.N.; Anderson, A.L.; Tyagi, S.; Holt, J.E.; McLaughlin, E.A. The MicroRNA Signature of Mouse Spermatozoa Is Substantially Modified during Epididymal Maturation1. Biol. Reprod. 2015, 93, 91. [Google Scholar] [CrossRef] [PubMed]
- Fuster, C.D.; Farrell, D.; Stern, F.A.; Hecht, N.B. RNA polymerase activity in bovine spermatozoa. J. Cell Biol. 1977, 74, 698–706. [Google Scholar] [CrossRef]
- Wilkerson, D.C.; Sarge, K.D. RNA polymerase II interacts with the Hspa1b promoter in mouse epididymal spermatozoa. Reproduction 2009, 137, 923–929. [Google Scholar] [CrossRef] [Green Version]
- Yanagimachi, R.; Kamiguchi, Y.; Mikamo, K.; Suzuki, F. Maturation of spermatozoa in the epididymis of the Chinese hamster. Am. J. Anat. 1985, 172, 317–330. [Google Scholar] [CrossRef]
- Păunescu, T.G.; Shum, W.W.; Huynh, C.; Lechner, L.; Goetze, B.; Brown, D.; Breton, S.; Unescu, T.G.P. High-resolution helium ion microscopy of epididymal epithelial cells and their interaction with spermatozoa. Mol. Hum. Reprod. 2014, 20, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, R.; Saez, F.; Girouard, J.; Frenette, G. Role of exosomes in sperm maturation during the transit along the male reproductive tract. Blood Cells, Mol. Dis. 2005, 35, 1–10. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Hammadeh, M.; Schmitt, J.; Leidinger, P.; Keller, A.; Meese, E.; Backes, C. Altered microRNA expression profiles of human spermatozoa in patients with different spermatogenic impairments. Fertil. Steril. 2013, 99, 1249–1255.e16. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Hammadeh, M.; Backes, C.; Fischer, U.; Leidinger, P.; Lubbad, A.M.; Keller, A.; Meese, E. Panel of five microRNAs as potential biomarkers for the diagnosis and assessment of male infertility. Fertil. Steril. 2014, 102, 989–997.e1. [Google Scholar] [CrossRef] [PubMed]
- Salas-Huetos, A.; Blanco, J.; Vidal, F.; Godo, A.; Grossmann, M.; Pons, M.C.; F-Fernández, S.; Garrido, N.; Anton, E. Spermatozoa from patients with seminal alterations exhibit a differential micro-ribonucleic acid profile. Fertil. Steril. 2015, 104, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, X.; Mata, A.; Bassas, L.; Larriba, S. Altered miRNA Signature of Developing Germ-cells in Infertile Patients Relates to the Severity of Spermatogenic Failure and Persists in Spermatozoa. Sci. Rep. 2015, 5, 17991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.; Li, D.; Wang, L.; Wu, J.; Hu, Y.; Wang, Z.; Chen, Y.; Cao, X.; Jiang, C.; Yan, W.; et al. MicroRNA-449 and MicroRNA-34b/c Function Redundantly in Murine Testes by Targeting E2F Transcription Factor-Retinoblastoma Protein (E2F-pRb) Pathway. J. Biol. Chem. 2012, 287, 21686–21698. [Google Scholar] [CrossRef] [Green Version]
- Bouhallier, F.; Allioli, N.; Lavial, F.; Chalmel, F.; Perrard, M.-H.; Durand, P.; Samarut, J.; Pain, B.; Rouault, J.-P. Role of miR-34c microRNA in the late steps of spermatogenesis. Rna 2010, 16, 720–731. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Johnson, J.; Chellappan, S. Small molecule regulators of Rb–E2F pathway as modulators of transcription. Biochim. et Biophys. Acta 2010, 1799, 788–794. [Google Scholar] [CrossRef] [Green Version]
- Comazzetto, S.; Di Giacomo, M.; Rasmussen, K.D.; Much, C.; Azzi, C.; Perlas, E.; Morgan, M.; O’Carroll, D. Oligoasthenoteratozoospermia and Infertility in Mice Deficient for miR-34b/c and miR-449 Loci. PLOS Genet. 2014, 10, e1004597. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Tang, C.; Zhang, Y.; Wu, J.; Bao, J.; Zheng, H.; Xu, C.; Yan, W. mir-34b/c and mir-449a/b/c are required for spermatogenesis, but not for the first cleavage division in mice. Biol. Open 2015, 4, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Gan, H.; Lin, X.; Zhang, Z.; Zhang, W.; Liao, S.; Wang, L.; Han, C. piRNA profiling during specific stages of mouse spermatogenesis. Rna 2011, 17, 1191–1203. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, Q.C.W.; Duan, E. Epigenetic inheritance of acquired traits through sperm RNAs and sperm RNA modifications. Nat. Rev. Genet. 2016, 17, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, I.; Skvortsova, Y.; Kondratieva, S.; Funikov, S.; Azhikina, T. Two modes of targeting transposable elements by piRNA pathway in human testis. Rna 2017, 23, 1614–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czech, B.; Munafò, M.; Ciabrelli, F.; Eastwood, E.L.; Fabry, M.H.; Kneuss, E.; Hannon, G.J. piRNA-Guided Genome Defense: From Biogenesis to Silencing. Annu. Rev. Genet. 2018, 52, 131–157. [Google Scholar] [CrossRef]
- Chuma, S.; Nakano, T. piRNA and spermatogenesis in mice. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20110338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.P.; Kotaja, N. Small RNAs in spermatogenesis. Mol. Cell. Endocrinol. 2014, 382, 498–508. [Google Scholar] [CrossRef]
- Choi, H.; Wang, Z.; Dean, J. Sperm acrosome overgrowth and infertility in mice lacking chromosome 18 pachytene piRNA. PLoS Genet. 2021, 17, e1009485. [Google Scholar] [CrossRef]
- Wu, P.-H.; Fu, Y.; Cecchini, K.; Özata, D.M.; Arif, A.; Yu, T.; Colpan, C.; Gainetdinov, I.; Weng, Z.; Zamore, P.D. The evolutionarily conserved piRNA-producing locus pi6 is required for male mouse fertility. Nat. Genet. 2020, 52, 728–739. [Google Scholar] [CrossRef]
- He, L.; Wu, X.; Wu, R.; Guo, P.; He, W.; Sun, W.; Chen, H. Seminal plasma piRNA array analysis and identification of possible biomarker piRNAs for the diagnosis of asthenozoospermia. Exp. Ther. Med. 2022, 23, 347. [Google Scholar] [CrossRef]
- Jodar, M.; Kalko, S.; Castillo, J.; Ballescà, J.L.; Oliva, R. Differential RNAs in the sperm cells of asthenozoospermic patients. Hum. Reprod. 2012, 27, 1431–1438. [Google Scholar] [CrossRef]
- Dougherty, G.W.; Mizuno, K.; Nöthe-Menchen, T.; Ikawa, Y.; Boldt, K.; Ta-Shma, A.; Aprea, I.; Minegishi, K.; Pang, Y.-P.; Pennekamp, P.; et al. CFAP45 deficiency causes situs abnormalities and asthenospermia by disrupting an axonemal adenine nucleotide homeostasis module. Nat. Commun. 2020, 11, 5520. [Google Scholar] [CrossRef]
- Hong, Y.; Wu, Y.; Zhang, J.; Yu, C.; Shen, L.; Chen, H.; Chen, L.; Zhou, X.; Gao, F. Decreased piRNAs in Infertile Semen Are Related to Downregulation of Sperm MitoPLD Expression. Front. Endocrinol. 2021, 12, 696121. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Fang, L.; Shi, B.; Qiu, S.; Ye, Y. Spermatozoa Expression of piR-31704, piR-39888, and piR-40349 and Their Correlation to Sperm Concentration and Fertilization Rate after ICSI. Reprod. Sci. 2018, 25, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cao, J.; Wang, X.; Guo, C.; Liu, Y.; Wang, T. Deciphering the tRNA-derived small RNAs: Origin, development, and future. Cell Death Dis. 2021, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.; Raghavan, S.; DasGupta, R.; Palakodeti, D. tRNA-derived fragments (tRFs): Establishing their turf in post-transcriptional gene regulation. Cell. Mol. Life Sci. 2021, 78, 2607–2619. [Google Scholar] [CrossRef] [PubMed]
- Magee, R.; Rigoutsos, I. On the expanding roles of tRNA fragments in modulating cell behavior. Nucleic Acids Res. 2020, 48, 9433–9448. [Google Scholar] [CrossRef]
- Shen, Y.; Yu, X.; Zhu, L.; Li, T.; Yan, Z.; Guo, J. Transfer RNA-derived fragments and tRNA halves: Biogenesis, biological functions and their roles in diseases. J. Mol. Med. 2018, 96, 1167–1176. [Google Scholar] [CrossRef]
- Peng, H.; Shi, J.; Zhang, Y.; Zhang, H.; Liao, S.; Li, W.; Lei, L.; Han, C.; Ning, L.; Cao, Y.; et al. A novel class of tRNA-derived small RNAs extremely enriched in mature mouse sperm. Cell Res. 2012, 22, 1609–1612. [Google Scholar] [CrossRef]
- Sharma, U.; Conine, C.C.; Shea, J.M.; Boskovic, A.; Derr, A.G.; Bing, X.Y.; Belleannee, C.; Kucukural, A.; Serra, R.W.; Sun, F.; et al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science 2016, 351, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zheng, Y.; Lei, A.; Zhang, H.; Niu, H.; Li, X.; Zhang, P.; Liao, M.; Lv, Y.; Zhu, Z.; et al. Early cleavage of preimplantation embryos is regulated by tRNAGln-TTG–derived small RNAs present in mature spermatozoa. J. Biol. Chem. 2020, 295, 10885–10900. [Google Scholar] [CrossRef]
- Chen, X.; Sun, Q.; Zheng, Y.; Liu, Z.; Meng, X.; Zeng, W.; Lu, H. Human sperm tsRNA as potential biomarker and therapy target for male fertility. Reproduction 2021, 161, 111–122. [Google Scholar] [CrossRef]
- Hua, M.; Liu, W.; Chen, Y.; Zhang, F.; Xu, B.; Liu, S.; Chen, G.; Shi, H.; Wu, L. Identification of small non-coding RNAs as sperm quality biomarkers for in vitro fertilization. Cell Discov. 2019, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, M.F.; Lehner, B. Intergenerational and transgenerational epigenetic inheritance in animals. Nature 2019, 21, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; Holmes, A. The ascent of mouse: Advances in modelling human depression and anxiety. Nat. Rev. Drug Discov. 2005, 4, 775–790. [Google Scholar] [CrossRef]
- Blewitt, M.; Whitelaw, E. The Use of Mouse Models to Study Epigenetics. Cold Spring Harb. Perspect. Biol. 2013, 5, a017939. [Google Scholar] [CrossRef]
- Hanna, C.W.; Demond, H.; Kelsey, G. Epigenetic regulation in development: Is the mouse a good model for the human? Hum. Reprod. Updat. 2018, 24, 556–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapp, K.; Jawaid, A.; Sarkies, P.; Bohacek, J.; Pelczar, P.; Prados, J.; Farinelli, L.; Miska, E.; Mansuy, I.M. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 2014, 17, 667–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, A.B.; Morgan, C.P.; Bronson, S.L.; Revello, S.; Bale, T.L. Paternal Stress Exposure Alters Sperm MicroRNA Content and Reprograms Offspring HPA Stress Axis Regulation. J. Neurosci. 2013, 33, 9003–9012. [Google Scholar] [CrossRef] [Green Version]
- Saavedra-Rodríguez, L.; Feig, L.A. Chronic Social Instability Induces Anxiety and Defective Social Interactions across Generations. Biol. Psychiatry 2012, 73, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Dickson, D.A.; Paulus, J.K.; Mensah, V.; Lem, J.; Saavedra-Rodriguez, L.; Gentry, A.; Pagidas, K.; Feig, L.A. Reduced levels of miRNAs 449 and 34 in sperm of mice and men exposed to early life stress. Transl. Psychiatry 2018, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Short, A.; Fennell, K.A.; Perreau, V.M.; Fox, A.; O’Bryan, M.K.; Kim, J.H.; Bredy, T.W.; Pang, T.; Hannan, A. Elevated paternal glucocorticoid exposure alters the small noncoding RNA profile in sperm and modifies anxiety and depressive phenotypes in the offspring. Transl. Psychiatry 2016, 6, e837. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Z.-P.; Hu, H.; Lei, J.; Zhou, Z.; Yao, B.; Chen, L.; Liang, G.; Zhan, S.; Zhu, X.; et al. Sperm microRNAs confer depression susceptibility to offspring. Sci. Adv. 2021, 7, eabd7605. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, A.B.; Morgan, C.P.; Leu, N.A.; Bale, T.L. Transgenerational epigenetic programming via sperm microRNA recapitulates effects of paternal stress. Proc. Natl. Acad. Sci. USA 2015, 112, 13699–13704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaisvaser, S.; Modai, S.; Farberov, L.; Lin, T.; Sharon, H.; Gilam, A.; Volk, N.; Admon, R.; Edry, L.; Fruchter, E.; et al. Neuro-Epigenetic Indications of Acute Stress Response in Humans: The Case of MicroRNA-29c. PLoS ONE 2016, 11, e0146236. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, D.P.; Eriksen, M.B.; Rajalingam, D.; Nymoen, I.; Nielsen, M.B.; Einarsen, S.; Gjerstad, J. Exposure to workplace bullying, microRNAs and pain; evidence of a moderating effect of miR-30c rs928508 and miR-223 rs3848900. Stress 2019, 23, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.P.; Shetty, A.C.; Chan, J.C.; Berger, D.S.; Ament, S.A.; Epperson, C.N.; Bale, T.L. Repeated sampling facilitates within- and between-subject modeling of the human sperm transcriptome to identify dynamic and stress-responsive sncRNAs. Sci. Rep. 2020, 10, 17498. [Google Scholar] [CrossRef]
- Hoyer, C.; Richter, H.; Brandwein, C.; Riva, M.A.; Gass, P. Preconceptional paternal exposure to a single traumatic event affects postnatal growth of female but not male offspring. NeuroReport 2013, 24, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Oakberg, E.F. Duration of spermatogenesis in the mouse and timing of stages of the cycle of the seminiferous epithelium. Am. J. Anat. 1956, 99, 507–516. [Google Scholar] [CrossRef]
- Chaudhuri, G.R.; Das, A.; Kesh, S.B.; Bhattacharya, K.; Dutta, S.; Sengupta, P.; Syamal, A.K. Obesity and male infertility: Multifaceted reproductive disruption. Middle East Fertil. Soc. J. 2022, 27, 8. [Google Scholar] [CrossRef]
- Broughton, D.E.; Moley, K.H. Obesity and female infertility: Potential mediators of obesity’s impact. Fertil. Steril. 2017, 107, 840–847. [Google Scholar] [CrossRef] [Green Version]
- Fullston, T.; Teague, E.M.C.O.; Palmer, N.O.; DeBlasio, M.J.; Mitchell, M.; Corbett, M.; Print, C.G.; Owens, J.A.; Lane, M. Paternal obesity initiates metabolic disturbances in two generations of mice with incomplete penetrance to the F 2 generation and alters the transcriptional profile of testis and sperm microRNA content. FASEB J. 2013, 27, 4226–4243. [Google Scholar] [CrossRef]
- Fullston, T.; Ohlsson-Teague, E.M.C.; Print, C.G.; Sandeman, L.Y.; Lane, M. Sperm microRNA Content Is Altered in a Mouse Model of Male Obesity, but the Same Suite of microRNAs Are Not Altered in Offspring’s Sperm. PLoS ONE 2016, 11, e0166076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakos, H.W.; Mitchell, M.; Setchell, B.P.; Lane, M. The effect of paternal diet-induced obesity on sperm function and fertilization in a mouse model. Int. J. Androl. 2010, 34, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.; Bakos, H.W.; Lane, M. Paternal diet-induced obesity impairs embryo development and implantation in the mouse. Fertil. Steril. 2011, 95, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Binder, N.K.; Hannan, N.J.; Gardner, D.K. Paternal Diet-Induced Obesity Retards Early Mouse Embryo Development, Mitochondrial Activity and Pregnancy Health. PLoS ONE 2012, 7, e52304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, S.; Soni, C.B.; Xie, J.; Li, Y.; Zhu, H.; Wu, F.; Zhi, X. MiR-30a-5p accelerates adipogenesis by negatively regulating Sirtuin 1. Int. J. Clin. Exp. Pathol. 2018, 11, 5203–5212. [Google Scholar] [PubMed]
- Saha, P.K.; Hamilton, M.P.; Rajapakshe, K.; Putluri, V.; Felix, J.B.; Masschelin, P.; Cox, A.R.; Bajaj, M.; Putluri, N.; Coarfa, C.; et al. miR-30a targets gene networks that promote browning of human and mouse adipocytes. Am. J. Physiol. Metab. 2020, 319, E667–E677. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.-H.; Peng, H.; Zhang, X.; Zhang, Y.; et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, X.; Shi, J.; Tuorto, F.; Li, X.; Liu, Y.; Liebers, R.; Zhang, L.; Qu, Y.; Qian, J.; et al. Dnmt2 mediates intergenerational transmission of paternally acquired metabolic disorders through sperm small non-coding RNAs. Nature 2018, 20, 535–540. [Google Scholar] [CrossRef] [Green Version]
- Grandjean, V.; Fourré, S.; De Abreu, D.A.F.; Derieppe, M.-A.; Remy, J.-J.; Rassoulzadegan, M. RNA-mediated paternal heredity of diet-induced obesity and metabolic disorders. Sci. Rep. 2015, 5, 18193. [Google Scholar] [CrossRef] [Green Version]
- Donkin, I.; Versteyhe, S.; Ingerslev, L.R.; Qian, K.; Mechta, M.; Nordkap, L.; Mortensen, B.; Appel, E.V.R.; Jørgensen, N.; Kristiansen, V.B.; et al. Obesity and Bariatric Surgery Drive Epigenetic Variation of Spermatozoa in Humans. Cell Metab. 2015, 23, 369–378. [Google Scholar] [CrossRef]
- Pitetti, J.-L.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M.; et al. An Essential Role for Insulin and IGF1 Receptors in Regulating Sertoli Cell Proliferation, Testis Size, and FSH Action in Mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Bedzhov, I.; Liszewska, E.; Kanzler, B.; Stemmler, M.P. Igf1r Signaling Is Indispensable for Preimplantation Development and Is Activated via a Novel Function of E-cadherin. PLoS Genet. 2012, 8, e1002609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortuza, R.; Feng, B.; Chakrabarti, S. miR-195 regulates SIRT1-mediated changes in diabetic retinopathy. Diabetologia 2014, 57, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Adamkova, K.; Yi, Y.-J.; Petr, J.; Zalmanova, T.; Hoskova, K.; Jelinkova, P.; Moravec, J.; Kralickova, M.; Sutovsky, M.; Sutovsky, P.; et al. SIRT1-dependent modulation of methylation and acetylation of histone H3 on lysine 9 (H3K9) in the zygotic pronuclei improves porcine embryo development. J. Anim. Sci. Biotechnol. 2017, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashapkin, V.; Suvorov, A.; Pilsner, J.R.; Krawetz, S.A.; Sergeyev, O. Age-associated epigenetic changes in mammalian sperm: Implications for offspring health and development. Hum. Reprod. Updat. 2022, dmac033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, L.; Sun, X.; Zhang, Z.; Liu, J.; Xin, Y.; Yu, J.; Jia, Y.; Sheng, J.; Hu, G.-F.; et al. Angiogenin mediates paternal inflammation-induced metabolic disorders in offspring through sperm tsRNAs. Nat. Commun. 2021, 12, 6673. [Google Scholar] [CrossRef]
- Rompala, G.R.; Mounier, A.; Wolfe, C.M.; Lin, Q.; Lefterov, I.; Homanics, G.E. Heavy Chronic Intermittent Ethanol Exposure Alters Small Noncoding RNAs in Mouse Sperm and Epididymosomes. Front. Genet. 2018, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Finegersh, A.; Homanics, G.E. Paternal Alcohol Exposure Reduces Alcohol Drinking and Increases Behavioral Sensitivity to Alcohol Selectively in Male Offspring. PLoS ONE 2014, 9, e99078. [Google Scholar] [CrossRef] [Green Version]
- Rompala, G.R.; Finegersh, A.; Slater, M.; Homanics, G.E. Paternal preconception alcohol exposure imparts intergenerational alcohol-related behaviors to male offspring on a pure C57BL/6J background. Alcohol 2017, 60, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Beeler, E.; Nobile, Z.L.; Homanics, G.E. Paternal Preconception Every-Other-Day Ethanol Drinking Alters Behavior and Ethanol Consumption in Offspring. Brain Sci. 2019, 9, 56. [Google Scholar] [CrossRef]
- Bedi, Y.; Chang, R.C.; Gibbs, R.; Clement, T.M.; Golding, M.C. Alterations in sperm-inherited noncoding RNAs associate with late-term fetal growth restriction induced by preconception paternal alcohol use. Reprod. Toxicol. 2019, 87, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.C.; Morgan, C.P.; Leu, N.A.; Shetty, A.; Cisse, Y.M.; Nugent, B.M.; Morrison, K.E.; Jašarević, E.; Huang, W.; Kanyuch, N.; et al. Reproductive tract extracellular vesicles are sufficient to transmit intergenerational stress and program neurodevelopment. Nat. Commun. 2020, 11, 1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshanbayeva, A.; Tanwar, D.K.; Roszkowski, M.; Manuella, F.; Mansuy, I.M. Early life stress affects the miRNA cargo of epididymal extracellular vesicles in mouse. Biol. Reprod. 2021, 105, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Rompala, G.R.; Ferguson, C.; Homanics, G.E. Coincubation of sperm with epididymal extracellular vesicle preparations from chronic intermittent ethanol-treated mice is sufficient to impart anxiety-like and ethanol-induced behaviors to adult progeny. Alcohol 2020, 87, 111–120. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Tan, D.; Zhang, X.; Yan, M.; Zhang, Y.; Franklin, R.; Shahbazi, M.; Mackinlay, K.; Liu, S.; et al. PANDORA-seq expands the repertoire of regulatory small RNAs by overcoming RNA modifications. Nature 2021, 23, 424–436. [Google Scholar] [CrossRef]
- Choy, K.H.K.; Chan, S.Y.; Lam, W.; Jin, J.; Zheng, T.; Law, T.Y.S.; Yu, S.S.; Wang, W.; Li, L.; Xie, G.; et al. The repertoire of testicular extracellular vesicle cargoes and their involvement in inter-compartmental communication associated with spermatogenesis. BMC Biol. 2022, 20, 78. [Google Scholar] [CrossRef]
- de Mouzon, J.; Lancaster, P.; Nygren, K.G.; Sullivan, E.; Zegers-Hochschild, F.; Mansour, R.; Ishihara, O.; Adamson, D. World Collaborative Report on Assisted Reproductive Technology, 2002. Hum. Reprod. 2009, 24, 2310–2320. [Google Scholar] [CrossRef] [Green Version]
- Adamson, G.D. O-043 International committee for monitoring assisted reproductive technologies (ICMART) preliminary world report on ART, 2017. Hum. Reprod. 2021, 36, deab126.042. [Google Scholar] [CrossRef]
- Miyaoka, R.; Esteves, S.C. Predictive factors for sperm retrieval and sperm injection outcomes in obstructive azoospermia: Do etiology, retrieval techniques and gamete source play a role? Clinics 2013, 68, 111–119. [Google Scholar] [CrossRef]
- Coward, R.M.; Mills, J.N. A step-by-step guide to office-based sperm retrieval for obstructive azoospermia. Transl. Androl. Urol. 2017, 6, 730–744. [Google Scholar] [CrossRef]
- Sullivan, R.; Mieusset, R. The human epididymis: Its function in sperm maturation. Hum. Reprod. Updat. 2016, 22, 574–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, C.L.; Chow, S.; Rosenwaks, Z.; Palermo, G.D. Development of ICSI. Reproduction 2018, 156, F51–F58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, B.; Morin, S.; Juneau, C.; Neal, S.; Landis, J.; Scott, R.; Hotaling, J. A comparison of the relative efficiency of ICSI and extended culture with epididymal sperm versus testicular sperm in patients with obstructive azoospermia. Asian J. Androl. 2020, 22, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Palermo, G.D.; Neri, Q.V.; Hariprashad, J.J.; Davis, O.K.; Veeck, L.L.; Rosenwaks, Z. ICSI and Its Outcome. Semin. Reprod. Med. 2000, 18, 161–170. [Google Scholar] [CrossRef]
- Kalsi, J.; Thum, M.Y.; Muneer, A.; Pryor, J.; Abdullah, H.; Minhas, S. Analysis of the outcome of intracytoplasmic sperm injection using fresh or frozen sperm. Br. J. Urol. 2010, 107, 1124–1128. [Google Scholar] [CrossRef]
- Jin, L.; Li, Z.; Gu, L.; Huang, B. Neonatal outcome of children born after ICSI with epididymal or testicular sperm: A 10-year study in China. Sci. Rep. 2020, 10, 5145. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Lu, S.; Yuan, M.; Ma, G.; Li, X.; Zhang, T.; Gao, S.; Wei, D.; Chen, Z.-J.; Liu, H.; et al. Does ICSI outcome in obstructive azoospermia differ according to the origin of retrieved spermatozoa or the cause of epididymal obstruction? A comparative study. Int. Urol. Nephrol. 2022, 54, 3087–3095. [Google Scholar] [CrossRef]
- Shih, K.-W.; Shen, P.-Y.; Wu, C.-C.; Kang, Y.-N. Testicular versus percutaneous epididymal sperm aspiration for patients with obstructive azoospermia: A systematic review and meta-analysis. Transl. Androl. Urol. 2019, 8, 631–640. [Google Scholar] [CrossRef]
- van Wely, M.; Barbey, N.; Meissner, A.; Repping, S.; Silber, S.J. Live birth rates after MESA or TESE in men with obstructive azoospermia: Is there a difference? Hum. Reprod. 2015, 30, 761–766. [Google Scholar] [CrossRef]
- Fedder, J.; Gabrielsen, A.; Humaidan, P.; Erb, K.; Ernst, E.; Loft, A. Malformation rate and sex ratio in 412 children conceived with epididymal or testicular sperm. Hum. Reprod. 2007, 22, 1080–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belva, F.; De Schrijver, F.; Tournaye, H.; Liebaers, I.; Devroey, P.; Haentjens, P.; Bonduelle, M.-L. Neonatal outcome of 724 children born after ICSI using non-ejaculated sperm. Hum. Reprod. 2011, 26, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.B.; Lusignan, M.F.; Son, W.; Sabbah, M.; Buckett, W.; Chan, P. ICSI outcomes using testicular spermatozoa in non-azoospermic couples with recurrent ICSI failure and no previous live births. Andrology 2019, 7, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Fan, D.; Wang, X.; Mao, C.; Chu, Y.; Zhang, H.; Liu, W.; Ding, S.; Liu, Q.; Yuan, M.; et al. ICSI outcomes for infertile men with severe or complete asthenozoospermia. Basic Clin. Androl. 2022, 32, 6. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-N.; Hsiao, Y.-W.; Chen, C.-Y.; Wu, C.-C. Testicular sperm is superior to ejaculated sperm for ICSI in cryptozoospermia: An update systematic review and meta-analysis. Sci. Rep. 2018, 8, 7874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollero, M.; Gil-Guzman, E.; Lopez, M.C.; Sharma, R.; Agarwal, A.; Larson, K.; Evenson, D.; Thomas, A.J.; Alvarez, J.G. Characterization of subsets of human spermatozoa at different stages of maturation: Implications in the diagnosis and treatment of male infertility. Hum. Reprod. 2001, 16, 1912–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arafa, M.; Almalki, A.; Albadr, M.; Burjaq, H.; Majzoub, A.; Alsaid, S.; ElBardisi, H. ICSI outcome in patients with high DNA fragmentation: Testicular versus ejaculated spermatozoa. Andrologia 2017, 50, e12835. [Google Scholar] [CrossRef] [PubMed]
- Awaga, H.A.; Bosdou, J.K.; Goulis, D.G.; Chatzimeletiou, K.; Salem, M.; Roshdy, S.; Grimbizis, G.; Tarlatzis, B.C.; Kolibianakis, E.M. Testicular versus ejaculated spermatozoa for ICSI in patients without azoospermia: A systematic review. Reprod. Biomed. Online 2018, 37, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Esteves, S.C.; Roque, M.; Bradley, C.K.; Garrido, N. Reproductive outcomes of testicular versus ejaculated sperm for intracytoplasmic sperm injection among men with high levels of DNA fragmentation in semen: Systematic review and meta-analysis. Fertil. Steril. 2017, 108, 456–467.e1. [Google Scholar] [CrossRef] [Green Version]
- Esteves, S.C. Testicular versus ejaculated sperm should be used for intracytoplasmic sperm injection (ICSI) in cases of infertility associated with sperm DNA fragmentation|Opinion: Yes. Int. Braz. J. Urol. 2018, 44, 667–675. [Google Scholar] [CrossRef]
- Ambar, R.F.; Agarwal, A.; Majzoub, A.; Vij, S.; Tadros, N.N.; Cho, C.-L.; Parekh, N.; Borges, E.; Glina, S. The Use of Testicular Sperm for Intracytoplasmic Sperm Injection in Patients with High Sperm DNA Damage: A Systematic Review. World J. Men’s Health 2021, 39, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Kort, H.I.; Massey, J.B.; Elsner, C.W.; Mitchell-Leef, D.; Shapiro, D.B.; Witt, M.A.; Roudebush, W.E. Impact of Body Mass Index Values on Sperm Quantity and Quality. J. Androl. 2006, 27, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.; Faure, C.; Sermondade, N.; Boubaya, M.; Eustache, F.; Clément, P.; Briot, P.; Berthaut, I.; Levy, V.; Cedrin-Durnerin, I.; et al. Obesity leads to higher risk of sperm DNA damage in infertile patients. Asian J. Androl. 2013, 15, 622–625. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, S.Y.; Wan, C.W.T.; Law, T.Y.S.; Chan, D.Y.L.; Fok, E.K.L. The Sperm Small RNA Transcriptome: Implications beyond Reproductive Disorder. Int. J. Mol. Sci. 2022, 23, 15716. https://doi.org/10.3390/ijms232415716
Chan SY, Wan CWT, Law TYS, Chan DYL, Fok EKL. The Sperm Small RNA Transcriptome: Implications beyond Reproductive Disorder. International Journal of Molecular Sciences. 2022; 23(24):15716. https://doi.org/10.3390/ijms232415716
Chicago/Turabian StyleChan, Sze Yan, Crystal Wing Tung Wan, Tin Yu Samuel Law, David Yiu Leung Chan, and Ellis Kin Lam Fok. 2022. "The Sperm Small RNA Transcriptome: Implications beyond Reproductive Disorder" International Journal of Molecular Sciences 23, no. 24: 15716. https://doi.org/10.3390/ijms232415716