
A Microwell-Based Intestinal Organoid-Macrophage Co-Culture System to Study Intestinal Inflammation
 , , and
, , and 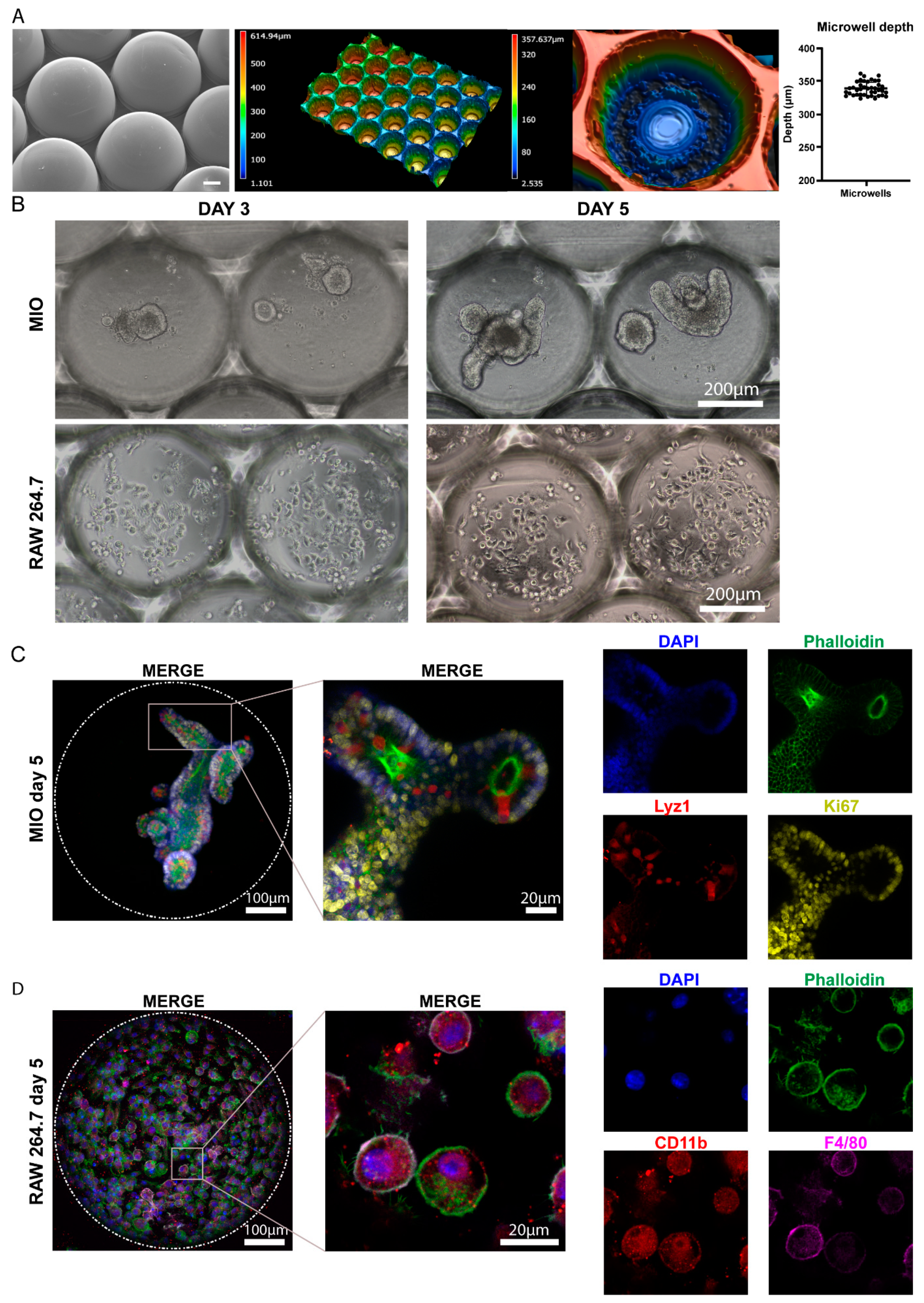{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Monocultures of Intestinal Organoids and RAW 264.7 Macrophages in Microwell Arrays
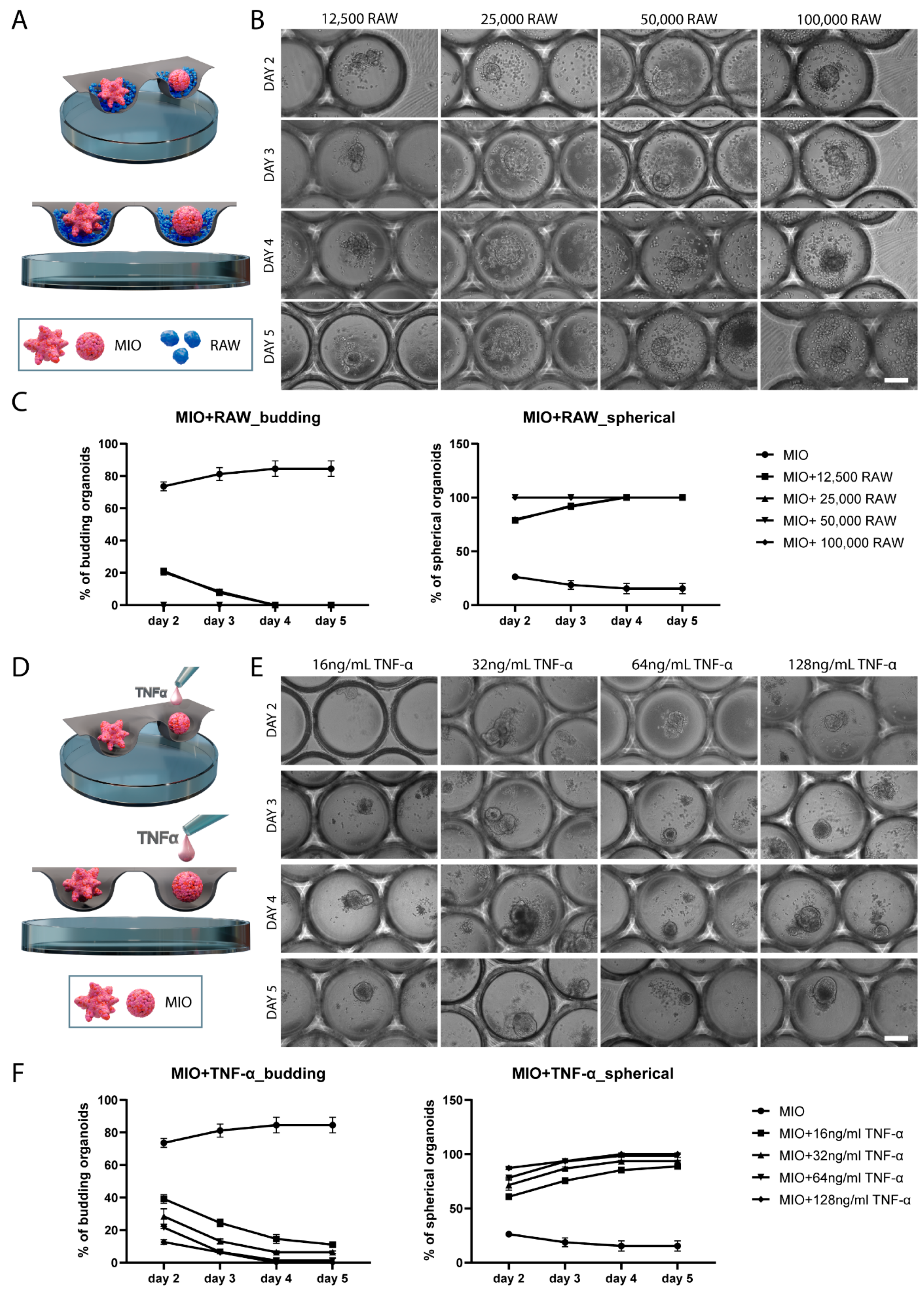
2.2. Direct Co-Culture of Intestinal Organoids and RAW 264.7 Macrophages in Comparison with TNF-α Treatment of Intestinal Organoid Monocultures
2.3. TNF-α-Induces Changes in Organoid Morphology
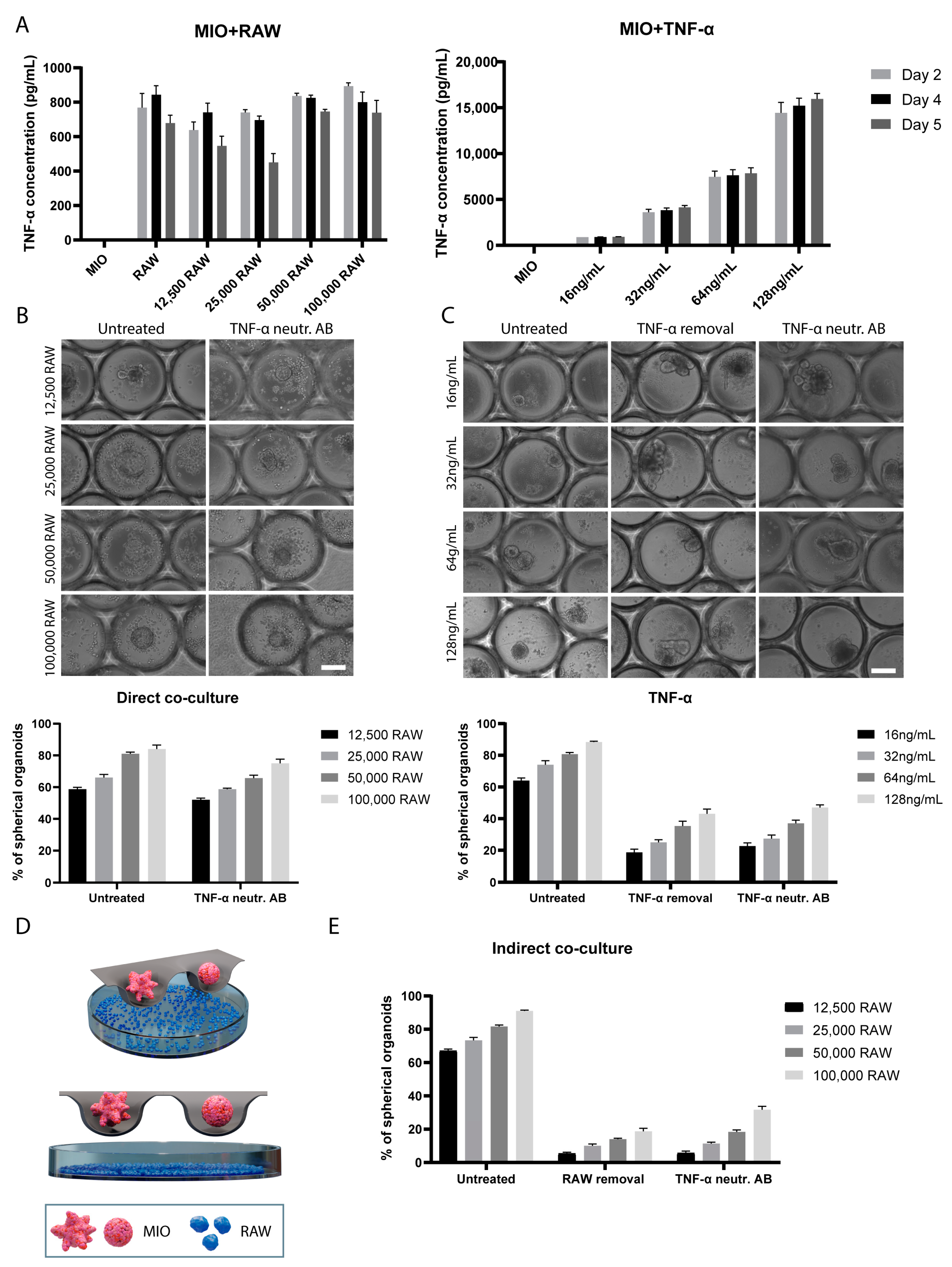
2.4. Indirect Co-Culture of Intestinal Organoids and RAW 264.7 Macrophages
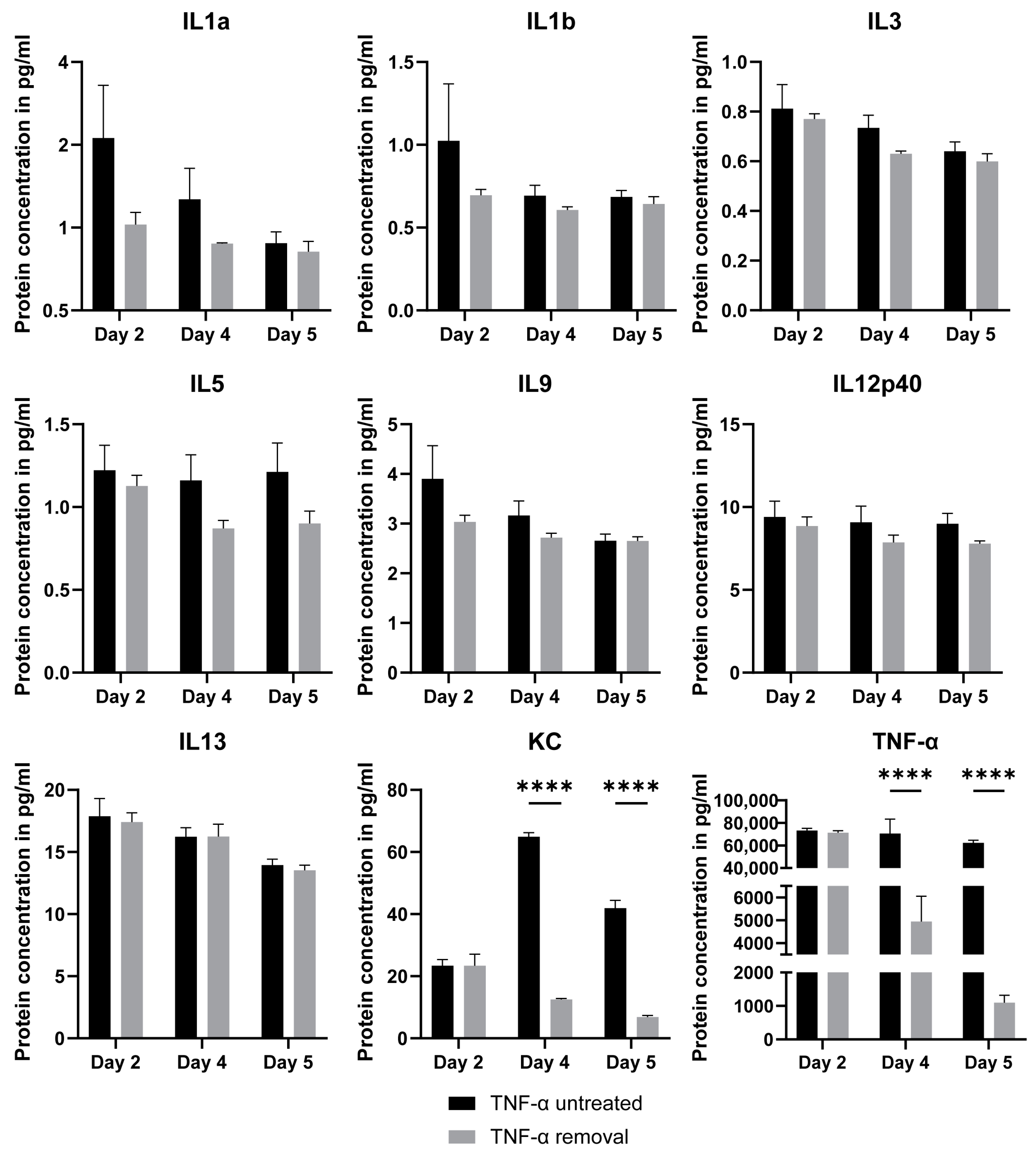
2.5. Luminex Assay for TNF-α-Treated Intestinal Organoids
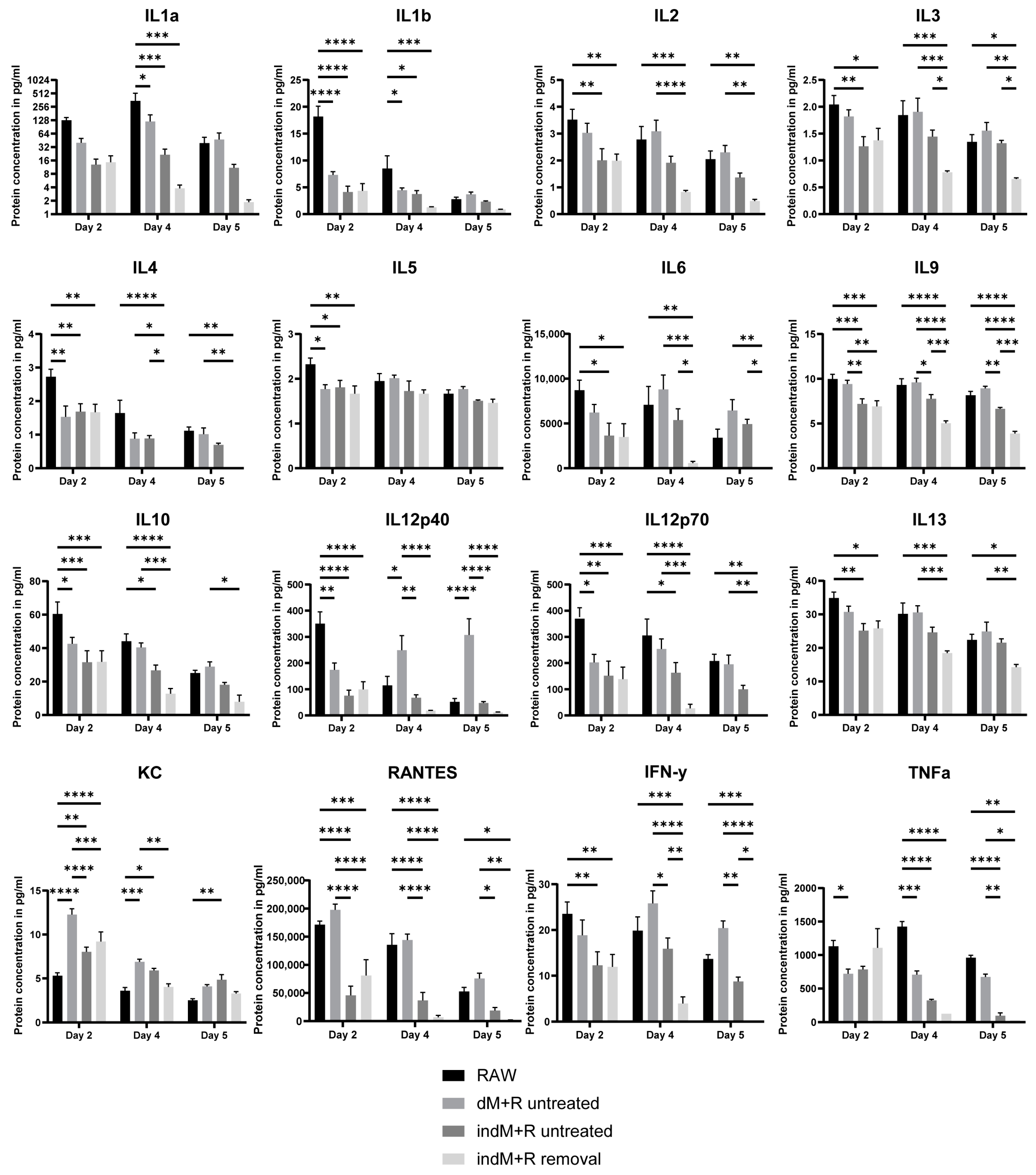
2.6. Luminex Assay for Direct and Indirect Co-Culture of Intestinal Organoids with Macrophages
3. Discussion
4. Materials and Methods
4.1. Fabrication and Preparation of Microwells for Organoid Culture
4.2. Intestinal Organoid Culture
4.3. Macrophage Culture
4.4. Co-Culture of Intestinal Organoids and Macrophages
4.5. Immunofluorescence and Confocal Microscopy
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Luminex Assay
4.8. Scanning Electron Microscopy (SEM)
4.9. Forskolin Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mowat, A.M.; Agace, W.W. Regional Specialization within the Intestinal Immune System. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef]
- Angus, H.C.K.; Butt, A.G.; Schultz, M.; Kemp, R.A. Intestinal Organoids as a Tool for Inflammatory Bowel Disease Research. Front. Med. 2019, 6, 334. [Google Scholar] [CrossRef] [PubMed]
- Wakisaka, Y.; Sugimoto, S.; Sato, T. Organoid Medicine for Inflammatory Bowel Disease. Stem Cells 2022, 40, 123–132. [Google Scholar] [CrossRef]
- Schneeweiss, S.; Korzenik, J.; Solomon, D.H.; Canning, C.; Lee, J.; Bressler, B. Infliximab and Other Immunomodulating Drugs in Patients with Inflammatory Bowel Disease and the Risk of Serious Bacterial Infections. Aliment. Pharmacol. Ther. 2009, 30, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Ten Hove, T.; Van Montfrans, C.; Peppelenbosch, M.P.; Van Deventer, S.J.H. Infliximab Treatment Induces Apoptosis of Lamina Propria T Lymphocytes in Crohn’s Disease. Gut 2002, 50, 206–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, A.; Soni, A.; Acharya, S. In vitro Models and ex vivo Systems Used in Inflammatory Bowel Disease. Vitr. Model. 2022, 1, 213–227. [Google Scholar] [CrossRef]
- Mizoguchi, A. Animal Models of Inflammatory Bowel Disease. Prog. Mol. Biol. Transl. Sci. 2012, 105, 263–320. [Google Scholar] [CrossRef]
- Kiesler, P.; Fuss, I.J.; Strober, W. Experimental Models of Inflammatory Bowel Diseases. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 154–170. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 Stem Cells Build Crypt-Villus Structures in Vitro without a Mesenchymal Niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Spence, J.R.; Mayhew, C.N.; Rankin, S.A.; Kuhar, M.F.; Vallance, J.E.; Tolle, K.; Hoskins, E.E.; Kalinichenko, V.V.; Wells, S.I.; Zorn, A.M.; et al. Directed Differentiation of Human Pluripotent Stem Cells into Intestinal Tissue in Vitro. Nature 2011, 470, 105–109. [Google Scholar] [CrossRef]
- Kakni, P.; Truckenmüller, R.; Habibović, P.; Giselbrecht, S. Challenges to, and Prospects for, Reverse Engineering the Gastrointestinal Tract Using Organoids. Trends Biotechnol. 2022, 40, 932–944. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.; Brevini, T.; Sampaziotis, F.; Neurath, M.F. What Gastroenterologists and Hepatologists Should Know about Organoids in 2019. Dig. Liver Dis. 2019, 51, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.A.; Matheis, F.; Mucida, D. Gut Macrophages: Key Players in Intestinal Immunity and Tissue Physiology. Curr. Opin. Immunol. 2020, 62, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Ding, S.; Jiang, H.; Liu, G. Roles of Macrophages in the Development and Treatment of Gut Inflammation. Front. Cell Dev. Biol. 2021, 9, 385. [Google Scholar] [CrossRef] [PubMed]
- Andrews, C.; McLean, M.H.; Durum, S.K. Cytokine Tuning of Intestinal Epithelial Function. Front. Immunol. 2018, 9, 1270. [Google Scholar] [CrossRef] [Green Version]
- Neurath, M.F. Cytokines in Inflammatory Bowel Disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef]
- Kakni, P.; Hueber, R.; Knoops, K.; López-Iglesias, C.; Truckenmüller, R.; Habibovic, P.; Giselbrecht, S. Intestinal Organoid Culture in Polymer Film-Based Microwell Arrays. Adv. Biosyst. 2020, 40, 932–944. [Google Scholar] [CrossRef]
- Mashukova, A.; Wald, F.A.; Salas, P.J. Tumor Necrosis Factor Alpha and Inflammation Disrupt the Polarity Complex in Intestinal Epithelial Cells by a Posttranslational Mechanism. Mol. Cell. Biol. 2011, 31, 756–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef] [Green Version]
- Bain, C.C.; Schridde, A. Origin, Differentiation, and Function of Intestinal Macrophages. Front. Immunol. 2018, 9, 2733. [Google Scholar] [CrossRef] [Green Version]
- Hahn, S.; Nam, M.O.; Noh, J.H.; Lee, D.H.; Han, H.W.; Kim, D.H.; Hahm, K.B.; Hong, S.P.; Yoo, J.H.; Yoo, J. Organoid-Based Epithelial to Mesenchymal Transition (OEMT) Model: From an Intestinal Fibrosis Perspective. Sci. Rep. 2017, 7, 2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leppkes, M.; Roulis, M.; Neurath, M.F.; Kollias, G.; Becker, C. Pleiotropic Functions of TNF-α in the Regulation of the Intestinal Epithelial Response to Inflammation. Int. Immunol. 2014, 26, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Tanoue, T.; Nishitani, Y.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. In Vitro Model to Estimate Gut Inflammation Using Co-Cultured Caco-2 and RAW264.7 Cells. Biochem. Biophys. Res. Commun. 2008, 374, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.Y.; Boivin, M.A.; Ye, D.; Pedram, A.; Said, H.M. Mechanism of TNF-α Modulation of Caco-2 Intestinal Epithelial Tight Junction Barrier: Role of Myosin Light-Chain Kinase Protein Expression. Am. J. Physiol.—Gastrointest. Liver Physiol. 2005, 288, 422–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Rollins, D.; Qi, Y.; Fredericks, J.; Mansell, T.J.; Jergens, A.; Phillips, G.J.; Wannemuehler, M.; Wang, Q. TNFα Regulates Intestinal Organoids from Mice with Both Defined and Conventional Microbiota. Int. J. Biol. Macromol. 2020, 164, 548–556. [Google Scholar] [CrossRef]
- Lee, C.; Hong, S.N.; Kim, E.R.; Chang, D.K.; Kim, Y.H. Epithelial Regeneration Ability of Crohn’s Disease Assessed Using Patient-Derived Intestinal Organoids. Int. J. Mol. Sci. 2021, 22, 6013. [Google Scholar] [CrossRef]
- Dekkers, J.F.; Wiegerinck, C.L.; De Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; De Winter-De Groot, K.M.; Brandsma, A.M.; De Jong, N.W.M.; Bijvelds, M.J.C.; Scholte, B.J.; et al. A Functional CFTR Assay Using Primary Cystic Fibrosis Intestinal Organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Baniak, N.; Luan, X.; Grunow, A.; Machen, T.E.; Ianowski, J.P. The Cytokines Interleukin-1β and Tumor Necrosis Factor-α Stimulate CFTR-Mediated Fluid Secretion by Swine Airway Submucosal Glands. Am. J. Physiol.—Lung Cell. Mol. Physiol. 2012, 303, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Muzes, G.; Molnár, B.; Tulassay, Z.; Sipos, F. Changes of the Cytokine Profile in Inflammatory Bowel Diseases. World J. Gastroenterol. 2012, 18, 5848. [Google Scholar] [CrossRef] [Green Version]
- Hentschel, V.; Seufferlein, T.; Armacki, M. Intestinal Organoids in Coculture: Redefining the Boundaries of Gut Mucosa Ex Vivo Modeling. Am. J. Physiol.—Gastrointest. Liver Physiol. 2021, 321, G693–G704. [Google Scholar] [CrossRef]
- Noel, G.; Baetz, N.W.; Staab, J.F.; Donowitz, M.; Kovbasnjuk, O.; Pasetti, M.F.; Zachos, N.C. A Primary Human Macrophage-Enteroid Co-Culture Model to Investigate Mucosal Gut Physiology and Host-Pathogen Interactions. Sci. Rep. 2017, 7, 45270. [Google Scholar] [CrossRef] [Green Version]
- Kozuka, K.; He, Y.; Koo-McCoy, S.; Kumaraswamy, P.; Nie, B.; Shaw, K.; Chan, P.; Leadbetter, M.; He, L.; Lewis, J.G.; et al. Development and Characterization of a Human and Mouse Intestinal Epithelial Cell Monolayer Platform. Stem Cell Rep. 2017, 9, 1976–1990. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qi, Z.; Li, X.; Du, Y.; Chen, Y.G. Monolayer Culture of Intestinal Epithelium Sustains Lgr5+ Intestinal Stem Cells. Cell Discov. 2018, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanDussen, K.L.; Marinshaw, J.M.; Shaikh, N.; Miyoshi, H.; Moon, C.; Tarr, P.I.; Ciorba, M.A.; Stappenbeck, T.S. Development of an Enhanced Human Gastrointestinal Epithelial Culture System to Facilitate Patient-Based Assays. Gut 2015, 64, 911–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozaki, K.; Mochizuki, W.; Matsumoto, Y.; Matsumoto, T.; Fukuda, M.; Mizutani, T.; Watanabe, M.; Nakamura, T. Co-Culture with Intestinal Epithelial Organoids Allows Efficient Expansion and Motility Analysis of Intraepithelial Lymphocytes. J. Gastroenterol. 2016, 51, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, S.S.; De Zuani, M.; Tidu, F.; Hortová Kohoutková, M.; Pazzagli, L.; Forte, G.; Spaccapelo, R.; Zelante, T.; Frič, J. Comparison of Two Human Organoid Models of Lung and Intestinal Inflammation Reveals Toll-like Receptor Signalling Activation and Monocyte Recruitment. Clin. Transl. Immunol. 2020, 9, e1131. [Google Scholar] [CrossRef] [PubMed]
- Marchiando, A.M.; Shen, L.; Vallen Graham, W.; Weber, C.R.; Schwarz, B.T.; Austin, J.R.; Raleigh, D.R.; Guan, Y.; Watson, A.J.M.; Montrose, M.H.; et al. Caveolin-1-Dependent Occludin Endocytosis Is Required for TNF-Induced Tight Junction Regulation in Vivo. J. Cell Biol. 2010, 189, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, K.E.; Turner, J.R. Myosin Light Chain Kinase: Pulling the Strings of Epithelial Tight Junction Function. Ann. N. Y. Acad. Sci. 2012, 1258, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Teitelbaum, D.H. Tumour Necrosis Factor-α-Induced Loss of Intestinal Barrier Function Requires TNFR1 and TNFR2 Signalling in a Mouse Model of Total Parenteral Nutrition. J. Physiol. 2013, 591, 3709. [Google Scholar] [CrossRef] [Green Version]
- Feghali, C.A.; Wright, T.M. Cytokines in Acute and Chronic Inflammation. Front. Biosci. 1997, 2, 12–26. [Google Scholar] [CrossRef]
- Chomarat, P.; Banchereau, J. Interleukin-4 and Lnterleukin-13: Their Similarities and Discrepancies. Int. Rev. Immunol. 2009, 17, 1–52. [Google Scholar] [CrossRef]
- West, G.A.; Matsuura, T.; Levine, A.D.; Klein, J.S.; Fiocchi, C. Interleukin 4 in Inflammatory Bowel Disease and Mucosal Immune Reactivity. Gastroenterology 1996, 110, 1683–1695. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dranoff, G.; Dougan, S.K. GM-CSF, IL-3, and IL-5 Family of Cytokines: Regulators of Inflammation. Immunity 2019, 50, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Borriello, F.; Galdiero, M.R.; Varricchi, G.; Loffredo, S.; Spadaro, G.; Marone, G. Innate Immune Modulation by GM-CSF and IL-3 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalleweg, N.; Chiriac, M.T.; Podstawa, E.; Lehmann, C.; Rau, T.T.; Atreya, R.; Krauss, E.; Hundorfean, G.; Fichtner-Feigl, S.; Hartmann, A.; et al. IL-9 and Its Receptor Are Predominantly Involved in the Pathogenesis of UC. Gut 2014, 64, 743–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajuebor, M.N.; Swain, M.G. Role of Chemokines and Chemokine Receptors in the Gastrointestinal Tract. Immunology 2002, 105, 137. [Google Scholar] [CrossRef] [PubMed]
- Ansari, N.; Abdulla, J.; Zayyani, N.; Brahmi, U.; Taha, S.; Satir, A.A. Comparison of RANTES Expression in Crohn’s Disease and Ulcerative Colitis: An Aid in the Differential Diagnosis? J. Clin. Pathol. 2006, 59, 1066. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Xu, X.; Luo, X.; Li, L.; Huang, B.; Li, X.; Tao, D.; Hu, J.; Gong, J. Role of RANTES and Its Receptor in Gastric Cancer Metastasis. J. Huazhong Univ. Sci. Technol. Med. Sci. 2011, 31, 342–347. [Google Scholar] [CrossRef]
- Puleston, J.; Cooper, M.; Murch, S.; Bid, K.; Makh, S.; Ashwood, P.; Bingham, A.H.; Green, H.; Moss, P.; Dhillon, A.; et al. A Distinct Subset of Chemokines Dominates the Mucosal Chemokine Response in Inflammatory Bowel Disease. Aliment. Pharmacol. Ther. 2005, 21, 109–120. [Google Scholar] [CrossRef]
- Singh, U.P.; Singh, N.P.; Murphy, E.A.; Price, R.L.; Fayad, R.; Nagarkatti, M.; Nagarkatti, P.S. Chemokine and Cytokine Levels in Inflammatory Bowel Disease Patients. Cytokine 2016, 77, 44. [Google Scholar] [CrossRef] [PubMed]
- Giselbrecht, S.; Gietzelt, T.; Gottwald, E.; Trautmann, C.; Truckenmüller, R.; Weibezahn, K.F.; Welle, A. 3D Tissue Culture Substrates Produced by Microthermoforming of Pre-Processed Polymer Films. Biomed. Microdevices 2006, 8, 191–199. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakni, P.; Truckenmüller, R.; Habibović, P.; van Griensven, M.; Giselbrecht, S. A Microwell-Based Intestinal Organoid-Macrophage Co-Culture System to Study Intestinal Inflammation. Int. J. Mol. Sci. 2022, 23, 15364. https://doi.org/10.3390/ijms232315364
Kakni P, Truckenmüller R, Habibović P, van Griensven M, Giselbrecht S. A Microwell-Based Intestinal Organoid-Macrophage Co-Culture System to Study Intestinal Inflammation. International Journal of Molecular Sciences. 2022; 23(23):15364. https://doi.org/10.3390/ijms232315364
Chicago/Turabian StyleKakni, Panagiota, Roman Truckenmüller, Pamela Habibović, Martijn van Griensven, and Stefan Giselbrecht. 2022. "A Microwell-Based Intestinal Organoid-Macrophage Co-Culture System to Study Intestinal Inflammation" International Journal of Molecular Sciences 23, no. 23: 15364. https://doi.org/10.3390/ijms232315364