

Unveiling the Metal-Dependent Aggregation Properties of the C-terminal Region of Amyloidogenic Intrinsically Disordered Protein Isoforms DPF3b and DPF3a


Abstract
:1. Introduction
2. Results
2.1. DPF3 C-terminal Regions Have Intrinsic Disorder Properties
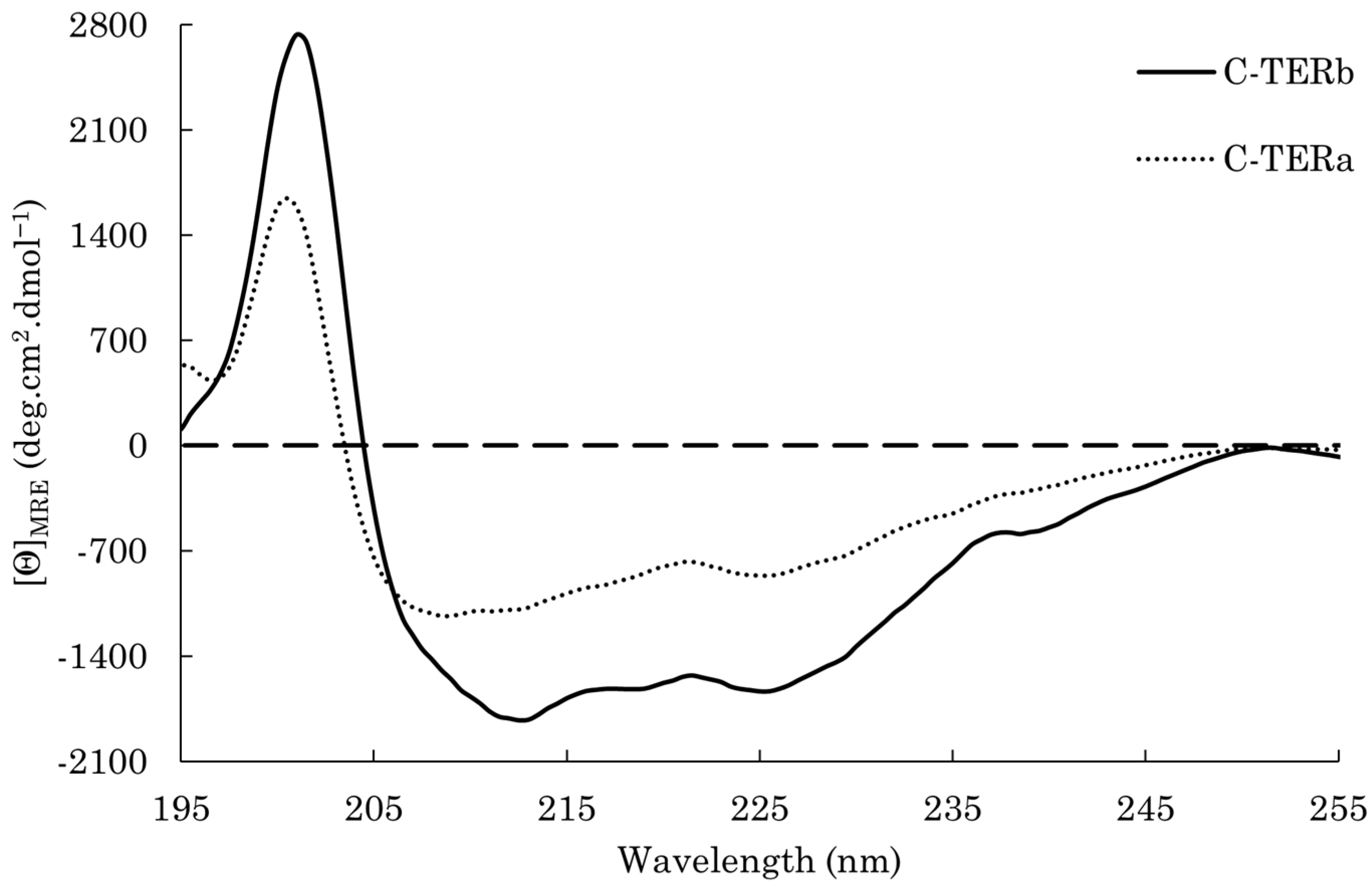
2.2. DPF3 C-terminal Regions Are Prone to Aggregate In Vitro
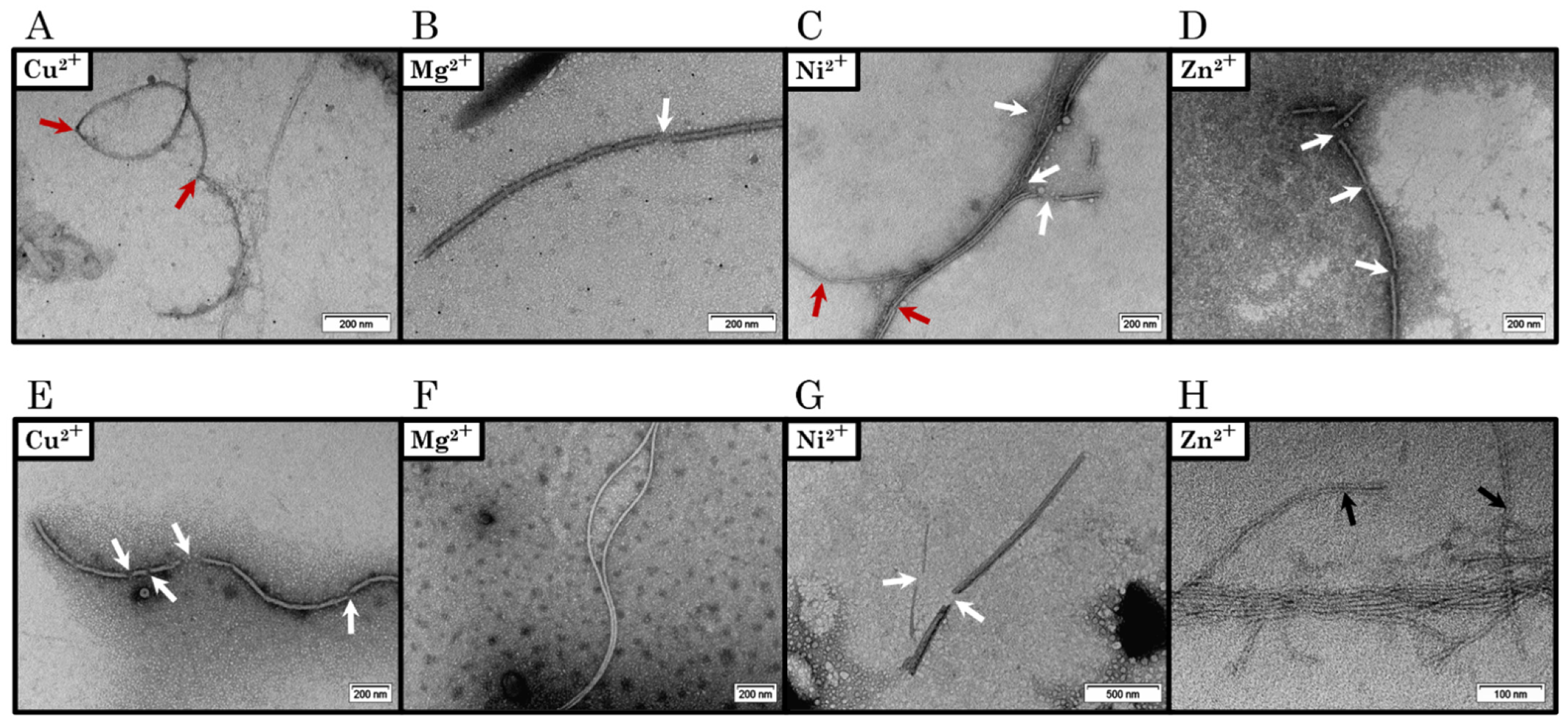
2.3. Transmission Electron Microscopy Reveals the Morphologic Diversity of DPF3 C-terminal Region Aggregates
2.4. Metal Cations Have Distinct Effects on the Aggregation of DPF3 C-terminal Regions
3. Discussion
4. Materials and Methods
4.1. Overexpression and Purification of C-terminal Regions
4.2. Protein Concentration and UV–Visible Absorption Spectroscopy
4.3. Metal Cation and Protein Sample Preparation
4.4. Bioinformatics Analyses
4.5. Far- and Near-UV Circular Dichroism Spectroscopy
4.6. Intrinsic Fluorescence Spectroscopy (ITF, ITyrF, and dbAF)
4.7. Thioflavin T Binding Assay
4.8. Transmission Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lange, M.; Kaynak, B.; Forster, U.B.; Tönjes, M.; Fischer, J.J.; Grimm, C.; Schlesinger, J.; Just, S.; Dunkel, I.; Krueger, T.; et al. Regulation of Muscle Development by DPF3, a Novel Histone Acetylation and Methylation Reader of the BAF Chromatin Remodeling Complex. Genes Dev. 2008, 22, 2370–2384. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Zhang, Q.; Li, S.; Plotnikov, A.N.; Walsh, M.J.; Zhou, M.M. Mechanism and Regulation of Acetylated Histone Binding by the Tandem PHD Finger of DPF3b. Nature 2010, 466, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhao, A.; Tempel, W.; Loppnau, P.; Liu, Y. Crystal Structure of DPF3b in Complex with an Acetylated Histone Peptide. J. Struct. Biol. 2016, 195, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Schlesinger, J.; Schoenhals, S.; Tönjes, M.; Dunkel, I.; Meierhofer, D.; Cano, E.; Schulz, K.; Berger, M.F.; Haack, T.; et al. Phosphorylation of the Chromatin Remodeling Factor DPF3a Induces Cardiac Hypertrophy through Releasing HEY Repressors from DNA. Nucleic Acids Res. 2015, 44, 2538–2553. [Google Scholar] [CrossRef] [Green Version]
- Guanglei, W.; Bingbing, W.; Peixin, Y. Epigenetics in Congenital Heart Disease. J. Am. Heart Assoc. 2022, 11, e025163. [Google Scholar] [CrossRef]
- Liu, S.Y.; Zhang, C.J.; Peng, H.Y.; Sun, H.; Lin, K.Q.; Huang, X.Q.; Huang, K.; Chu, J.Y.; Yang, Z.Q. Strong Association of SLC1A1 and DPF3 Gene Variants with Idiopathic Male Infertility in Han Chinese. Asian J. Androl. 2016, 18, 486–492. [Google Scholar] [CrossRef]
- Sato, Y.; Hasegawa, C.; Tajima, A.; Nozawa, S.; Yoshiike, M.; Koh, E.; Kanaya, J.; Namiki, M.; Matsumiya, K.; Tsujimura, A.; et al. Association of TUSC1 and DPF3 Gene Polymorphisms with Male Infertility. J. Assist. Reprod. Genet. 2018, 35, 257–263. [Google Scholar] [CrossRef]
- Liu, H.; Luo, Y.; Li, S.; Wang, S.; Wang, N.; Jin, X. Expression Profiles of HA117 and Its Neighboring Gene DPF3 in Different Colon Segments of Hirschsprung’s Disease. Int. J. Clin. Exp. Pathol. 2014, 7, 3966–3974. [Google Scholar]
- Lin, W.-H.; Dai, W.-G.; Xu, X.-D.; Yu, Q.-H.; Zhang, B.; Li, J.; Li, H.-P. Downregulation of DPF3 Promotes the Proliferation and Motility of Breast Cancer Cells through Activating JAK2/STAT3 Signaling. Biochem. Biophys. Res. Commun. 2019, 514, 639–644. [Google Scholar] [CrossRef]
- Theodorou, M.; Speletas, M.; Mamara, A.; Papachristopoulou, G.; Lazou, V.; Scorilas, A.; Katsantoni, E. Identification of a STAT5 Target Gene, Dpf3, Provides Novel Insights in Chronic Lymphocytic Leukemia. PLoS ONE 2013, 8, e76155. [Google Scholar] [CrossRef]
- Hiramatsu, H.; Kobayashi, K.; Kobayashi, K.; Haraguchi, T.; Ino, Y.; Todo, T.; Iba, H. The Role of the SWI/SNF Chromatin Remodeling Complex in Maintaining the Stemness of Glioma Initiating Cells. Sci. Rep. 2017, 7, 889. [Google Scholar] [CrossRef]
- Banaganapalli, B.; Mallah, B.; Alghamdi, K.S.; Albaqami, W.F.; Alshaer, D.S.; Alrayes, N.; Elango, R.; Shaik, N.A. Integrative Weighted Molecular Network Construction from Transcriptomics and Genome Wide Association Data to Identify Shared Genetic Biomarkers for COPD and Lung Cancer. PLoS ONE 2022, 17, e0274629. [Google Scholar] [CrossRef]
- Colli, L.M.; Jessop, L.; Myers, T.A.; Camp, S.Y.; Machiela, M.J.; Choi, J.; Cunha, R.; Onabajo, O.; Mills, G.C.; Schmid, V.; et al. Altered Regulation of DPF3, a Member of the SWI/SNF Complexes, Underlies the 14q24 Renal Cancer Susceptibility Locus. Am. J. Hum. Genet. 2021, 108, 1590–1610. [Google Scholar] [CrossRef]
- Protze, J.; Naas, S.; Krüger, R.; Stöhr, C.; Kraus, A.; Grampp, S.; Wiesener, M.; Schiffer, M.; Hartmann, A.; Wullich, B.; et al. The Renal Cancer Risk Allele at 14q24.2 Activates a Novel Hypoxia-Inducible Transcription Factor-Binding Enhancer of DPF3 Expression. J. Biol. Chem. 2022, 3, 298. [Google Scholar] [CrossRef]
- Cui, H.; Yi, H.; Bao, H.; Tan, Y.; Tian, C.; Shi, X.; Gan, D.; Zhang, B.; Liang, W.; Chen, R.; et al. The SWI/SNF Chromatin Remodeling Factor DPF3 Regulates Metastasis of CcRCC by Modulating TGF-β Signaling. Nat. Commun. 2022, 13, 4680. [Google Scholar] [CrossRef]
- Mignon, J.; Mottet, D.; Verrillo, G.; Matagne, A.; Perpète, E.A.; Michaux, C. Revealing Intrinsic Disorder and Aggregation Properties of the DPF3a Zinc Finger Protein. ACS Omega 2021, 6, 18793–18801. [Google Scholar] [CrossRef]
- Mignon, J.; Mottet, D.; Leyder, T.; Uversky, V.N.; Perp, E.A.; Michaux, C. Structural Characterisation of Amyloidogenic Intrinsically Disordered Zinc Finger Protein Isoforms DPF3b and DPF3a. Int. J. Biol. Macromol. 2022, 218, 57–71. [Google Scholar] [CrossRef]
- Ayyadevara, S.; Ganne, A.; Balasubramaniam, M.; Reis, R.J.S. Intrinsically Disordered Proteins Identified in the Aggregate Proteome Serve as Biomarkers of Neurodegeneration. Metab. Brain Dis. 2022, 37, 147–152. [Google Scholar] [CrossRef]
- Mezzaroba, L.; Alfieri, D.F.; Simão, A.N.C.; Reiche, E.M.V. The Role of Zinc, Copper, Manganese and Iron in Neurodegenerative Diseases. Neurotoxicology 2019, 74, 230–241. [Google Scholar] [CrossRef]
- Kirkland, A.E.; Sarlo, G.L.; Holton, K.F. The Role of Magnesium in Neurological Disorders. Nutrients 2018, 10, 730. [Google Scholar] [CrossRef] [Green Version]
- Anyachor, C.P.; Dooka, D.B.; Orish, C.N.; Amadi, C.N.; Bocca, B.; Ruggieri, F.; Senofonte, M.; Frazzoli, C.; Orisakwe, O.E. Mechanistic Considerations and Biomarkers Level in Nickel-Induced Neurodegenerative Diseases: An Updated Systematic Review. IBRO Neurosci. Rep. 2022, 13, 136–146. [Google Scholar] [CrossRef]
- Breydo, L.; Uversky, V.N. Role of Metal Ions in Aggregation of Intrinsically Disordered Proteins in Neurodegenerative Diseases. Metallomics 2011, 3, 1163–1180. [Google Scholar] [CrossRef]
- Leal, S.S.; Botelho, H.M.; Gomes, C.M. Metal Ions as Modulators of Protein Conformation and Misfolding in Neurodegeneration. Coord. Chem. Rev. 2012, 256, 2253–2270. [Google Scholar] [CrossRef]
- Faller, P.; Hureau, C.; La Penna, G. Metal Ions and Intrinsically Disordered Proteins and Peptides: From Cu/Zn Amyloid-β to General Principles. Acc. Chem. Res. 2014, 47, 2252–2259. [Google Scholar]
- Poulson, B.G.; Szczepski, K.; Lachowicz, J.I.; Jaremko, L.; Emwas, A.H.; Jaremko, M. Aggregation of Biologically Important Peptides and Proteins: Inhibition or Acceleration Depending on Protein and Metal Ion Concentrations. RSC Adv. 2019, 10, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Arena, G.; Rizzarelli, E. Zn2+ Interaction with Amyloid-B: Affinity and Speciation. Molecules 2019, 24, 2796. [Google Scholar] [CrossRef] [Green Version]
- Rana, M.; Sharma, A.K. Cu and Zn Interactions with Aβ Peptides: Consequence of Coordination on Aggregation and Formation of Neurotoxic Soluble Aβ Oligomers. Metallomics 2019, 11, 64–84. [Google Scholar] [CrossRef]
- Cristóvão, J.S.; Santos, R.; Gomes, C.M. Metals and Neuronal Metal Binding Proteins Implicated in Alzheimer’s Disease. Oxid. Med. Cell Longev. 2016, 2016, 9812178. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G. A Simple Method for Detecting Phosphorylation of Proteins by Using Zn2+ -Phos-Tag SDS-PAGE at Neutral PH. Methods Mol. Biol. 2018, 1853, 223–229. [Google Scholar] [CrossRef]
- Ahmadi, S.; Zhu, S.; Sharma, R.; Wu, B.; Soong, R.; Dutta Majumdar, R.; Wilson, D.J.; Simpson, A.J.; Kraatz, H.B. Aggregation of Microtubule Binding Repeats of Tau Protein Is Promoted by Cu 2+. ACS Omega 2019, 4, 5356–5366. [Google Scholar] [CrossRef] [Green Version]
- Gorantla, N.V.; Das, R.; Balaraman, E.; Chinnathambi, S. Transition Metal Nickel Prevents Tau Aggregation in Alzheimer’s Disease. Int. J. Biol. Macromol. 2020, 156, 1359–1365. [Google Scholar] [CrossRef]
- Uversky, V.N.; Li, J.; Fink, A.L. Metal-Triggered Structural Transformations, Aggregation, and Fibrillation of Human α-Synuclein: A Possible Molecular Link between Parkinson’s Disease and Heavy Metal Exposure. J. Biol. Chem. 2001, 276, 44284–44296. [Google Scholar] [CrossRef]
- Wright, J.A.; Wang, X.; Brown, D.R. Unique Copper-induced Oligomers Mediate Alpha-synuclein Toxicity. FASEB J. 2009, 23, 2384–2393. [Google Scholar] [CrossRef]
- Golts, N.; Snyder, H.; Frasier, M.; Theisler, C.; Choi, P.; Wolozin, B. Magnesium Inhibits Spontaneous and Iron-Induced Aggregation of α-Synuclein. J. Biol. Chem. 2002, 277, 16116–16123. [Google Scholar] [CrossRef] [Green Version]
- Valiente-Gabioud, A.A.; Torres-Monserrat, V.; Molina-Rubino, L.; Binolfi, A.; Griesinger, C.; Fernández, C.O. Structural Basis behind the Interaction of Zn2 + with the Protein α-Synuclein and the Aβ Peptide: A Comparative Analysis. J. Inorg. Biochem. 2012, 117, 334–341. [Google Scholar]
- Al-Harthi, S.; Kharchenko, V.; Mandal, P.; Gourdoupis, S.; Jaremko, L. Zinc Ions Prevent α-Synuclein Aggregation by Enhancing Chaperone Function of Human Serum Albumin. Int. J. Biol. Macromol. 2022, 222, 2878–2887. [Google Scholar] [CrossRef]
- Blalock, E.M.; Geddes, J.W.; Chen, K.C.; Porter, N.M.; Markesbery, W.R.; Landfield, P.W. Incipient Alzheimer’s Disease: Microarray Correlation Analyses Reveal Major Transcriptional and Tumor Suppressor Responses. Proc. Natl. Acad. Sci. USA 2004, 101, 2173–2178. [Google Scholar] [CrossRef] [Green Version]
- Satoh, J.I. Molecular Network Analysis of Human MicroRNA Targetome: From Cancers to Alzheimers Disease. BioData Min. 2012, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Rudnitskaya, E.A.; Burnyasheva, A.O.; Kozlova, T.A.; Peunov, D.A.; Kolosova, N.G.; Stefanova, N.A. Changes in Glial Support of the Hippocampus during the Development of an Alzheimer’s Disease-like Pathology and Their Correction by Mitochondria-Targeted Antioxidant SkQ1. Int. J. Mol. Sci. 2022, 23, 1134. [Google Scholar] [CrossRef]
- Huang, M.; Xu, L.; Liu, J.; Huang, P.; Tan, Y.; Chen, S. Cell–Cell Communication Alterations via Intercellular Signaling Pathways in Substantia Nigra of Parkinson’s Disease. Front. Aging Neurosci. 2022, 14, 828457. [Google Scholar] [CrossRef]
- Uversky, V.N. Intrinsically Disordered Proteins and Their “Mysterious” (Meta) Physics. Front. Phys. 2019, 7, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Oldfield, C.J.; Dunker, A.K.; Uversky, V.N. CDF It All: Consensus Prediction of Intrinsically Disordered Proteins Based on Various Cumulative Distribution Functions. FEBS Lett. 2009, 583, 1469–1474. [Google Scholar] [CrossRef] [Green Version]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why Are “natively Unfolded” Proteins Unstructured under Physiologic Conditions? Proteins Struct. Funct. Genet. 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Das, R.K.; Pappu, R.V. Conformations of Intrinsically Disordered Proteins Are Influenced by Linear Sequence Distributions of Oppositely Charged Residues. Proc. Natl. Acad. Sci. USA 2013, 110, 13392–13397. [Google Scholar] [CrossRef] [Green Version]
- Holehouse, A.S.; Das, R.K.; Ahad, J.N.; Richardson, M.O.G.; Pappu, R.V. CIDER: Resources to Analyze Sequence-Ensemble Relationships of Intrinsically Disordered Proteins. Biophys. J. 2017, 112, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Das, R.K.; Ruff, K.M.; Pappu, R.V. Relating Sequence Encoded Information to Form and Function of Intrinsically Disordered Proteins. Curr. Opin. Struct. Biol. 2015, 32, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.W.; Holehouse, A.S.; Grace, C.R.; Hughes, A.; Pappu, R.V.; Mittag, T. Sequence Determinants of the Conformational Properties of an Intrinsically Disordered Protein Prior to and upon Multisite Phosphorylation. J. Am. Chem. Soc. 2016, 138, 15323–15335. [Google Scholar] [CrossRef] [Green Version]
- Biter, A.B.; Pollet, J.; Chen, W.H.; Strych, U.; Hotez, P.J.; Bottazzi, M.E. A Method to Probe Protein Structure from UV Absorbance Spectra. Anal. Biochem. 2019, 587, 113450. [Google Scholar] [CrossRef]
- Schramm, A.; Bignon, C.; Brocca, S.; Grandori, R.; Santambrogio, C.; Longhi, S. An Arsenal of Methods for the Experimental Characterization of Intrinsically Disordered Proteins—How to Choose and Combine Them? Arch. Biochem. Biophys. 2019, 676, 108055. [Google Scholar] [CrossRef]
- Gong, Z.; Ikonomova, S.P.; Karlsson, A.J. Secondary Structure of Cell-Penetrating Peptides during Interaction with Fungal Cells. Protein Sci. 2018, 27, 702–713. [Google Scholar] [CrossRef] [Green Version]
- Tromelin, A.; Andriot, I.; Guichard, E. Protein-Flavour Interactions; Woodhead Publishing Limited: Sawston, UK, 2006; ISBN 9781855739604. [Google Scholar]
- Vivian, J.T.; Callis, P.R. Mechanisms of Tryptophan Fluorescence Shifts in Proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [Green Version]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer Science: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Davis, K.B.; Zhang, Z.; Karpova, E.A.; Zhang, J. Application of Tyrosine-Tryptophan Fluorescence Resonance Energy Transfer in Monitoring Protein Size Changes. Anal. Biochem. 2018, 557, 142–150. [Google Scholar] [CrossRef]
- Zhdanova, N.G.; Maksimov, E.G.; Arutyunyan, A.M.; Fadeev, V.V.; Shirshin, E.A. Tyrosine Fluorescence Probing of Conformational Changes in Tryptophan-Lacking Domain of Albumins. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 174, 223–229. [Google Scholar] [CrossRef]
- Khrapunov, S.; Pastor, N.; Brenowitz, M. Solution Structural Studies of the Saccharomyces Cerevisiae TATA Binding Protein (TBP). Biochemistry 2002, 41, 9559–9571. [Google Scholar] [CrossRef]
- Szabo, A.G.; Lynn, K.R.; Krajcarski, D.T.; Rayner, D.M. Tyrosinate Fluorescence Maxima at 345 Nm in Proteins Lacking Tryptophan at PH 7. FEBS Lett. 1978, 94, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Avila, J.; Jiménez, J.S.; Sayas, C.L.; Bolós, M.; Zabala, J.C.; Rivas, G.; Hernández, F. Tau Structures. Front. Aging Neurosci. 2016, 8, 262. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.F.; Xu, T.H.; Yan, Y.; Zhou, Y.R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid Beta: Structure, Biology and Structure-Based Therapeutic Development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [Green Version]
- Camino, J.D.; Gracia, P.; Chen, S.W.; Sot, J.; De La Arada, I.; Sebastián, V.; Arrondo, J.L.R.; Goñi, F.M.; Dobson, C.M.; Cremades, N. The Extent of Protein Hydration Dictates the Preference for Heterogeneous or Homogeneous Nucleation Generating Either Parallel or Antiparallel β-Sheet α-Synuclein Aggregates. Chem. Sci. 2020, 11, 11902–11914. [Google Scholar] [CrossRef]
- Gracia, P.; Camino, J.D.; Volpicelli-daley, L.; Cremades, N. Multiplicity of α-Synuclein Aggregated Species and Their Possible Roles in Disease. Int. J. Mol. Sci. 2020, 21, 8043. [Google Scholar] [CrossRef]
- Pignataro, M.F.; Herrera, M.G.; Dodero, V.I. Evaluation of Peptide/Protein Self-Assembly and Aggregation by Spectroscopic Methods. Molecules 2020, 25, 4854. [Google Scholar]
- Naeem, A.; Khan, T.A.; Muzaffar, M.; Ahmad, S.; Saleemuddin, M. A Partially Folded State of Ovalbumin at Low PH Tends to Aggregate. Cell Biochem. Biophys. 2011, 59, 29–38. [Google Scholar] [CrossRef]
- Szasz, C.; Alexa, A.; Toth, K.; Rakacs, M.; Langowski, J.; Tompa, P. Protein Disorder Prevails under Crowded Conditions. Biochemistry 2011, 50, 5834–5844. [Google Scholar] [CrossRef]
- Abe, H.; Nakanishi, H. Novel Observation of a Circular Dichroism Band Originating from Amyloid Fibril. Anal. Sci. 2003, 19, 171–173. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, E.; Kędracka-Krok, S.; Bystranowska, D.; Ptak, M.; Wiak, K.; Wygralak, Z.; Jankowska, U.; Ożyhar, A. Destabilisation of the Structure of Transthyretin Is Driven by Ca2+. Int. J. Biol. Macromol. 2021, 166, 409–423. [Google Scholar] [CrossRef]
- Wygralak, Z.; Sylwia, K.; Bezara, P. Deep Blue Autofluorescence Reflects the Oxidation State of Human Transthyretin. Redox Biol. 2022, 56, 102434. [Google Scholar] [CrossRef]
- Pinotsi, D.; Grisanti, L.; Mahou, P.; Gebauer, R.; Kaminski, C.F.; Hassanali, A.; Kaminski Schierle, G.S. Proton Transfer and Structure-Specific Fluorescence in Hydrogen Bond-Rich Protein Structures. J. Am. Chem. Soc. 2016, 138, 3046–3057. [Google Scholar] [CrossRef] [Green Version]
- Stephens, A.D.; Qaisrani, M.N.; Ruggiero, M.T.; Mirón, G.D.; Morzan, U.N.; González Lebrero, M.C.; Jones, S.T.E.; Poli, E.; Bond, A.D.; Woodhams, P.J.; et al. Short Hydrogen Bonds Enhance Nonaromatic Protein-Related Fluorescence. Proc. Natl. Acad. Sci. USA 2021, 118, e2020389118. [Google Scholar] [CrossRef]
- Grelich-Mucha, M.; Garcia, A.M.; Torbeev, V.; Ożga, K.; Berlicki, Ł.; Olesiak-Bańska, J. Autofluorescence of Amyloids Determined by Enantiomeric Composition of Peptides. J. Phys. Chem. B 2021, 125, 5502–5510. [Google Scholar] [CrossRef]
- Shaham-Niv, S.; Arnon, Z.A.; Sade, D.; Lichtenstein, A.; Shirshin, E.A.; Kolusheva, S.; Gazit, E. Intrinsic Fluorescence of Metabolite Amyloids Allows Label-Free Monitoring of Their Formation and Dynamics in Live Cells. Angew. Chemie—Int. Ed. 2018, 57, 12444–12447. [Google Scholar] [CrossRef]
- Ren, X.; Zou, Q.; Yuan, C.; Chang, R.; Xing, R.; Yan, X. The Dominant Role of Oxygen in Modulating the Chemical Evolution Pathways of Tyrosine in Peptides: Dityrosine or Melanin. Angew. Chemie—Int. Ed. 2019, 58, 5872–5876. [Google Scholar] [CrossRef]
- Arnon, Z.A.; Kreiser, T.; Yakimov, B.; Brown, N.; Aizen, R.; Shaham-Niv, S.; Makam, P.; Qaisrani, M.N.; Poli, E.; Ruggiero, A.; et al. On-off Transition and Ultrafast Decay of Amino Acid Luminescence Driven by Modulation of Supramolecular Packing. iScience 2021, 24, 102695. [Google Scholar] [CrossRef]
- Fricano, A.; Librizzi, F.; Rao, E.; Alfano, C.; Vetri, V. Blue Autofluorescence in Protein Aggregates “Lighted on” by UV Induced Oxidation. Biochim. Biophys. Acta—Proteins Proteom. 2019, 1867, 140258. [Google Scholar] [CrossRef]
- Niyangoda, C.; Miti, T.; Breydo, L.; Uversky, V.; Muschol, M. Carbonyl-Based Blue Autofluorescence of Proteins and Amino Acids. PLoS ONE 2017, 12, e0176983. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.A. Interpretation of α-Synuclein UV Absorption Spectra in the Peptide Bond and the Aromatic Regions. J. Photochem. Photobiol. B Biol. 2020, 212, 112022. [Google Scholar] [CrossRef]
- Robustelli, P.; Ibanez-De-Opakua, A.; Campbell-Bezat, C.; Giordanetto, F.; Becker, S.; Zweckstetter, M.; Pan, A.C.; Shaw, D.E. Molecular Basis of Small-Molecule Binding to α-Synuclein. J. Am. Chem. Soc. 2022, 144, 2501–2510. [Google Scholar] [CrossRef]
- Alali, S.; Riazi, G.; Ashrafi-Kooshk, M.R.; Meknatkhah, S.; Ahmadian, S.; Ardakani, M.H.; Hosseinkhani, B. Cannabidiol Inhibits Tau Aggregation in Vitro. Cells 2021, 10, 3521. [Google Scholar] [CrossRef]
- Xue, C.; Lin, T.Y.; Chang, D.; Guo, Z. Thioflavin T as an Amyloid Dye: Fibril Quantification, Optimal Concentration and Effect on Aggregation. R. Soc. Open Sci. 2017, 4, 160696. [Google Scholar] [CrossRef] [Green Version]
- Close, W.; Neumann, M.; Schmidt, A.; Hora, M.; Annamalai, K.; Schmidt, M.; Reif, B.; Schmidt, V.; Grigorieff, N.; Fändrich, M. Physical Basis of Amyloid Fibril Polymorphism. Nat. Commun. 2018, 9, 699. [Google Scholar] [CrossRef] [Green Version]
- Tikhonova, T.N.; Rovnyagina, N.R.; Zherebker, A.Y.; Sluchanko, N.N.; Rubekina, A.A.; Orekhov, A.S.; Nikolaev, E.N.; Fadeev, V.V.; Uversky, V.N.; Shirshin, E.A. Dissection of the Deep-Blue Autofluorescence Changes Accompanying Amyloid Fibrillation. Arch. Biochem. Biophys. 2018, 651, 13–20. [Google Scholar] [CrossRef]
- Meisl, G.; Kirkegaard, J.B.; Arosio, P.; Michaels, T.C.T.; Vendruscolo, M.; Dobson, C.M.; Linse, S.; Knowles, T.P.J. Molecular Mechanisms of Protein Aggregation from Global Fitting of Kinetic Models. Nat. Protoc. 2016, 11, 252–272. [Google Scholar] [CrossRef]
- Mittal, M.; Gautam, S.; Chowdhury, P.K.; Deep, S.; Sapra, S. Role of Tryptophan in Protein-Nanocrystals Interaction: Energy or Charge Transfer. Zeitschrift fur Phys. Chemie 2019, 233, 41–54. [Google Scholar] [CrossRef]
- Prendergast, F.G.; Hampton, P.D.; Jones, B. Characteristics of Tyrosinate Fluorescence Emission in A- and β-Purothionins. Biochemistry 1984, 23, 6690–6697. [Google Scholar] [CrossRef]
- Nag, N.; Bahadur, P.; Uversky, V.N.; Giri, R.; Tripathi, T. Experimental Methods to Study Intrinsically Disordered Proteins. In Advances in Protein Molecular and Structural Biology Methods; Elsevier Inc.: Amsterdam, The Netherlands, 2022; pp. 505–533. ISBN 9780323902649. [Google Scholar]
- Manning, M.C.; Illangasekare, M.; Woody, R.W. Circular Dichroism Studies of Distorted α-Helices, Twisted β-Sheets, and β-Turns. Biophys. Chem. 1988, 31, 77–86. [Google Scholar] [CrossRef]
- Iyer, A.; Roeters, S.J.; Kogan, V.; Woutersen, S.; Claessens, M.M.A.E.; Subramaniam, V. C-Terminal Truncated α-Synuclein Fibrils Contain Strongly Twisted β-Sheets. J. Am. Chem. Soc. 2017, 139, 15392–15400. [Google Scholar] [CrossRef]
- Ghosh, D.; Singh, P.K.; Sahay, S.; Jha, N.N.; Jacob, R.S.; Sen, S.; Kumar, A.; Riek, R.; Maji, S.K. Structure Based Aggregation Studies Reveal the Presence of Helix-Rich Intermediate during α-Synuclein Aggregation. Sci. Rep. 2015, 5, 9228. [Google Scholar] [CrossRef] [Green Version]
- Shoffner, S.K.; Schnell, S. Estimation of the Lag Time in a Subsequent Monomer Addition Model for Fibril Elongation. Phys. Chem. Chem. Phys. 2016, 18, 21259–215698. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.F. Fluorescence Quenching As A Parameter For Measuring Complex Formation Between Metal Ions And Aromatic Amino Acids And Peptides. Anal. Lett. 1986, 19, 963–977. [Google Scholar] [CrossRef]
- Wang, G.; Wang, A.J.; Hu, K.S. Tryptophan Fluorescence Quenching by Alkaline Earth Metal Cations in Deionized Bacteriorhodopsin. J. Photochem. Photobiol. B Biol. 2000, 59, 38–41. [Google Scholar] [CrossRef]
- Gorantla, N.V.; Landge, V.G.; Nagaraju, P.G.; Priyadarshini Cg, P.; Balaraman, E.; Chinnathambi, S. Molecular Cobalt(II) Complexes for Tau Polymerization in Alzheimer’s Disease. ACS Omega 2019, 4, 16702–16714. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, D.; Kaji, Y.; Yagi, H.; Sakurai, K.; Kawakami, T.; Naiki, H.; Goto, Y. Destruction of Amyloid Fibrils of Keratoepithelin Peptides by Laser Irradiation Coupled with Amyloid-Specific Thioflavin T. J. Biol. Chem. 2011, 286, 10856–10863. [Google Scholar] [CrossRef] [Green Version]
- Barber-Zucker, S.; Shaanan, B.; Zarivach, R. Transition Metal Binding Selectivity in Proteins and Its Correlation with the Phylogenomic Classification of the Cation Diffusion Facilitator Protein Family. Sci. Rep. 2017, 7, 16381. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Hu, X.; Zhang, X.; Gao, S.; Ding, C.; Feng, Y.; Bao, W. Identification of Metal Ion Binding Sites Based on Amino Acid Sequences. PLoS ONE 2017, 12, e0183756. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, B.J.H.; Gruppen, H. Prediction of Molar Extinction Coefficients of Proteins and Peptides Using UV Absorption of the Constituent Amino Acids at 214 Nm to Enable Quantitative Reverse Phase High-Performance Liquid Chromatography-Mass Spectrometry Analysis. J. Agric. Food Chem. 2007, 55, 5445–5451. [Google Scholar] [CrossRef]
- Williams, R.M.; Obradovi, Z.; Mathura, V.; Braun, W.; Garner, E.C.; Takayama, S.; Brown, C.J.; Dunker, A.K. The Protein Non-Folding Problem: Amino Acid Determinants of Intrinsic Order and Disorder. Pac Symp Biocomp. 2001, 2000, 89–100. [Google Scholar]
- Peng, K.; Vucetic, S.; Radivojac, P.; Brown, C.J.; Dunker, A.K.; Obradovic, Z. Optimizing Long Intrinsic Disorder Predictors With Protein Evolutionary Information. J. Bioinform. Comput. Biol. 2005, 3, 35–60. [Google Scholar] [CrossRef]
- Garner; Romero; Dunker; Brown; Obradovic Predicting Binding Regions within Disordered Proteins. Genome Inform. Ser. Workshop Genome Inform. 1999, 10, 41–50.
- Walsh, I.; Martin, A.J.M.; Di domenico, T.; Tosatto, S.C.E. Espritz: Accurate and Fast Prediction of Protein Disorder. Bioinformatics 2012, 28, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Ishida, T.; Kinoshita, K. PrDOS: Prediction of Disordered Protein Regions from Amino Acid Sequence. Nucleic Acids Res. 2007, 35, 460–464. [Google Scholar] [CrossRef] [Green Version]
- Erdos, G.; Pajkos, M.; Dosztányi, Z. IUPred3: Prediction of Protein Disorder Enhanced with Unambiguous Experimental Annotation and Visualization of Evolutionary Conservation. Nucleic Acids Res. 2021, 49, 297–303. [Google Scholar] [CrossRef]
- Emenecker, R.J.; Griffith, D.; Holehouse, A.S. Metapredict: A Fast, Accurate, and Easy-to-Use Predictor of Consensus Disorder and Structure. Biophys. J. 2021, 120, 4312–4319. [Google Scholar] [CrossRef]
- Bernhofer, M.; Dallago, C.; Karl, T.; Satagopam, V.; Heinzinger, M.; Littmann, M.; Olenyi, T.; Qiu, J.; Schütze, K.; Yachdav, G.; et al. PredictProtein—Predicting Protein Structure and Function for 29 Years. Nucleic Acids Res. 2021, 49, 535–540. [Google Scholar] [CrossRef]
- Dass, R.; Mulder, F.A.A.; Nielsen, J.T. ODiNPred: Comprehensive Prediction of Protein Order and Disorder. Sci. Rep. 2020, 10, 14780. [Google Scholar] [CrossRef]
- Xue, B.; Dunbrack, R.L.; Williams, R.W.; Dunker, A.K.; Uversky, V.N. PONDR-FIT: A Meta-Predictor of Intrinsically Disordered Amino Acids. Biochim. Biophys. Acta—Proteins Proteom. 2010, 1804, 996–1010. [Google Scholar] [CrossRef] [Green Version]
- Micsonai, A.; Moussong, É.; Wien, F.; Boros, E.; Vadászi, H.; Murvai, N.; Lee, Y.; Molnár, T.; Réfrégiers, M.; Goto, Y.; et al. BeStSel: Webserver for Secondary Structure and Fold Prediction for Protein CD Spectroscopy. Nucleic Acids Res. 2022, 50, 90–98. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Structure Content (%) | ||||||
|---|---|---|---|---|---|---|
| C-TER (time) | RMSD | α-Helix | Antiparallel β-Sheet | Parallel β-Sheet | Turn | Coil |
| C-TERb (0 h) | 0.04 | 4 | 38 | 0 | 17 | 40 |
| C-TERb (24 h) | 0.05 | 5 | 35 | 0 | 15 | 46 |
| C-TERb (48 h) | 0.05 | 0 | 38 | 3 | 16 | 43 |
| C-TERb (72 h) | 0.08 | 0 | 24 | 9 | 21 | 46 |
| C-TERb (96 h) | 0.08 | 0 | 24 | 15 | 21 | 40 |
| C-TERa (0 h) | 0.03 | 2 | 32 | 0 | 18 | 48 |
| C-TERa (24 h) | 0.02 | 0 | 37 | 0 | 18 | 45 |
| C-TERa (48 h) | 0.04 | 0 | 37 | 0 | 16 | 47 |
| C-TERa (72 h) | 0.06 | 0 | 34 | 5 | 19 | 42 |
| C-TERa (96 h) | 0.07 | 0 | 25 | 10 | 22 | 43 |
| Secondary Structure Content (%) | ||||||
|---|---|---|---|---|---|---|
| C-TER (M2+) | RMSD | α-Helix | Antiparallel β-Sheet | Parallel β-Sheet | Turn | Coil |
| C-TERb | 0.08 | 0 | 24 | 9 | 21 | 46 |
| C-TERb (Cu2+) | 0.06 | 0 | 31 | 8 | 18 | 43 |
| C-TERb (Mg2+) | 0.06 | 0 | 27 | 8 | 20 | 45 |
| C-TERb (Ni2+) | 0.07 | 0 | 33 | 6 | 20 | 41 |
| C-TERb (Zn2+) | 0.07 | 0 | 30 | 7 | 19 | 44 |
| C-TERa | 0.06 | 0 | 34 | 5 | 19 | 42 |
| C-TERa (Cu2+) | 0.05 | 0 | 33 | 5 | 17 | 45 |
| C-TERa (Mg2+) | 0.09 | 0 | 21 | 11 | 23 | 45 |
| C-TERa (Ni2+) | 0.05 | 0 | 38 | 3 | 18 | 41 |
| C-TERa (Zn2+) | 0.06 | 0 | 38 | 3 | 18 | 41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leyder, T.; Mignon, J.; Mottet, D.; Michaux, C. Unveiling the Metal-Dependent Aggregation Properties of the C-terminal Region of Amyloidogenic Intrinsically Disordered Protein Isoforms DPF3b and DPF3a. Int. J. Mol. Sci. 2022, 23, 15291. https://doi.org/10.3390/ijms232315291
Leyder T, Mignon J, Mottet D, Michaux C. Unveiling the Metal-Dependent Aggregation Properties of the C-terminal Region of Amyloidogenic Intrinsically Disordered Protein Isoforms DPF3b and DPF3a. International Journal of Molecular Sciences. 2022; 23(23):15291. https://doi.org/10.3390/ijms232315291
Chicago/Turabian StyleLeyder, Tanguy, Julien Mignon, Denis Mottet, and Catherine Michaux. 2022. "Unveiling the Metal-Dependent Aggregation Properties of the C-terminal Region of Amyloidogenic Intrinsically Disordered Protein Isoforms DPF3b and DPF3a" International Journal of Molecular Sciences 23, no. 23: 15291. https://doi.org/10.3390/ijms232315291