
Key Amino Acids for Transferase Activity of GDSL Lipases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Sequence Alignment and Two-Entropy Analysis of GELPs
2.2. Molecular Phylogenetic Analysis of GELPs
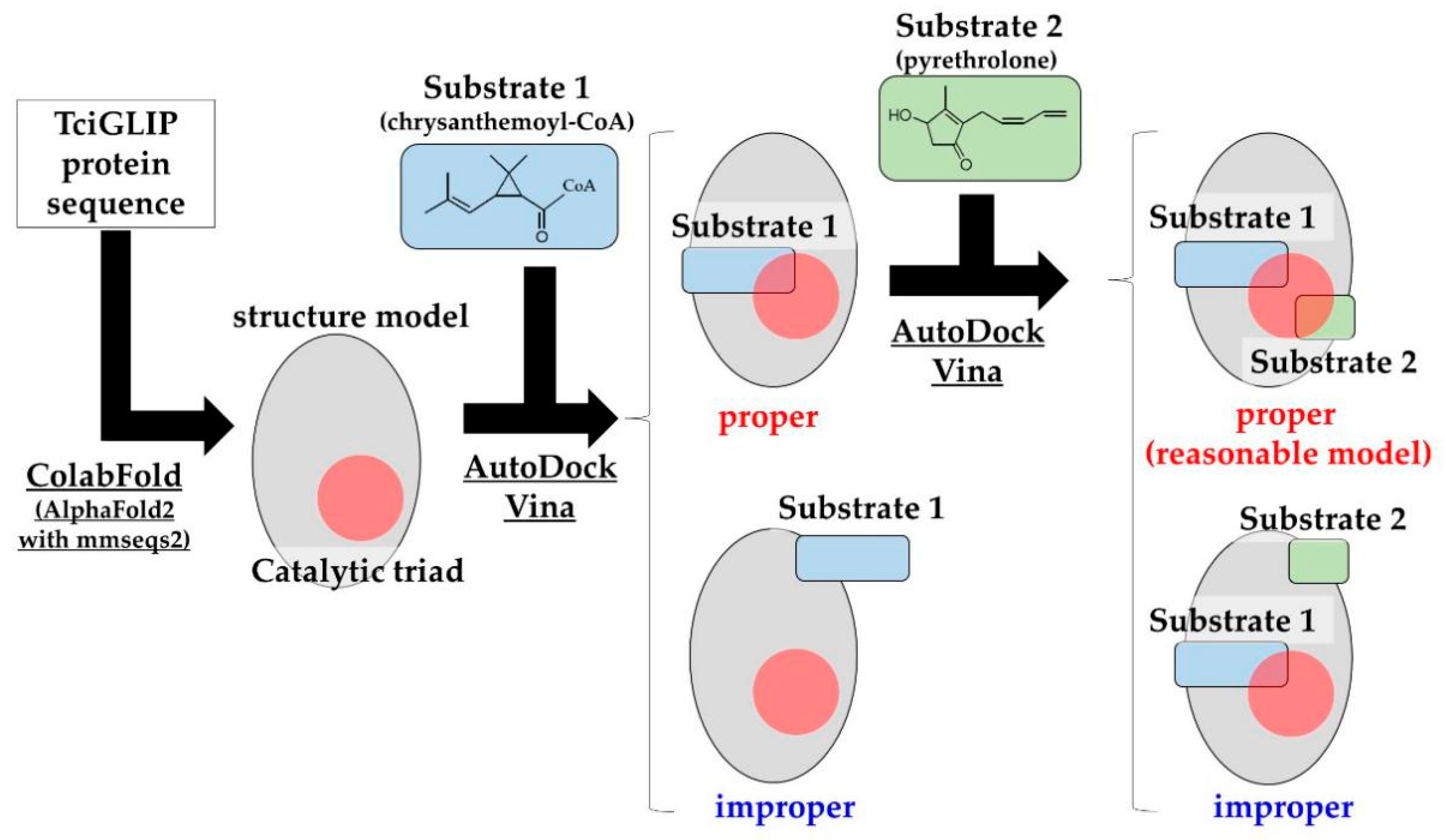
2.3. Prediction of Protein Structures and Substrate Docking Simulations of TciGLIP
3. Materials and Methods
3.1. Sequence Collection and Phylogenetic Analysis
3.2. Two-Entropy Analysis of GELPs
3.3. Protein Structure Modeling and Substrate-Binding Simulations of TciGLIP
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shen, G.; Sun, W.; Chen, Z.; Shi, L.; Hong, J.; Shi, J. Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development. Plants 2022, 11, 468. [Google Scholar] [CrossRef]
- Clauss, K.; Baumert, A.; Nimtz, M.; Milkowski, C.; Strack, D. Role of a GDSL lipase-like protein as sinapine esterase in Brassicaceae. Plant J. 2008, 53, 802–813. [Google Scholar] [CrossRef]
- Takahashi, K.; Shimada, T.; Kondo, M.; Tamai, A.; Mori, M.; Nishimura, M.; Hara-Nishimura, I. Ectopic expression of an esterase, which is a candidate for the unidentified plant cutinase, causes cuticular defects in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 123–131. [Google Scholar] [CrossRef]
- Gao, M.; Yin, X.; Yang, W.; Lam, S.M.; Tong, X.; Liu, J.; Wang, X.; Li, Q.; Shui, G.; He, Z. GDSL lipases modulate immunity through lipid homeostasis in rice. PLoS Pathog. 2017, 13, e1006724. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, M.; Woll, J.; Giritch, A.; Genady, E.; Ma, X.; Stockigt, J. Functional expression of an ajmaline pathway-specific esterase from Rauvolfia in a novel plant-virus expression system. Planta 2005, 222, 888–898. [Google Scholar] [CrossRef]
- Kikuta, Y.; Ueda, H.; Takahashi, M.; Mitsumori, T.; Yamada, G.; Sakamori, K.; Takeda, K.; Furutani, S.; Nakayama, K.; Katsuda, Y.; et al. Identification and characterization of a GDSL lipase-like protein that catalyzes the ester-forming reaction for pyrethrin biosynthesis in Tanacetum cinerariifolium—A new target for plant protection. Plant J. 2012, 71, 183–193. [Google Scholar] [CrossRef]
- Yamashiro, T.; Shiraishi, A.; Nakayama, K.; Satake, H. Draft Genome of Tanacetum Coccineum: Genomic Comparison of Closely Related Tanacetum-Family Plants. Int. J. Mol. Sci. 2022, 23, 7039. [Google Scholar] [CrossRef]
- Zeng, T.; Li, J.W.; Zhou, L.; Xu, Z.Z.; Li, J.J.; Hu, H.; Luo, J.; Zheng, R.R.; Wang, Y.Y.; Wang, C.Y. Transcriptional Responses and GCMS Analysis for the Biosynthesis of Pyrethrins and Volatile Terpenes in Tanacetum coccineum. Int. J. Mol. Sci. 2021, 22, 13005. [Google Scholar] [CrossRef]
- Watkins, J.L.; Li, M.; McQuinn, R.P.; Chan, K.X.; McFarlane, H.E.; Ermakova, M.; Furbank, R.T.; Mares, D.; Dong, C.; Chalmers, K.J.; et al. A GDSL Esterase/Lipase Catalyzes the Esterification of Lutein in Bread Wheat. Plant Cell 2019, 31, 3092–3112. [Google Scholar] [CrossRef]
- Teutschbein, J.; Gross, W.; Nimtz, M.; Milkowski, C.; Hause, B.; Strack, D. Identification and localization of a lipase-like acyltransferase in phenylpropanoid metabolism of tomato (Solanum lycopersicum). J. Biol. Chem. 2010, 285, 38374–38381. [Google Scholar] [CrossRef]
- Akoh, C.C.; Lee, G.C.; Liaw, Y.C.; Huang, T.H.; Shaw, J.F. GDSL family of serine esterases/lipases. Prog. Lipid Res. 2004, 43, 534–552. [Google Scholar] [CrossRef]
- Kikuta, Y.; Yamada, G.; Mitsumori, T.; Takeuchi, T.; Nakayama, K.; Katsuda, Y.; Hatanaka, A.; Matsuda, K. Requirement of catalytic-triad and related amino acids for the acyltransferase activity of Tanacetum cinerariifolium GDSL lipase/esterase TcGLIP for ester-bond formation in pyrethrin biosynthesis. Biosci. Biotechnol. Biochem. 2013, 77, 1822–1825. [Google Scholar] [CrossRef] [Green Version]
- Abdelkafi, S.; Ogata, H.; Barouh, N.; Fouquet, B.; Lebrun, R.; Pina, M.; Scheirlinckx, F.; Villeneuve, P.; Carriere, F. Identification and biochemical characterization of a GDSL-motif carboxylester hydrolase from Carica papaya latex. Biochim. Biophys. Acta 2009, 1791, 1048–1056. [Google Scholar] [CrossRef]
- Witasari, L.D.; Huang, F.-C.; Schwab, W. Identification and characterization of strawberry FvGELP1 (Fragaria vesca GDSL esterase/lipase). Biodiversitas 2022, 23, 907–915. [Google Scholar] [CrossRef]
- Li, K.B. ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinformatics 2003, 19, 1585–1586. [Google Scholar] [CrossRef] [Green Version]
- Ye, K.; Lameijer, E.W.; Beukers, M.W.; Ijzerman, A.P. A two-entropies analysis to identify functional positions in the transmembrane region of class A G protein-coupled receptors. Proteins 2006, 63, 1018–1030. [Google Scholar] [CrossRef] [Green Version]
- Zea, D.J.; Anfossi, D.; Nielsen, M.; Marino-Buslje, C. MIToS.jl: Mutual information tools for protein sequence analysis in the Julia language. Bioinformatics 2017, 33, 564–565. [Google Scholar] [CrossRef] [Green Version]
- van Westen, G.J.; van den Hoven, O.O.; van der Pijl, R.; Mulder-Krieger, T.; de Vries, H.; Wegner, J.K.; Ijzerman, A.P.; van Vlijmen, H.W.; Bender, A. Identifying novel adenosine receptor ligands by simultaneous proteochemometric modeling of rat and human bioactivity data. J. Med. Chem. 2012, 55, 7010–7020. [Google Scholar] [CrossRef]
- Gonzalez, A.; Duran, L.S.; Araya-Secchi, R.; Garate, J.A.; Pessoa-Mahana, C.D.; Lagos, C.F.; Perez-Acle, T. Computational modeling study of functional microdomains in cannabinoid receptor type 1. Bioorg. Med. Chem. 2008, 16, 4378–4389. [Google Scholar] [CrossRef]
- Szollosi, D.; Erdei, A.; Gyimesi, G.; Magyar, C.; Hegedus, T. Access Path to the Ligand Binding Pocket May Play a Role in Xenobiotics Selection by AhR. PLoS ONE 2016, 11, e0146066. [Google Scholar] [CrossRef]
- Rabal, O.; Pastor, F.; Villanueva, H.; Soldevilla, M.M.; Hervas-Stubbs, S.; Oyarzabal, J. In Silico Aptamer Docking Studies: From a Retrospective Validation to a Prospective Case Study-TIM3 Aptamers Binding. Mol. Ther. Nucleic Acids 2016, 5, e376. [Google Scholar] [CrossRef]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamashiro, T.; Shiraishi, A.; Nakayama, K.; Satake, H. Key Amino Acids for Transferase Activity of GDSL Lipases. Int. J. Mol. Sci. 2022, 23, 15141. https://doi.org/10.3390/ijms232315141
Yamashiro T, Shiraishi A, Nakayama K, Satake H. Key Amino Acids for Transferase Activity of GDSL Lipases. International Journal of Molecular Sciences. 2022; 23(23):15141. https://doi.org/10.3390/ijms232315141
Chicago/Turabian StyleYamashiro, Takanori, Akira Shiraishi, Koji Nakayama, and Honoo Satake. 2022. "Key Amino Acids for Transferase Activity of GDSL Lipases" International Journal of Molecular Sciences 23, no. 23: 15141. https://doi.org/10.3390/ijms232315141